Burkholderia sp. JXJ16低溫耐有機溶劑脂肪酶產酶條件優化及粗酶酶學性質
2017-03-14 08:46:30謝玉婷查代明石紅璆張炳火李漢全
食品工業科技 2017年4期
關鍵詞:影響
謝玉婷,查代明,石紅璆,張炳火,李漢全
(九江學院藥學與生命科學學院,江西九江 332000)

謝玉婷,查代明*,石紅璆,張炳火,李漢全
(九江學院藥學與生命科學學院,江西九江 332000)
以Burkholderiasp. JXJ-16為出發菌株,對其低溫耐有機溶劑脂肪酶的產酶條件進行單因素實驗并研究其粗酶酶學性質。該菌株的最佳產酶條件為:蔗糖3.75 g/L、尿素11.25 g/L、K2HPO42 g/L、(NH4)2SO41 g/L、MnSO40.25 g/L、豬油乳化液體積分數2.5%,初始pH9.0、培養溫度30 ℃、裝樣量20 mL/250 mL、接種量3%、發酵時間20 h。JXJ-16脂肪酶粗酶對中鏈對硝基苯酚酯有最大水解活力,最適底物為對硝基苯酚辛酸酯;該粗酶在35 ℃、pH8.5~9.0時酶活力最高,且具有較好的溫度(30~60 ℃)和pH(5.0~10.5)穩定性;Na+、Mg2+、Ca2+、Mn2+、EDTA對粗酶活力具有激活作用,Zn2+、Cu2+、Fe3+對粗酶活力具有抑制作用,K+對粗酶活力沒有顯著性影響;除乙醇和乙腈外,該粗酶在一定體積分數的異丙醇、甲醇、丙酮、乙酸乙酯、三氯甲烷、二甲苯和正己烷中具有良好耐受性,且處于激活狀態。綜上,該菌株能利用廉價易得的培養基原料達到最佳產酶效果,其所產脂肪酶為低溫堿性脂肪酶,具有較好的溫度和pH穩定性,對多數供試有機溶劑具有良好耐受性。
低溫脂肪酶,伯克霍爾德屬,有機溶劑耐受性
脂肪酶(triacylglycerol acylhydrolases,EC 3.1.1.3)屬于α/β型水解酶超家族,在油水界面上能催化長鏈脂肪酸甘油酯水解為甘油和脂肪酸,而在有機相中的催化反應能極大地拓寬其應用,如酯化、轉酯、醇解、氨解、酸解反應等,因而脂肪酶具有良好的有機溶劑耐受性對其應用非常有利[1-3]。與水相相比,脂肪酶在有機相中的催化反應一般來說表現出多種優勢:更高的活性、穩定性、區域選擇性和/或立體選擇性;更好的底物溶解性和相對容易的產物回收;降低底物和/或產物的抑制程度;使反應平衡向合成方向移動[4]。因此,脂肪酶的有機相催化反應已被廣泛應用于多個工業領域,比如食品、油脂、新型材料、精細化工、醫藥、生物能源等[5-6]。
大多數脂肪酶在有機溶劑中的穩定性很差,不能有效地發揮其催化功能[7],因此從自然界中篩選有機溶劑耐受性脂肪酶是當前和今后的熱點方向之一。Fan等[8]利用宏基因組技術從油田土壤中克隆到一個脂肪酶基因,其表達產物具有較好的有機溶劑耐受性。李俊峰等[9]以苯作為選擇壓力,從花生地土壤中篩選出一株耐有機溶劑脂肪酶產生菌。Xie等[10]以二甲基亞砜和甲苯為選擇壓力篩選出一株有機溶劑耐受性菌株BurkholderiacepaciaRQ3,其分泌的脂肪酶具有廣譜有機溶劑耐受性。伯克霍爾德屬(Burkholderia)脂肪酶具有活性高、抗逆性好等優良特性,已被廣泛應用于工業生產,比如藥品生產、洗滌劑配方、有機合成、生物能源生產等[11-12]。本實驗以Burkholderiasp. JXJ-16為出發菌株,對其脂肪酶的產酶條件以及粗酶酶學性質進行研究,旨在為進一步應用提供理論基礎和技術指導。
