西藏靈菇中產(chǎn)胞外多糖假腸膜明串珠菌發(fā)酵性能及流變學(xué)特性研究
2017-03-14 09:05:44楊希娟
食品工業(yè)科技 2017年4期
楊希娟,張 杰,黨 斌,*
(1.青海大學(xué)農(nóng)林科學(xué)院,青海西寧 810016;2.青海省農(nóng)林科學(xué)院,青海省青藏高原農(nóng)產(chǎn)品加工重點實驗室,青海西寧 810016)
西藏靈菇中產(chǎn)胞外多糖假腸膜明串珠菌發(fā)酵性能及流變學(xué)特性研究
楊希娟1,2,張 杰1,2,黨 斌1,2,*
(1.青海大學(xué)農(nóng)林科學(xué)院,青海西寧 810016;2.青海省農(nóng)林科學(xué)院,青海省青藏高原農(nóng)產(chǎn)品加工重點實驗室,青海西寧 810016)
以分離自西藏靈菇發(fā)酵液的3株產(chǎn)胞外多糖的假腸膜明串珠菌為研究對象,對其產(chǎn)胞外多糖能力、發(fā)酵性能及流變學(xué)特性進行了研究。結(jié)果表明:本研究分離得到的3株假腸膜明串珠菌均具有較高的產(chǎn)孢外多糖的能力,菌株R5的胞外多糖產(chǎn)量最高,達到454.67 mg/L;三株菌生長過程符合細菌生長典型規(guī)律,產(chǎn)酸時期主要在菌株的對數(shù)生長期,適于發(fā)酵乳制品生產(chǎn),其中菌株R5發(fā)酵酸乳的組織狀態(tài)、風(fēng)味的感官評分為86分,明顯優(yōu)于其他兩株菌。流變學(xué)特性表明,三株菌制備的發(fā)酵乳的表觀黏度都隨剪切時間的延長而降低,呈現(xiàn)剪切稀釋的流動特征,黏度大小依次為R5>R2>R1;均能夠形成觸變環(huán),為正觸變性流體,R1發(fā)酵乳與R5發(fā)酵乳觸變環(huán)面積相近,分別為2301.72、2924.09 1/s Pa,較R2發(fā)酵乳(4697.82 1/s Pa)小;三株菌制備的發(fā)酵乳的G′值(彈性模量)都高于G″值(粘性模量),均是彈性模量占優(yōu)勢,表現(xiàn)出類固體特性,菌株R5發(fā)酵出的酸乳具有較高的彈性和黏性。通過比較三株菌的發(fā)酵性能與流變學(xué)特性,表明菌株R5相較于其他兩株菌具有較強的產(chǎn)胞外多糖的能力,較高的表觀黏度,較好的粘彈性,結(jié)構(gòu)恢復(fù)能力較強,發(fā)酵的酸乳具有更好的組織結(jié)構(gòu),具有一定的應(yīng)用潛力。
西藏靈菇,胞外多糖,假腸膜明串珠菌,發(fā)酵性能,流變學(xué)特性
西藏靈菇是乳酸菌、酵母菌和醋酸菌共生而成的不規(guī)則粒狀混合物,其形狀與開菲爾粒(Kefir grains)極其相似[1]。西藏靈菇菌粒在生長過程中會產(chǎn)生胞外多糖[2],鑒于胞外多糖產(chǎn)生菌發(fā)酵的酸奶能明顯改善酸奶的組織狀態(tài)和穩(wěn)定性[3-4],因此篩選和分離產(chǎn)胞外多糖乳酸菌成為現(xiàn)在發(fā)酵乳領(lǐng)域研究的熱點。研究發(fā)現(xiàn)從西藏靈菇中分離的乳酸菌在不同的培養(yǎng)基中可以分泌胞外多糖,但其胞外多糖產(chǎn)生菌的種類存在明顯的地域性差異[5-8]。目前西藏靈菇中產(chǎn)胞外多糖乳酸菌的研究主要集中在嗜熱鏈球菌、馬乳酒樣乳桿菌和植物乳桿菌[9-12]。還未見分離自西藏靈菇中且能產(chǎn)胞外多糖的假腸膜明串珠菌的相關(guān)研究。