1 材料與方法
1.1 材料與儀器
Burkholderiasp. JXJ-16 本實驗室分離并保藏;對硝基苯酚酯(p-nitrophenyl esters,pNP酯)、Tris堿 美國Sigma公司;聚乙烯醇(PVA)、橄欖油、牛肉浸膏 國藥集團化學試劑有限公司;其他植物油 中糧集團有限公司;蛋白胨、酵母提取物 安琪酵母股份有限公司;酵母浸膏 北京雙旋微生物培養基制品廠;其他化學試劑 均為國產分析純;LB平板培養基(g/L) 蛋白胨10、酵母提取物5、NaCl 10、瓊脂條20,自然pH;種子培養基(g/L) 蛋白胨10、酵母提取物5、NaCl 10,自然pH;基礎發酵培養基(g/L) 麥芽糖5、蛋白胨10、K2HPO42、(NH4)2SO41、MgSO40.5、橄欖油乳化液體積分數2%,pH7.0;橄欖油乳化液 橄欖油與體積分數2%的PVA按1∶3的比例混合,10000 r/min乳化10 min即可。
Sartorius電子天平、PB-10型pH計 賽多利斯科學儀器(北京)有限公司;XHF-D型高速分散器 寧波新芝生物科技有限公司;DHP-9162型電熱恒溫培養箱 上海一恒科學儀器有限公司;SKY-2112C型恒溫培養搖床 上海蘇坤實業有限公司;CT15RT型高速冷凍離心機 上海天美生化儀器設備工程有限公司;HH-4型數顯恒溫水浴鍋 常州榮華儀器制造有限公司;UV-3200PC型紫外可見分光光度計 上海美譜達儀器有限公司。
1.2 實驗方法
1.2.1 脂肪酶活力的測定 挑取LB平板培養基上的Burkholderiasp. JXJ-16單菌落,接種于種子培養基中,28 ℃、180 r/min培養20 h獲得種子液,再以2%的接種量接種于發酵培養基中,28 ℃、180 r/min培養24 h;4 ℃、5000 r/min離心10 min,取上清液測定脂肪酶活力,其測定方法為基于pNP酯底物的分光光度法[4]。具體操作為:配制反應體系,包括1.45 mL Tris-HCl(50 mmol/L,pH8.0)和10 μLpNP酯(10 mmol/L);30 ℃預熱5 min后,實驗組加入40 μL脂肪酶樣品,對照組加入等量的失活脂肪酶樣品;30 ℃反應5 min后,冰上放置5 min終止反應;取適量反應液,410 nm下測量吸光值。
1.2.2 培養基的確定
1.2.2.1 碳源對產酶的影響 以基礎發酵培養基為基礎,分別加入等量的葡萄糖、蔗糖、乳糖、可溶性淀粉以代替麥芽糖進行發酵培養,測定酶活力以確定最佳碳源,不添加碳源設為對照組;添加不同量的最佳碳源(1.250、1.875、2.500、3.125、3.750、4.375 g/L),以確定最佳碳源添加量。
1.2.2.2 氮源對產酶的影響 以基礎發酵培養基為基礎,分別加入等量的NaNO3、牛肉浸膏、酵母浸膏、尿素以代替蛋白胨進行發酵培養,測定酶活力以確定最佳氮源,不添加氮源設為對照組;添加不同量的最佳氮源(7.50、8.75、10.00、11.25、12.50 g/L),以確定最佳氮源添加量。