本課題在前期研究中,從西藏靈菇發(fā)酵乳中分離得到了產(chǎn)孢外多糖的三株假腸膜明串珠菌,且三株菌均具有發(fā)酵凝乳的特性。但是這三株菌的發(fā)酵特性是否存在差異,是否具有應(yīng)用前景仍不清楚。食品流變學(xué)作為一種研究在外力作用下食品材料變形情況的科學(xué),可研究發(fā)酵型酸乳流體的流動、變形隨時間和力的變化而表現(xiàn)出的性質(zhì),對評價不同菌株發(fā)酵酸乳的質(zhì)地、黏彈性、穩(wěn)定性方面具有重要作用[13]。因此本文在前期研究的基礎(chǔ)上,從發(fā)酵性能及流變學(xué)特性方面全面評價分離自西藏靈菇中產(chǎn)胞外多糖的三株假腸膜明串珠菌發(fā)酵酸乳的品質(zhì),篩選出具有應(yīng)用價值的菌株,為其開發(fā)與應(yīng)用提供理論依據(jù)。
1 材料與方法
1.1 材料與儀器
乳酸菌R1、R2、R5 由青海省青藏高原農(nóng)產(chǎn)品加工重點實驗室分離西藏靈菇乳得到,經(jīng)鑒定均為假腸膜明串珠菌;脫脂乳培養(yǎng)基 新鮮牛乳在4000 r/min下離心10 min,每100 mL分裝于250 mL的錐形瓶中,105 ℃下滅菌20 min;MRS瓊脂培養(yǎng)基 蛋白胨10 g、牛肉膏10 g、酵母提取物5 g、K2HPO42 g、檸檬酸二銨2 g、乙酸鈉5 g、葡萄糖20 g、吐溫80 1 mL、MgSO4·7H2O 0.58 g、MnSO4·4H2O 0.25 g、瓊脂15 g、蒸餾水1 L,調(diào)節(jié)pH到6.2~6.4,121 ℃下滅菌30 min;鮮牛乳 采自青海省良種繁殖場;其他試劑 均為分析純。
DHR流變儀 美國TA儀器;pHS-3C型精密酸度計 上海儀電科學(xué)儀器股份有限公司;LDZX-75KBS立式壓力蒸汽滅菌器 上海申安醫(yī)療器械廠;SW-CJ-2D型雙人凈化工作臺 蘇州凈化設(shè)備有限公司;AL204萬分之一分析天平 梅特勒-托利多儀器(上海)有限公司。
1.2 實驗方法
1.2.1 菌種斜面培養(yǎng) 將保藏的菌種懸液轉(zhuǎn)接至MRS瓊脂斜面培養(yǎng)基中,37 ℃培養(yǎng)24 h,培養(yǎng)3次。
1.2.2 菌種活化 用接種環(huán)挑取斜面菌種兩環(huán),接種于裝有100 mL MRS液體培養(yǎng)基的250 mL三角瓶中,37 ℃培養(yǎng)24 h進行活化。
1.2.3 菌種胞外多糖的提取及多糖含量測定 假腸膜明串珠菌多糖的提取參考文獻[14-15]稍作修改。三株菌株以5%的接種量分別接入脫脂乳培養(yǎng)基中,在25 ℃下靜置培養(yǎng)24 h,無菌條件下濾去菌粒,得到的發(fā)酵乳沸水浴加熱15 min,冷卻后離心(20 min,4000 r/min,4 ℃),上清液中加入80%(w/v)的三氯乙酸至終濃度4%(w/v),4 ℃冰箱靜置過夜后離心(20 min,8000 r/min,4 ℃),上清液中添加95%(v/v)乙醇至終濃度75%(v/v),4 ℃靜置22 h,離心(20 min,8000 r/min,4 ℃)后取沉淀物,加入樣品體積1/5的Sevage試劑除蛋白,濃縮液透析48 h,減壓濃縮后冷凍干燥得到西藏靈菇的胞外多糖。
胞外多糖含量的測定:苯酚-硫酸法[16]。以葡萄糖為標準品制作標準曲線。根據(jù)標準曲線可得回歸方程:y=0.0145x-0.0173,R2=0.9954。式中:y為樣品吸光度A;x為樣品中胞外多糖質(zhì)量濃度(mg/L)。
1.2.4 三株假腸膜明串珠菌的生長特性研究 將三株活化后的菌種以體積分數(shù)5%的接種量接入MRS液體培養(yǎng)基,37 ℃培養(yǎng)27 h,每3 h取樣,測定其OD600值及pH的變化[17]。
1.2.5 三株假腸膜明串珠菌在牛乳中發(fā)酵性能測定 發(fā)酵劑的制備:取3環(huán)乳酸菌穿刺保藏菌種接種于裝有5 mL滅菌脫脂乳試管中,在37 ℃培養(yǎng)活化至凝乳,再按5%接種量接入100 mL滅菌脫脂乳中,于37 ℃培養(yǎng)至牛乳凝固時即為乳酸菌發(fā)酵劑。再以5%接種量接入滅菌牛乳中發(fā)酵,至牛乳凝固,4 ℃冷藏24 h得成品,測定三株菌發(fā)酵乳成品的凝乳時間、pH、滴定酸度及感官品質(zhì)評定,比較三株菌的發(fā)酵特性[18]。
1.2.5.1 發(fā)酵凝乳時間的測定 一般以肉眼觀察乳變粘稠,呈凝膠狀態(tài),即已達到發(fā)酵終點。每1 h觀察一次,記錄培養(yǎng)至乳凝固的時間[19]。
1.2.5.2 酸度的測定 采用0.1 mol/L NaOH滴定法測定發(fā)酵乳的酸度[20-21]。
1.2.5.3 感官評定 根據(jù)三株菌發(fā)酵產(chǎn)物的色澤(顏色分布均勻、有無雜色)、組織狀態(tài)(乳清析出多少,凝乳是否細膩均勻)和滋味氣味(包括口感滑潤剎口、醇香、酯香、酸度情況以及有無澀味、苦味情況)對發(fā)酵乳評級打分,總分100分,品嘗人員8名,取平均值。其中口感40分,組織狀態(tài)30分,香氣滋味30分,評分標準見表1[19]。
1.2.6 流變學(xué)特性測定
1.2.6.1 表觀黏度的測定 參考文獻[22]的方法并稍作修改。在恒溫4 ℃,設(shè)定剪切速率范圍為0.1~100 s-1,椎板(40 mm,1°),連續(xù)測量30個數(shù)據(jù)點,檢測樣品的表觀黏度隨剪切時間的變化情況,測定時間為300 s。
1.2.6.2 觸變特性檢測 參照文獻[22]的方法并稍作修改。在4 ℃條件下檢測樣品剪切應(yīng)力隨剪切速率變化的情況:首先轉(zhuǎn)子的剪切率由0 s-1線性升高到600 s-1,采集的數(shù)據(jù)點為30個,測試時間150 s;到達600 s-1后再線性降速到0 s-1,采集的數(shù)據(jù)點為30個,測試時間150 s,椎板(40 mm,1°)。

表1 樣品感官評定標準Table 1 Sensory evaluation of samples
1.2.6.3 粘彈性檢測 參照文獻[22]的方法并稍作修改。在4 ℃條件下,椎板(40 mm,1°),應(yīng)變?yōu)?%,頻率由0.1 Hz變化到10 Hz,對發(fā)酵乳樣品進行應(yīng)變掃描,采集的數(shù)據(jù)點為30個。
1.3 數(shù)據(jù)處理
所有實驗均重復(fù)3次。數(shù)據(jù)用DPS 6.5進行方差分析和多重比較,以p<0.05為顯著性檢驗標準。
2 結(jié)果與分析
2.1 三株假腸膜明串珠菌發(fā)酵產(chǎn)胞外多糖的含量分析
三株分離自西藏靈菇乳中的假腸膜明串珠菌的胞外多糖產(chǎn)量見圖1,三株菌胞外多糖的產(chǎn)量存在顯著差異(F0.05),其中R5的胞外多糖產(chǎn)量最高,可達到454.67 mg/L,R2次之,R1的胞外多糖產(chǎn)量最低,只有332.54 mg/L。但本研究供試菌株的多糖產(chǎn)量明顯高于李達等人[23]報道的分離自西藏靈菇的嗜熱鏈球菌產(chǎn)孢外多糖的含量(140~162 mg/L)和曹永強[24]等人報道的分離自西藏靈菇中植物乳桿菌產(chǎn)孢外多糖的含量(131.26 mg/L)。說明不同來源的西藏靈菇的菌群構(gòu)成及菌株的特性具有明顯的差異性,本研究分離得到的三株假腸膜明串珠菌具有更高的產(chǎn)孢外多糖的能力,有一定的應(yīng)用前景。