1.2.2.3 誘導劑對菌株產酶的影響 以基礎發酵培養基為基礎,分別加入等量的菜籽油乳化液、豬油乳化液、玉米油乳化液、花生油乳化液以代替橄欖油乳化液作為誘導劑進行發酵培養,測定酶活力以確定最佳誘導劑,不添加誘導劑設為對照組;添加不同量的最佳誘導劑(2.00%、2.25%、2.50%、2.75%、3.00%),以確定最佳誘導劑添加量。
1.2.2.4 金屬離子對產酶的影響 以基礎發酵培養基為基礎,分別加入等量的CuSO4、CaSO4、FeSO4、MnSO4以代替MgSO4進行發酵培養,測定酶活力以確定金屬離子對產酶的影響,不添加金屬離子設為對照組;添加不同量的最佳金屬離子(0.1250、0.1875、0.2500、0.3125、0.3750 g/L),以確定最佳金屬離子添加量。
1.2.3 發酵條件的確定 采用優化后的發酵培養基,以初始發酵條件(初始pH7.0、培養溫度28 ℃、裝樣量20 mL/250 mL、接種量2%、發酵時間24 h)為基礎,按照以下操作分別優化初始pH、培養溫度、裝樣量、接種量和發酵時間,以確定發酵條件。
1.2.3.1 初始pH對產酶的影響 將發酵培養基的初始pH設置為4、5、6、7、8、9、10七個梯度,發酵培養后測定酶活力,以確定最佳產酶pH。
1.2.3.2 培養溫度對產酶的影響 種子液接種于發酵培養基中后,分別選定15、20、25、30、35 ℃五個溫度梯度進行發酵培養,測定酶活力以確定最佳產酶溫度。
1.2.3.3 裝樣量對產酶的影響 250 mL三角瓶中分別裝有10、20、30、40 mL培養基進行發酵培養,測定酶活力以確定產酶的最佳裝樣量。
1.2.3.4 接種量對產酶的影響 種子液分別以0.5%、1%、2%、3%、4%、5%的接種量接種于發酵培養基中,發酵培養后測定酶活力,以確定產酶的最佳接種量。
1.2.3.5 發酵時間曲線的確定 種子液分別接種于7瓶發酵培養基中同時進行培養,每隔4 h取發酵液進行酶活力測定,以確定發酵時間曲線。
1.2.4 粗酶酶學性質研究
1.2.4.1 底物特異性 以對硝基苯酚乙酸酯(C2)、對硝基苯酚丁酸酯(C4)、對硝基苯酚辛酸酯(C8)、對硝基苯酚葵酸酯(C10)、對硝基苯酚月桂酸酯(C12)、對硝基苯酚豆蔻酸酯(C14)和對硝基苯酚棕櫚酸酯(C16)為底物分別測定酶活力,最高酶活力設為100%。
1.2.4.2 溫度對酶活力的影響 在50 mmol/L Tris-HCl緩沖液(pH8.0)中測定不同反應溫度(15~45 ℃)下的酶活力,最高酶活力設為100%。粗酶樣品在不同溫度(30~60 ℃)下溫浴1 h,測定殘余酶活力以確定溫度穩定性,處理前的酶活力設為100%。
1.2.4.3 pH對酶活力的影響 在50 mmol/L Na2HPO4-檸檬酸緩沖液(pH5.0~8.0)和50 mmol/L Tris-HCl緩沖液(pH8.0~10.5)中測定不同反應pH下的酶活力,最高酶活力設為100%。在不同pH緩沖液中,28 ℃水浴處理1 h,測定殘余酶活力以確定pH穩定性,pH7.0時的殘余酶活力設為100%。