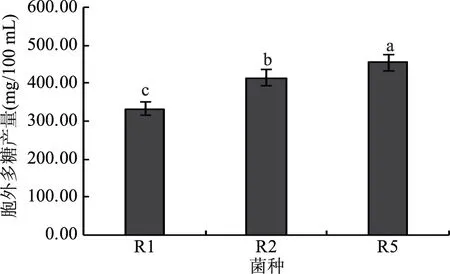
圖1 三株假腸膜明串珠菌胞外多糖產(chǎn)量Fig.1 The exopolysaccharide production of three strains
2.2 三株假腸膜明串珠菌生長特性研究
2.2.1 三株假腸膜明串珠菌生長曲線 三株假腸膜明串珠菌生長曲線見圖2。由圖2可見,R1、R2、R5生長規(guī)律基本一致,R1菌株0~3 h處于生長延滯期,在3~6 h菌數(shù)增加較快,6 h后達到對數(shù)生長期,12 h后進入生長平穩(wěn)期;R2菌株無明顯的生長延滯期,在3~9 h時,處于對數(shù)生長期,活菌數(shù)增加迅速,在12 h時活菌數(shù)達到一個峰值,12 h后進入生長穩(wěn)定期,21 h后活菌數(shù)稍有下降;R5菌株在0~3 h期間生長較緩,3~12 h生長較快,12 h以后生長逐漸趨于平穩(wěn),可見這三株菌活菌數(shù)的變化符合細菌生長典型規(guī)律。從整個生長過程來看,R2菌株在對數(shù)生長期生長較快,其次為R5,穩(wěn)定生長期R2和R1菌株的活菌數(shù)較為接近,高于R5的活菌數(shù)。
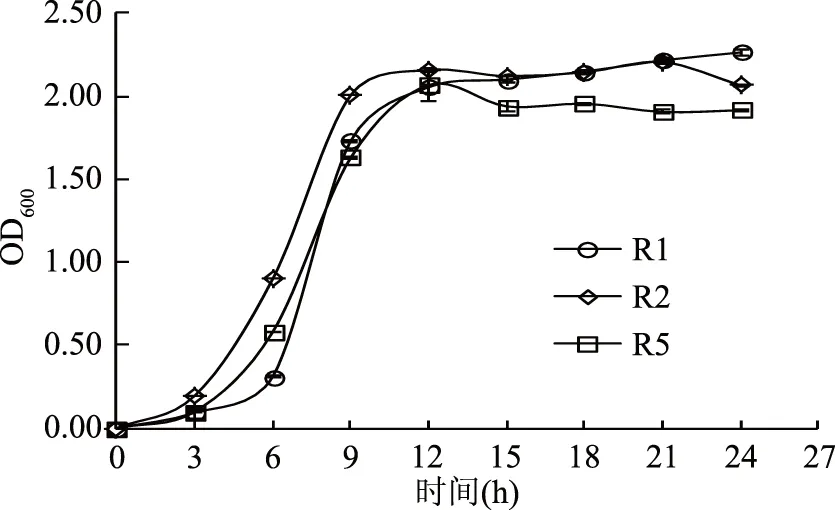
圖2 三株菌生長曲線Fig.2 The growth chart of three strains
2.2.2 三株假腸膜明串珠菌發(fā)酵過程中pH的變化 發(fā)酵乳pH的變化是由于菌株發(fā)酵過程中產(chǎn)酸而引起,在一定程度上可以反映菌株產(chǎn)酸量的多少。三株菌在不同發(fā)酵時間的pH變化趨勢見圖3。由圖3可知,三株菌pH均隨發(fā)酵時間的延長而下降,但是產(chǎn)酸的速度和時間有差異。其中R1菌株的pH在0~3 h期間下降較為緩慢,3~9 h下降速度較快,9 h時達到4.55,之后逐漸緩慢,趨于穩(wěn)定;R2菌株pH在6 h之前下降迅速,達到4.38,之后趨于平緩;R5菌株pH在0~6 h期間下降較為迅速達到4.63,6~12 h下降較為緩慢,12 h后趨于平緩。發(fā)酵前12 h,R2發(fā)酵液pH下降較快,產(chǎn)酸明顯高于R1、R5,在發(fā)酵12 h時pH均達到了4.29,12 h后三株菌產(chǎn)酸基本趨于穩(wěn)定。由此可見這三株菌的產(chǎn)酸時期主要在菌株的對數(shù)生長期,適于發(fā)酵乳制品生產(chǎn),并利于提高生產(chǎn)效率。
2.2 三株假腸膜明串珠菌發(fā)酵性能的比較

表2 三株假腸膜明串珠菌發(fā)酵性能比較Table 2 The comparison in fermentation performance of three strains
注:同一列數(shù)據(jù)后標不同字母表示差異顯著(p<0.05)。
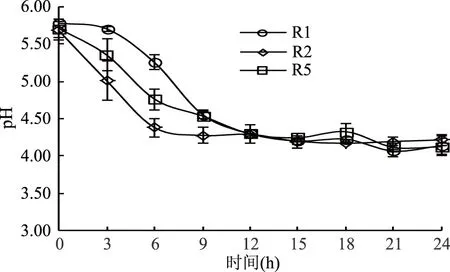
圖3 三株菌發(fā)酵過程中pH變化曲線Fig.3 The pH change chart of three strains in fermentation course