1.2.4.4 金屬離子和EDTA對酶活力的影響 粗酶樣品中分別加入終濃度為10 mmol/L的金屬離子(Na+、K+、Ca2+、Mg2+、Mn2+、Cu2+、Zn2+和Fe3+)和EDTA,加入等量雙蒸水的粗酶樣品設為對照,28 ℃搖床處理1 h,測定殘余酶活力,對照組的酶活力設為100%。
1.2.4.5 有機溶劑對酶活力的影響 粗酶樣品中分別加入有機溶劑(甲醇、乙醇、異丙醇、乙酸乙酯、二甲苯、乙腈、丙酮、三氯甲烷和正己烷),有機溶劑的體積分數設置三個水平,分別為10%、30%、50%,加入等量雙蒸水的粗酶樣品設為對照,28 ℃搖床處理1 h,測定殘余酶活力,對照組的酶活力設為100%。
1.2.5 統計學分析 實驗數據來自三次獨立重復實驗,以平均數±標準差表示。
2 結果與分析
2.1 產酶條件的確定
2.1.1 培養基的確定
2.1.1.1 碳源對產酶的影響 由圖1A可知,供試碳源對菌株JXJ-16產酶的影響較小;蔗糖和麥芽糖對菌株產酶具有促進作用,但是它們的影響沒有顯著性;葡萄糖、乳糖和可溶性淀粉不同程度地抑制菌株產酶,其中葡萄糖對產酶沒有顯著性影響。因此,選擇相對廉價的蔗糖作為菌株產酶的最適碳源。由圖1B可知,蔗糖加入量為3.75 g/L時,菌株的產酶活力最強,因此選擇該濃度為蔗糖的最適加入量。

圖1 碳源(A)及蔗糖加入量(B)對菌株JXJ-16產脂肪酶的影響Fig.1 Effects of carbon sources(A)and sucrose concentration(B)on lipase production of strain JXJ-16
2.1.1.2 氮源對產酶的影響 由圖2A可知,供試氮源對菌株JXJ-16產酶的影響非常顯著,其中尿素對菌株產酶的促進作用最強,其次是酵母浸膏、蛋白胨和牛肉浸膏,而NaNO3對菌株產酶具有抑制作用,但是其影響沒有顯著性。由圖2B可知,尿素加入量為11.25 g/L時,菌株的產酶活力最強,因此選擇該濃度為尿素的最適加入量。
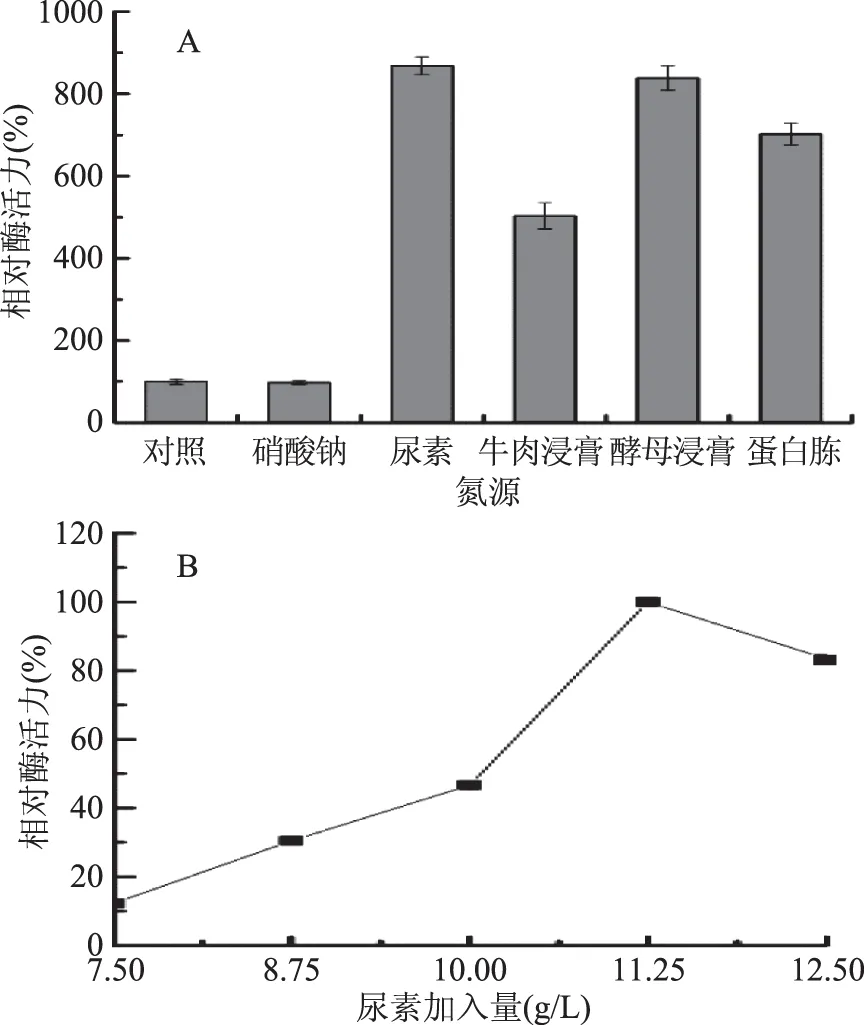
圖2 氮源(A)及尿素加入量(B)對菌株JXJ-16產脂肪酶的影響Fig.2 Effects of nitrogen sources(A)and urea concentration(B)on lipase production of strain JXJ-16
2.1.1.3 誘導物對產酶的影響 由圖3A可知,供試油脂乳化液作為誘導物對菌株JXJ-16產酶均具有顯著性促進作用,其中菜籽油的誘導效果最好,其次是橄欖油、花生油、豬油和玉米油,表明該菌株以誘導型方式表達脂肪酶。菜籽油、橄欖油、花生油和豬油之間的誘導效果沒有顯著性差異,因此選擇廉價易得的豬油作為菌株產酶的最適誘導物。由圖3B可知,豬油乳化液加入量為體積分數2.5%時,菌株的產酶活力最強,因此選擇該體積分數為豬油乳化液的最適加入量。
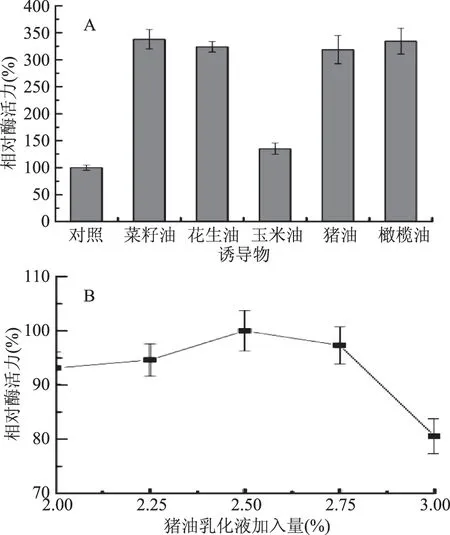