由表2可知,三個乳酸菌中,菌株R2的凝乳時間最短,產(chǎn)酸最快,菌株R5次之,菌株R1稍慢。從組織狀態(tài)和風(fēng)味來考察,菌株R5凝乳的組織狀態(tài)和風(fēng)味均較好,R1的凝乳風(fēng)味較好,組織狀態(tài)較好。R2有較多的乳清析出,凝乳不均勻,且風(fēng)味帶有異味。綜合評分評價各菌種的優(yōu)缺點,菌株R5發(fā)酵性能優(yōu)于其他菌株。這可能與R5菌株孢外多糖產(chǎn)量較高有關(guān),胞外多糖可以賦予發(fā)酵乳特殊的風(fēng)味質(zhì)構(gòu),可影響發(fā)酵乳的黏性和組織結(jié)構(gòu),減少乳清析出[25]。
2.3 三株假腸膜明串珠菌發(fā)酵酸奶的流變特性
2.3.1 恒溫變速條件下三株菌發(fā)酵乳表觀粘度隨時間的變化 表觀粘度是影響酸奶感官品質(zhì)的一個重要參數(shù)[22]。在0~100 s-1的剪切速率下,測定了三株菌發(fā)酵酸乳的表觀粘度,結(jié)果如圖4所示。三株菌發(fā)酵酸乳的表觀黏度都隨剪切時間的延長而降低,這與王松松等[23]研究的恒溫恒速條件下發(fā)酵乳制品表觀黏度變化趨勢相似,但黏度明顯高于其報道,說明本研究中產(chǎn)多糖的三株菌發(fā)酵的酸奶具有更穩(wěn)定的黏度和組織結(jié)構(gòu)。0~150 s,三株菌發(fā)酵酸乳均呈現(xiàn)剪切稀釋的流動特征,三株菌在剪切外力的作用下,經(jīng)過一段時間后,由于外力大于體系內(nèi)部由某種懸浮液中粒子所形成的結(jié)構(gòu)被破壞,黏度隨剪切時間下降,表現(xiàn)出具有時間因素的切稀現(xiàn)象[22]。150 s之后,隨著剪切時間的延長,剪切速率不斷增加,三株菌發(fā)酵酸乳樣品均表現(xiàn)為恒定的理想牛頓流體行為。這種特性對于乳品加工具有重要意義,在攪拌過程中,隨著攪拌速度的增加,乳品的表觀黏度下降;而當(dāng)攪拌速度減慢直至停止時,乳品的黏度增加,這更有利于凝固性發(fā)酵乳的加工生產(chǎn)[26]。三株菌發(fā)酵酸奶的黏度大小依次為:R5>R2>R1,這與三株菌胞外多糖產(chǎn)量的大小順序一致,說明胞外多糖產(chǎn)量較高的菌株具有相對較高的表觀黏度。
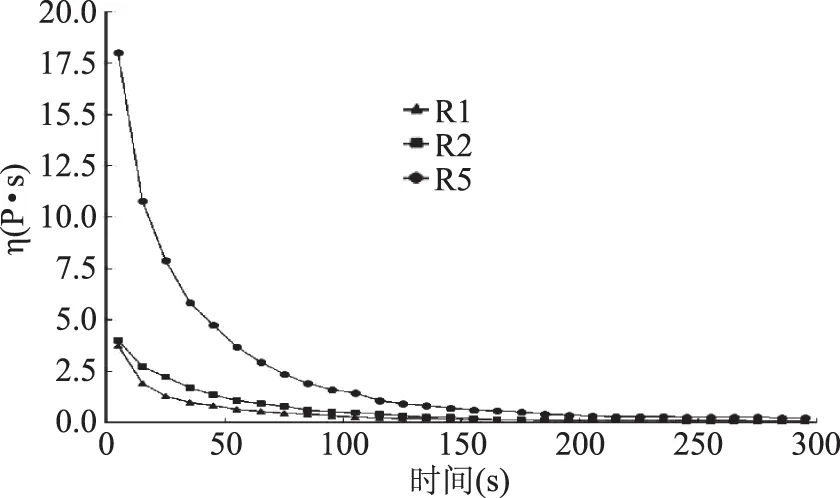
圖4 恒溫變速條件下三株菌發(fā)酵乳樣品表觀黏度隨剪切時間變化曲線Fig.4 Change curves of apparent viscosity with shear time at constant temperature and variable speed for fermented dairy products from three strains
2.3.2 三株菌發(fā)酵乳樣品的剪切應(yīng)力隨剪切速率的變化 觸變性是指某體系在攪動或其他機械作用下,其黏度或剪切應(yīng)力隨時間變化的一種流變現(xiàn)象。在外切力作用下,若體系的黏度隨剪切時間的延長而下降,靜止后又恢復(fù),即具有時間因素的切稀現(xiàn)象,稱為正觸變性;反之,若體系的黏度上升,靜止后又恢復(fù),即具有時間因素的切稠現(xiàn)象,稱為負觸變性。當(dāng)一個特定體系可先后出現(xiàn)正觸變性和負觸變性特征,稱之為復(fù)合觸變性[22]。
三株菌發(fā)酵乳樣品在恒溫條件下的剪切應(yīng)力隨剪切速率的變化情況(即觸變性)的測定結(jié)果見圖5。由圖5可知,三株菌發(fā)酵乳樣品均能夠形成觸變環(huán),說明此三株菌發(fā)酵乳制品都是黏彈性和正觸變性流體[27-28]。三株菌株制備的發(fā)酵乳的觸變環(huán)形狀相似,但是R2發(fā)酵乳的觸變環(huán)的變化幅度較大,R5和R1變化幅度較小。
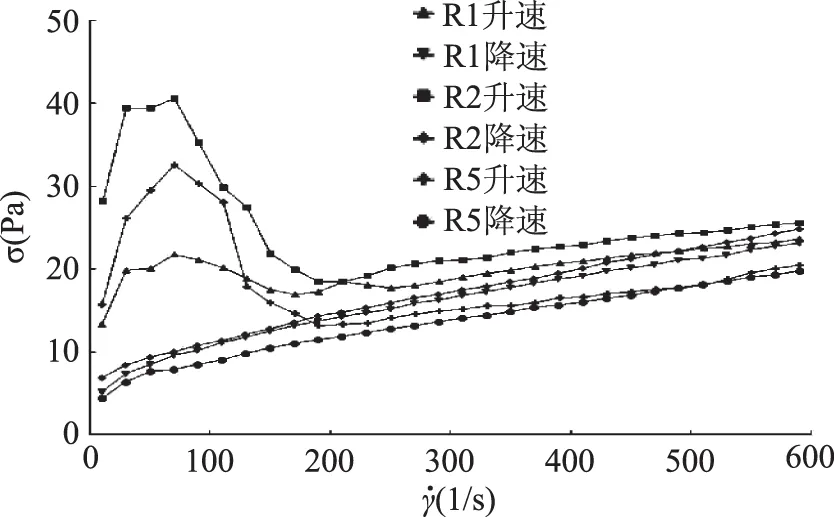
圖5 三株菌發(fā)酵乳樣品剪切應(yīng)力隨剪切速率變化曲線Fig.5 Curves of shear stress with shear rate at constant temperature and variable speed for fermented dairy products from three strains
在觸變性實驗中得到的滯后環(huán)其面積的大小可以代表樣品觸變性的情況,面積越大說明此樣品的結(jié)構(gòu)恢復(fù)速度越慢,越小則恢復(fù)速度越快。R1發(fā)酵乳、R2發(fā)酵乳和R5發(fā)酵乳的觸變環(huán)面積(升速曲線下方面積-降速曲線下方面積)分別為2301.72(1/s Pa)、4697.82(1/s Pa)、2924.09(1/s Pa)。從觸變環(huán)面積可以看出R1發(fā)酵乳與R5發(fā)酵乳觸變環(huán)面積相近,較R2發(fā)酵乳小,說明菌株R1、R5發(fā)酵出的酸奶結(jié)構(gòu)恢復(fù)速度快,發(fā)酵性能好。
2.3.3 三株菌發(fā)酵乳樣品的粘彈性 在應(yīng)變掃描過程中可以得到兩組數(shù)據(jù),它們分別為G′(彈性模量,儲能模量)和G″(黏性模量,損耗模量),G′表示樣品的彈性模量(儲存模量),是衡量樣品抵抗彈性變形能力大小的尺度,反映酸奶凝膠特性的不同。G″表示樣品的粘性模量(損耗模量),反映了樣品粘性的大小[29]。當(dāng)G′

圖6 三株菌發(fā)酵乳樣品彈性模量G′和黏性模量G″隨頻率變化曲線Fig.6 Curves of elastic modulus and viscous modulus with frequency for fermented dairy products from three strains
3 結(jié)論
3.1 本研究分離得到的三株假腸膜明串珠菌均具有較高的產(chǎn)孢外多糖的能力,三株菌活菌數(shù)的變化符合細菌生長典型規(guī)律,產(chǎn)酸時期主要在菌株的對數(shù)生長期,適于發(fā)酵乳制品生產(chǎn),其中菌株R5的胞外多糖產(chǎn)量及發(fā)酵特性明顯優(yōu)于其他兩株菌,具有一定的應(yīng)用價值。
3.2 三株菌的流變學(xué)特性表明,三株菌制備的發(fā)酵乳表現(xiàn)出典型的非牛頓假塑性流體特性,均為正觸變流體,具有一定的粘彈性。菌株R5發(fā)酵制備的酸乳相對于其他兩株菌具有相對較高的表觀黏度,較好的粘彈性,結(jié)構(gòu)恢復(fù)能力較強,發(fā)酵的酸乳具有更好的組織結(jié)構(gòu),具有一定的應(yīng)用潛力。
[1]楊希娟,師俊玲,樊明濤.西藏靈菇牛乳發(fā)酵液中的菌相與物質(zhì)變化[J].中國食品學(xué)報,2007,7(4):42-46.
[2]Zhou J Z,Liu X L,Jiang H H,et al.Analysis of the microflora in Tibetan kefir grains using denaturing gradient gel electrophoresis[J]. Food Microbiology,2009,26(8):770-775.
[3]Ruas-Madiedox S,Hugenholtz J,Zoon P. An overview of the functionality of exopolysaccharides produced by lactic acid bacteria[J]. International Dairy Journal,2002,12(2/3):163-171.