圖3 誘導物(A)及豬油乳化液加入量(B)對菌株JXJ-16產脂肪酶的影響Fig.3 Effects of inducers(A)and lard emulsion concentration(B)on lipase production of strain JXJ-16
2.1.1.4 金屬離子對產酶的影響 由圖4A可知,供試金屬離子對菌株JXJ-16產酶具有不同的影響,除Cu2+顯著性抑制菌株產酶外,其他4種金屬離子對菌株產酶均沒有顯著性影響,其中Mg2+、Fe2+和Mn2+有促進作用,而Ca2+有抑制作用。由圖4B可知,MnSO4加入量為0.25 g/L時,菌株的產酶活力最強,因此選擇該濃度為MnSO4的最適加入量。
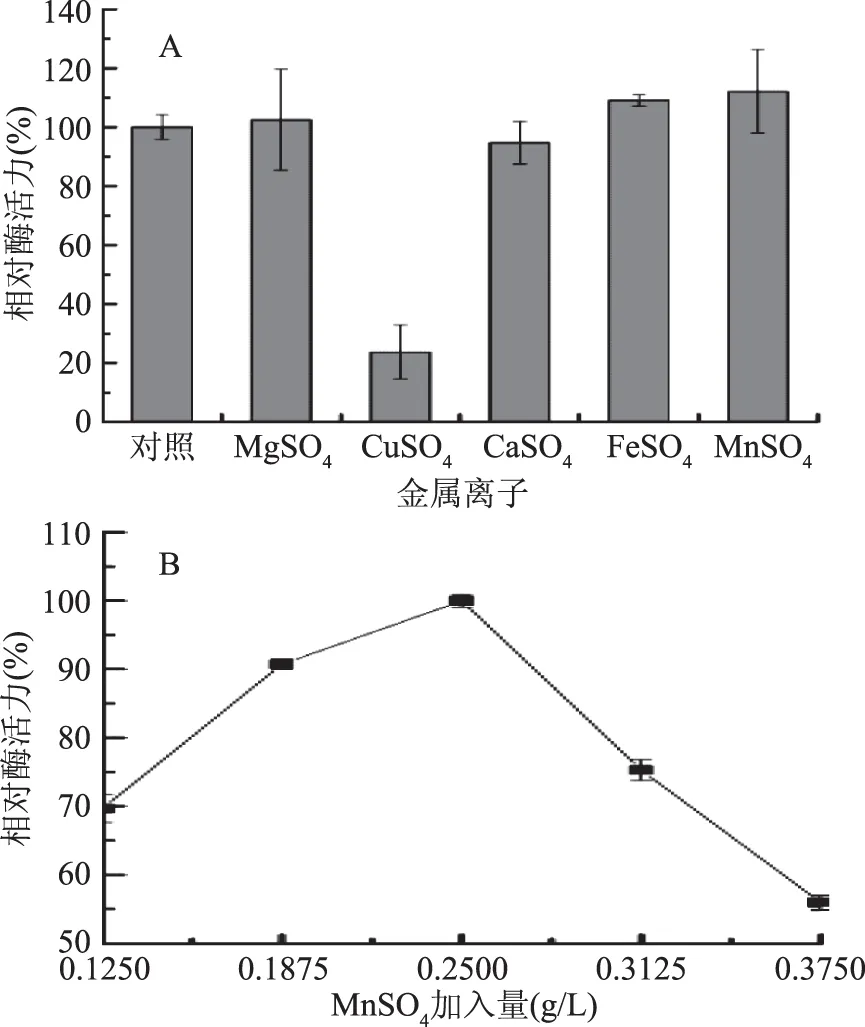
圖4 金屬離子(A)及MnSO4加入量(B)對菌株JXJ-16產脂肪酶的影響Fig.4 Effects of metal ions(A)and MnSO4 concentration(B) on lipase production of strain JXJ-16
2.1.2 發酵條件的確定 培養基初始pH、培養溫度、搖瓶裝樣量、菌種接種量和發酵時間對菌株JXJ-16產酶具有較大影響(圖5)。
由圖5A可知,菌株JXJ-16在初始pH為5.0~10.0的發酵培養基中具有較強的產酶活力,其中最佳產酶pH為9.0,而在初始pH為4.0的發酵培養基中幾乎沒有產酶活力。
由圖5B可知,培養溫度對菌株JXJ-16產酶的影響非常顯著,隨著溫度升高,菌株產酶活力逐漸增強,30 ℃時達到最佳,隨后迅速下降。
由圖5C可知,250 mL三角瓶裝樣量為10 mL時,菌株JXJ-16產酶活力最強,隨著裝樣量增加,菌株產酶活力逐漸下降。由于10 mL與20 mL時的酶活力沒有顯著性差異,因此選擇20 mL作為最佳裝樣量。
由圖5D可知,接種量是影響菌株JXJ-16產酶的關鍵因素之一,隨著接種量加大,菌株產酶活力逐漸增強,接種量為3%時達到最大,隨后略微下降。
由圖5E可知,菌株JXJ-16從培養8 h后開始逐漸增強產酶活力,培養16 h達到峰值,隨后略微下降。培養20 h的酶活力與培養16 h相比沒有顯著性差異,因此選擇培養20 h作為最佳發酵時間。
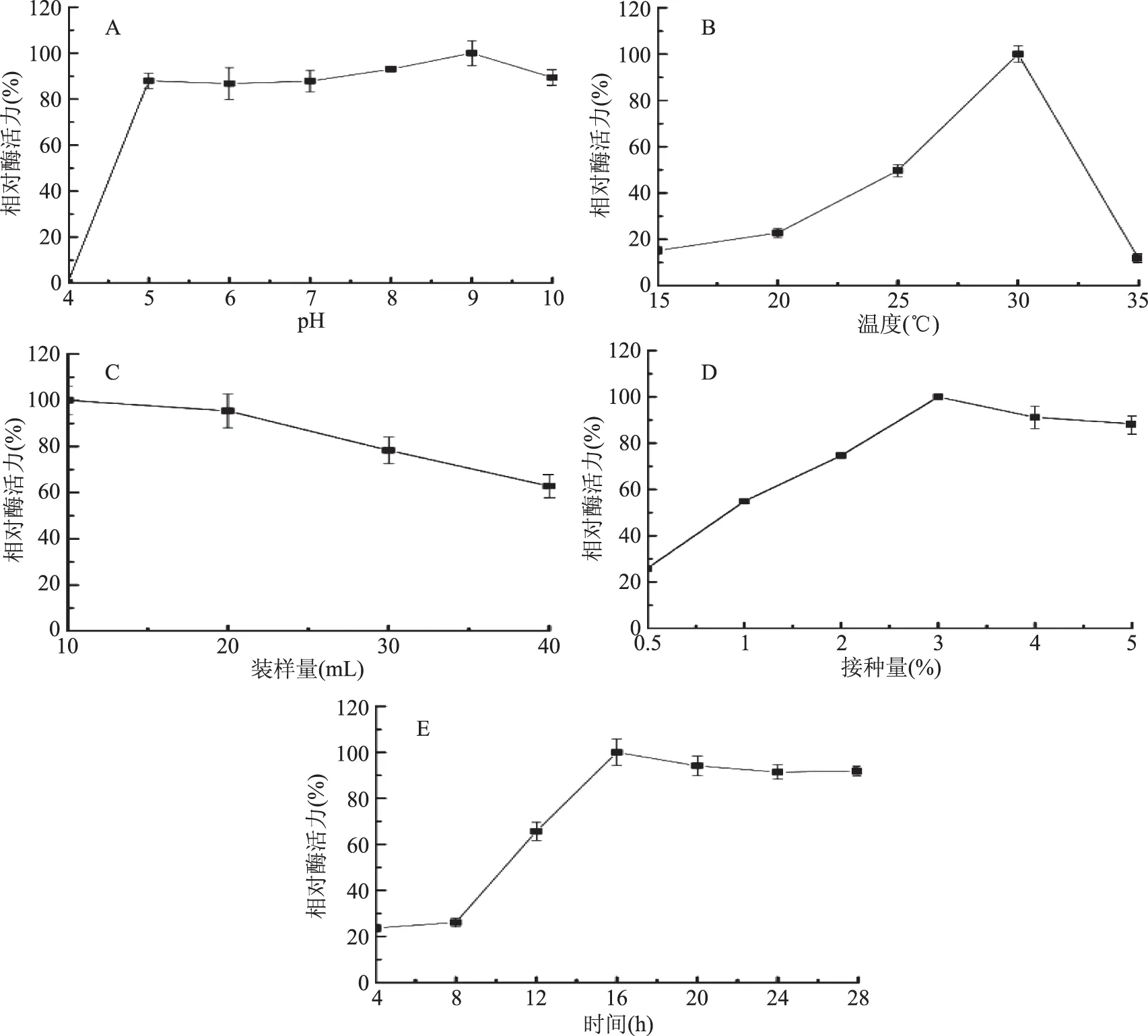
圖5 發酵條件對菌株JXJ-16產脂肪酶的影響Fig.5 Effects of fermentation conditions on lipase production of strain JXJ-16
2.2 粗酶酶學性質研究
2.2.1 底物特異性 由圖6可知,JXJ-16脂肪酶粗酶對中鏈pNP酯有最大水解活力,其最適底物為C8對硝基苯酚辛酸酯,其次為C10對硝基苯酚葵酸酯、C12對硝基苯酚月桂酸酯、C4對硝基苯酚丁酸酯、C14對硝基苯酚豆蔻酸酯、C2對硝基苯酚乙酸酯和C16對硝基苯酚棕櫚酸酯。
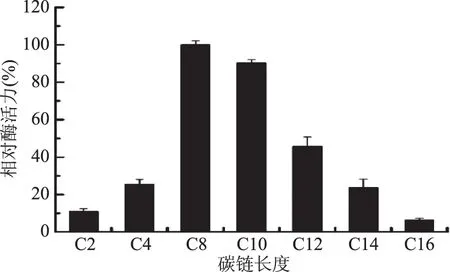
圖6 JXJ-16脂肪酶粗酶的底物特異性Fig.6 Substrate specificity of crude lipasefrom strain JXJ-16
2.2.2 溫度對酶活力的影響 由圖7A可知,JXJ-16脂肪酶粗酶的最適反應溫度為35 ℃,15~45 ℃時其酶活力保持在75%以上。由圖7B可知,30~60 ℃下溫浴1 h后,其殘余酶活力均保持在80%以上,其中30 ℃下處理1 h對酶活力沒有顯著性影響。低溫酶的最適反應溫度一般低于40 ℃,且通常具有一定的溫度穩定性[13]。因此,JXJ-16脂肪酶屬于低溫脂肪酶,且具有較好的溫度穩定性,這一特性非常有利于脂肪酶應用于生物轉化領域,特別是食品加工領域[14]。
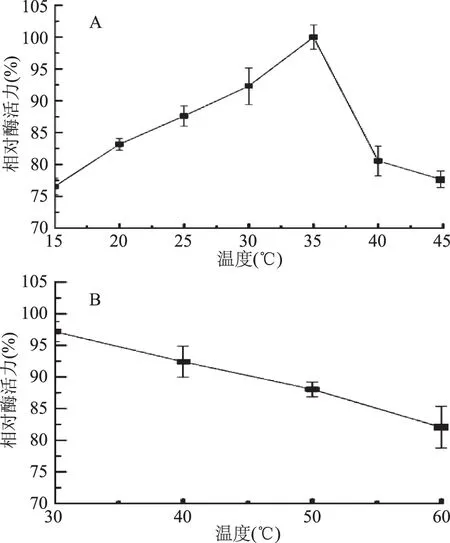
圖7 溫度對JXJ-16脂肪酶粗酶活力(A)和穩定性(B)的影響Fig.7 Effects of temperature on the activity(A) and the stability(B)of crude lipase from strain JXJ-16
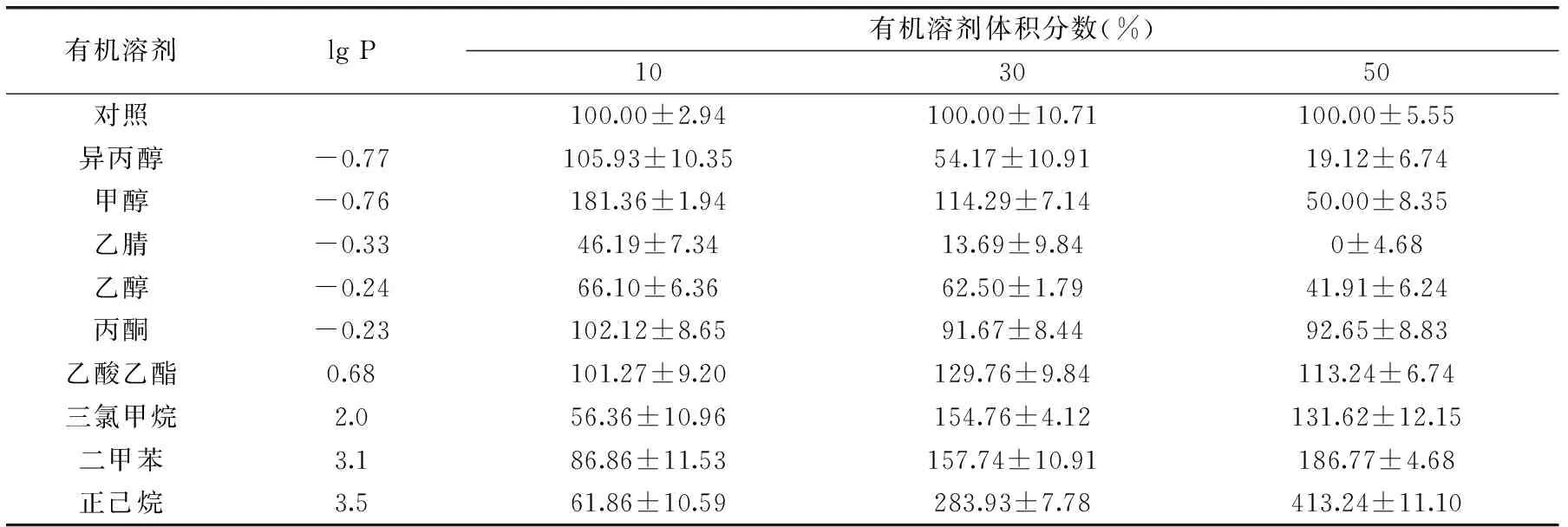
表1 有機溶劑對JXJ-16脂肪酶粗酶活力(%)的影響Table 1 Effects of organic solvents on the activity of crude lipase(%)from strain JXJ-16
2.2.3 pH對酶活力的影響 由圖8A可知,JXJ-16脂肪酶粗酶的最適反應pH為8.5~9.0,pH8.0~10.0時其酶活力保持在60%以上,pH低于8.0或高于10.0時其酶活力急劇下降。不同pH緩沖液(5.0~10.5)中30 ℃處理1 h后,殘余酶活力均保持在75%以上(圖8B)。可見,JXJ-16脂肪酶為低溫堿性脂肪酶,且具有較好的pH穩定性。
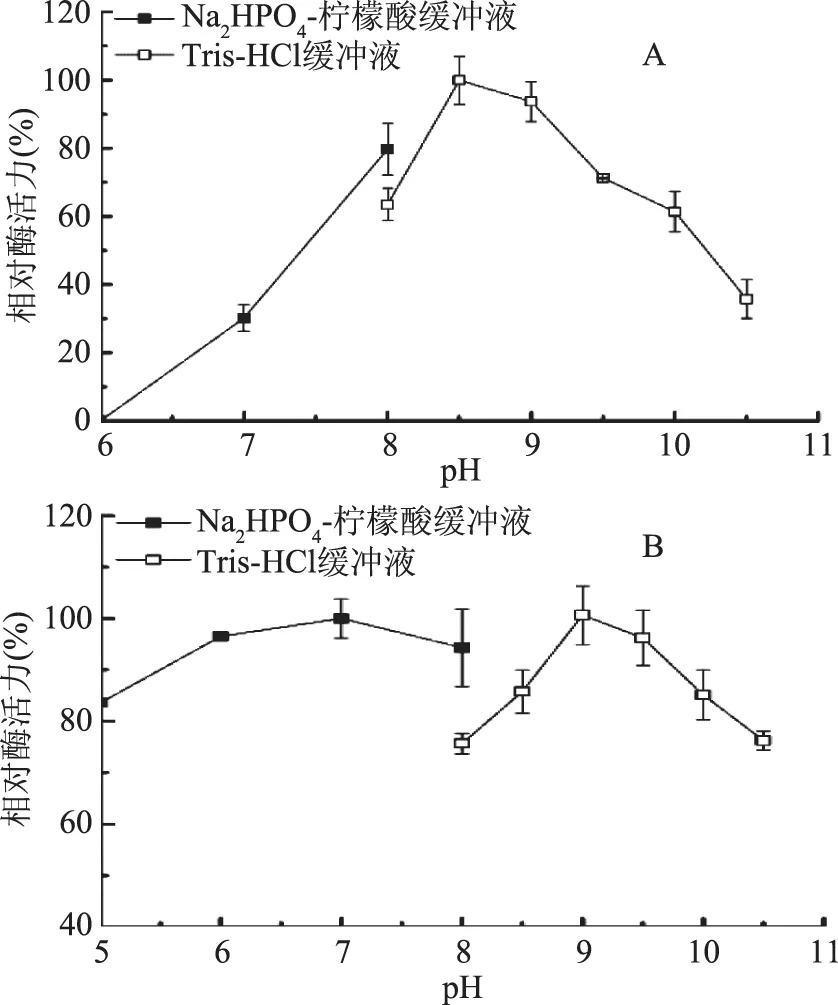
圖8 pH對JXJ-16脂肪酶粗酶活力(A)和穩定性(B)的影響Fig.8 Effects of pH on the activity(A) and the stability(B)of crude lipase from strain JXJ-16
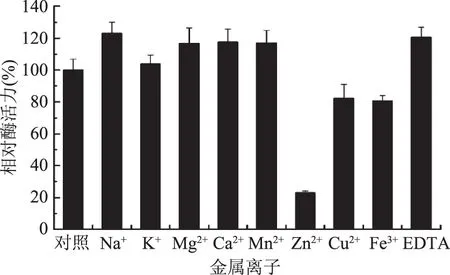
圖9 金屬離子和EDTA對JXJ-16脂肪酶粗酶活力的影響Fig.9 Effects of metal ions and EDTA on the activity of crude lipase from strain JXJ-16
2.2.4 金屬離子和EDTA對酶活力的影響 由圖9可知,供試金屬離子和EDTA對JXJ-16脂肪酶粗酶活力具有不同的影響,Na+、Mg2+、Ca2+、Mn2+、EDTA顯著性激活粗酶活力,Zn2+、Cu2+、Fe3+顯著性抑制粗酶活力,K+對粗酶活力沒有顯著性影響。可見,JXJ-16脂肪酶發揮催化作用不需要金屬離子的參與,因此其不屬于金屬酶。Mg2+、Ca2+、Mn2+的激活作用可能是由于它們與脂肪酸形成不溶性鹽而避免了產物的抑制作用[7]。