[4]劉慧,熊利霞,易欣欣,等. 藏靈菇中高產(chǎn)胞外多糖乳酸菌的篩選及其發(fā)酵性能的研究[J]. 食品科學(xué),2007,28(5):211-215.
[5]劉變芳,孔慶學(xué),郭藹光. 自然發(fā)酵劑“西藏雪蓮”的初步研究與菌種鑒定[J].中國釀造,2004,23(11):11-13.
[6]劉宇峰,王金英,曲曉軍,等.西藏靈菇菌的菌相菌學(xué)的研究[J].中國乳品工業(yè),2005,33(9):35-39.
[7]楊希娟,樊明濤,師俊玲,等. 西藏靈菇發(fā)酵乳中優(yōu)勢菌群的分離鑒定[J].中國釀造,2007,26(6):52-55.
[8]董健,陳歷俊,姜鐵民,等. 西藏靈菇顆粒中菌種的分離鑒定[J].食品科技,2015,40(1):6-9.
[9]Ahmed Z,Wang Y P,Anjum N,et al. Characterization of exopolysaccharide produced byLactobacilluskefiranofaciensZW3 isolated from Tibet kefir-Part II[J]. Food Hydrocolloids,2013,30(1):343-350.
[10]Jiang S J,Qian F,Ren X H,et al. Studies on the preliminary characterization of a novel exopolysaccharide produced bystreptococcusthermophilusstrain from Tibetan kefir grain[J]. Advanced Materials Research,2013,690/693:1374-1377.
[11]Qin Q Q,Xia B S,Xiong Y,et al. Structural characterization of the exopolysaccharide produced by Streptococcus thermophilus 05-34 and its in situ application in yogurt[J]. Journal of Food Science,2011,76(9):C1226-C1230.
[12]Wang Y P,Li C,Liu P,et al. Physical characterization of exopoloysaccharide produced byLactobacillusplantarumKF5 isolated from Tibet kefir[J]. Carbohydrate Polymers,2010,82(3):895-903.
[13]徐夢夢,孟岳成.熱殺菌處理對攪拌型酸奶流變性質(zhì)影響的研究[J].食品科技,2013,38(5):62-68.
[14]呂曉鵬. 黑果枸杞多糖的分離純化與結(jié)構(gòu)表征研究[D]. 西安:西北大學(xué),2012.
[15]Wang Y P,Ahmed Z,Wu F,et al. Physicochemical properties of exopolysaccharide produced byLactobacilluskefiranofaciensZW3 isolated from Tibet kefir[J]. International Journal of Biological Macromolecules,2008,43(3):283-288.