2.2.5 有機溶劑對酶活力的影響 由表1可知,有機溶劑對JXJ-16脂肪酶粗酶活力的影響與其lg P值有關。當lg P<0時,粗酶活力隨著有機溶劑體積分數的增大而降低,粗酶溶液在丙酮(10%~50%)、甲醇(10%~30%)和異丙醇(10%)中具有良好的耐受性,但是在乙醇(10%~50%)和乙腈(10%~50%)中其活力急劇下降甚至喪失。當0
3 結論
通過單因素分析,對影響菌株Burkholderiasp. JXJ-16產脂肪酶的發酵條件和培養基成分進行了單因素實驗,最佳產酶培養基為:蔗糖3.75 g/L、尿素11.25 g/L、K2HPO42 g/L、(NH4)2SO41 g/L、MnSO40.25 g/L、豬油乳化液體積分數2.5%。其最佳產酶發酵條件為:初始pH9、培養溫度30 ℃、裝樣量20 mL/250 mL、接種量3%、發酵時間20 h。總之,該菌株能夠利用蔗糖、尿素和豬油等廉價易得的培養基原料達到最佳產酶效果。
從底物、溫度、pH、金屬離子、EDTA、有機溶劑等方面研究了Burkholderiasp. JXJ-16所產脂肪酶粗酶的酶學性質,其特異性水解中鏈pNP酯,最適底物為對硝基苯酚辛酸酯;在35 ℃、pH8.5~9.0時酶活力最高,且在溫度(30~60 ℃)和pH(5.0~10.5)具有較好的穩定性;Na+、Mg2+、Ca2+、Mn2+、EDTA對酶活力具有激活作用,Zn2+、Cu2+、Fe3+對酶活力具有抑制作用,K+對酶活力沒有顯著性影響;除乙醇和乙腈外,粗酶在一定體積分數的異丙醇、甲醇、丙酮、乙酸乙酯、三氯甲烷、二甲苯和正己烷中具有良好耐受性,且處于激活狀態。總之,JXJ-16脂肪酶為低溫堿性脂肪酶,具有較好的溫度和pH穩定性,對多數供試有機溶劑具有良好耐受性。
[1]Peng R,Lin J,Wei D. Purification and characterization of an organic solvent-tolerant lipase fromPseudomonasaeruginosaCS-2[J]. Applied Biochemistry and Biotechnology,2010,162(3):733-743.
[2]查代明,閆云君. 細菌脂肪酶基因表達調控的研究進展[J]. 微生物學報,2015,55(11):1378-1384.
[3]查代明,張炳火,李漢全,等. 假單胞菌屬脂肪酶的分子生物學研究進展[J]. 中國生物工程雜志,2015,35(9):114-121.
[4]Zha D,Xu L,Zhang H,et al. Molecular identification of lipase LipA fromPseudomonasprotegensPf-5 and characterization of two whole-cell biocatalysts Pf-5 and Top10lipA[J]. Journal of Microbiology and Biotechnology,2014,24(5):619-628.
[5]Jaeger K E,Eggert T. Lipases for biotechnology[J]. Current Opinion in Biotechnology,2002,13(4):390-397.
[6]Hasan F,Shah A A,Hameed A. Industrial applications of microbial lipases[J]. Enzyme and Microbial Technology,2006,39(2):235-251.
[7]Khan M,Jithesh K. Expression and purification of organic solvent stable lipase from soil metagenomic library[J]. World Journal of Microbiology and Biotechnology,2012,28(6):2417-2424.
[8]Fan X,Liu X,Wang K,et al. Highly soluble expression and molecular characterization of an organic solvent-stable and thermotolerant lipase originating from the metagenome[J]. Journal of Molecular Catalysis B:Enzymatic,2011,72(3):319-326.
[9]李俊峰,李紅芳,段效輝,等. 耐有機溶劑脂肪酶產生菌的篩選及其粗酶酶學性質[J]. 食品科學,2012(3):116-120.
[10]Xie C,Wu B,Song Q,et al. A lipase with broad solvent stability fromBurkholderiacepaciaRQ3:isolation,characteristics and application for chiral resolution of 1-phenylethanol[J].Bioprocess and Biosystems Engineering,2015,39(1):1-8.
[11]于玉鳳,陸兆新,汪瑾,等. 高效產脂肪酶菌株BurkholderiacepaciaC1產酶條件優化[J]. 食品科學,2010,31(17):218-223.
[12]Yang W,He Y,Xu L,et al. A new extracellular thermo-solvent-stable lipase fromBurkholderiaubonensisSL-4:Identification,characterization and application for biodiesel production[J]. Journal of Molecular Catalysis B:Enzymatic,2016,126:76-89.
[13]Nichols D,Bowman J,Sanderson K,et al. Developments with Antarctic microorganisms:culture collections,bioactivity screening,taxonomy,PUFA production and cold-adapted enzymes[J]. Current Opinion in Biotechnology,1999,10(3):240-246.
[14]梁秋艷. 低溫脂肪酶產生菌株的篩選,發酵條件優化及其酶學性質研究[D]. 石河子:石河子大學,2014:7-9.
[15]Shu Z,Wu J,Cheng L,et al. Production and characteristics of the whole-cell lipase from organic solvent tolerantBurkholderiasp. ZYB002[J]. Applied Biochemistry and Biotechnology,2012,166(3):536-548.
[16]Yao C,Cao Y,Wu S,et al. An organic solvent and thermally stable lipase fromBurkholderiaambifariaYCJ01:Purification,characteristics and application for chiral resolution of mandelic acid[J]. Journal of Molecular Catalysis B:Enzymatic,2013,85-86:105-110.
[17]Xie C,Wu B,Qin S,et al. A lipase with broad solvent stability fromBurkholderiacepaciaRQ3:isolation,characteristics and application for chiral resolution of 1-phenylethanol[J]. Bioprocess and Biosystems Engineering,2016,39(1):59-66.
[18]Kim K R,Kwon D Y,Yoon S H,et al. Purification,refolding,and characterization of recombinantPseudomonasfluorescenslipase[J]. Protein Expression and Purification,2005,39(1):124-129.
[19]Zheng X,Chu X,Zhang W,et al. A novel cold-adapted lipase fromAcinetobactersp. XMZ-26:gene cloning and characterisation[J]. Applied Microbiology and Biotechnology,2011,90(3):971-980.
Optimization of enzyme production conditions and enzymatic properties of cold-adapted and organic solvent-tolerant lipase fromBurkholderiasp. JXJ-16
XIE Yu-ting,ZHA Dai-ming*,SHI Hong-qiu,ZHANG Bing-huo,LI Han-quan
(School of Pharmacy and Life Sciences,Jiujiang University,Jiujiang 332000,China)
Enzyme production conditions forBurkholderiasp. JXJ-16 producing cold-adapted and organic solvent-tolerant lipase were optimized by signal factor analysis to maximize lipase production. A maximum lipase production was obtained after 20 h fermentation of 20 mL of medium containing sucrose 3.75 g/L,urea 11.25 g/L,K2HPO42 g/L,(NH4)2SO41 g/L,MnSO40.25 g/L,and lard emulsion 2.5%(v/v)at an initial pH9.0 in 250 mL Erlenmeyer flask with 3% inoculum at 30 ℃. The crude JXJ-16 lipase preferentially catalyzed thep-nitrophenyl esters of different fatty acids with medium chains,and its best substrate wasp-nitrophenyl caprylate. The optimal reaction temperature for the crude lipase was 35 ℃,and it remained better stability at temperatures ranging from 30 to 60 ℃. The optimum hydrolytic activity of the crude lipase was observed at pH8.5 to 9.0,and it exhibited better stability at pH5.0 to 10.5. The activity of the crude lipase was activated by Na+,Mg2+,Ca2+,Mn2+,and EDTA,respectively,whereas inhibited by Zn2+,Cu2+and Fe3+,respectively. Moreover,K+had no remarkable effect on its activity. The crude lipase had high tolerance and enhanced activity in the presence of different concentration of isopropyl alcohol,methanol,acetone,ethyl acetate,chloroform,benzene,and n-hexane,respectively,except ethanol and acetonitrile. In a word,the strain JXJ-16 could utilize cheap raw materials as a fermentation medium to achieve the optimal lipase production,and its cold-adapted alkaline lipase had better temperature and pH stability and showed high tolerance to most of the tested organic solvents.
cold-adapted lipase;Burkholderia;organic solvent tolerance
2016-07-05
謝玉婷(1995-),女,大學本科,研究方向:微生物脂肪酶資源,E-mail:retar1995xyt@163.com。
*通訊作者:查代明(1985-),男,博士,研究方向:微生物脂肪酶資源及基因表達調控,E-mail:dmzha2015@126.com。
九江學院博士啟動基金(8879416)。
TS201.3
A
:1002-0306(2017)04-0207-07
10.13386/j.issn1002-0306.2017.04.031
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