[16]梁雪. 產(chǎn)胞外多糖酵母菌株的篩選鑒定及多糖結(jié)構(gòu)初步分析[D].哈爾濱:東北林業(yè)大學(xué),2010.
[17]王剛,劉娟,陳光,等. 產(chǎn)D-乳酸假腸膜明串珠菌生長特性分析[J].湖北農(nóng)業(yè)科學(xué),2015,55(5):1239-1241.
[18]王玉華,鄭明珠,關(guān)曉玲,等.2株風(fēng)味乳酸菌的篩選及其生長特性的研究[J].中國乳品工業(yè),2005,33(11):7-10.
[19]劉慧,李蘭平,熊利霞,等. 功能性開菲爾酸奶復(fù)合發(fā)酵劑的研制[J].食品科學(xué),2005,26(12):139-143.
[20]Guizel-Seydim Z B,Seydim A C,Greene A K,et al. Determination of Organic Acids and Volatile Flavor Substances in Kefir during Fermentation[J]. Journal of Food Composition and Analysis,2000(13):35-43.
[21]王叔淳.食品衛(wèi)生檢驗技術(shù)手冊(第3版)[M].北京:化學(xué)工業(yè)出版社,2002:506.
[22]王松松,陳慶森. 3種發(fā)酵乳制品流變性質(zhì)的比較與分析[J].食品科學(xué),2011,32(19):7-11.
[23]李達,張雪,張莉.西藏靈菇中產(chǎn)胞外多糖嗜熱鏈球菌的分離篩選及其發(fā)酵性能測定[J].食品科學(xué),2011,32(13):225-228.
[24]曹永強,王輯,趙笑,等. 植物乳桿菌 YW11 生產(chǎn)胞外多糖的發(fā)酵條件研究[J].食品科學(xué)技術(shù)學(xué)報,2016,34(1):42-49.
[25]楊同香,仲玉備,吳孔陽. 高產(chǎn)胞外多糖乳酸菌的篩選及其在發(fā)酵乳中的應(yīng)用[J].食品科技,2016,41(2):26-29.
[26]楊貞耐,張雪. 乳酸菌胞外多糖的流變學(xué)特性和分子結(jié)構(gòu)修飾[J].食品科學(xué),2007,28(12):535-538.
[27]Benezech T,Maingonnat J F. Characterization of the rheological properties of yogurt:a review[J]. Journal of Food Engineering,1994,21:447-472.
[28]Lorenzi L D,Pricl S,Torriano G. Rheological behavior of low-fat and full-fat stirred yogurt[J]. International Dairy Journal,1995,5:661-671.
[29]Osaana N D,Henriksson A,Vasiljevic T,et al. Rheological properties and sensory characteristics of set-type soy yogurt[J]. Journal of Agricultural and Food Chemistry,2007,55(24):9868-9876.
[30]Engkhampam N,Sagis L M C,DE Vries R,et al. Physicochemical properties of pectins from okra(Abelmoschusesculentus(L.)Moench)[J]. Food Hydrocolloids,2010,24(1):35-41.
Fermentability and rheologic properties of three strains of exopolysaccharide-producingLeuconostocpseudomesenteroidesfrom Tibetan kefir
YANG Xi-juan1,2.ZHANG Jie1,2,DANG Bin1,2,*
(1.Academy of Agriculture and Forestry,Qinghai University,Xining 810016,China;2.Qinghai Tibetan Plateau Key Laboratory of Agric-Product Processing,Qinghai Academy of Agriculture and Forestry,Xining 810016,China)
In this study,the exopolysaccharide content,fermentability and rheological properties of three srains with exopolysaccharide produced were evaluated. The results showed thatLeuconostocpeudomesenteroidesof three srains produce exopolysaccharide,exopolysaccharide content of R5 was 454.67 mg/L.The growth process three srains was typical bacterial growth,acid production period mainly was logarithmic phase of strain,they were suitable for fermented dairy production,R5 of them had better organization status and flavor and sensory score was 86. The result of rheological properties showed that apparent viscosity was reduced with the shearing time of fermented dairy products from three strains,they presented flow characteristics of the shear thinning,viscosity size was R5>R2>R1.Fermented milk of three strains could form the thixotropic loop,they were the thixotropic fluid,hixotropic loop area of fermented milk of R1 and R5 was 2301.72(1/s Pa)and 2924.09(1/s Pa). R2 was 4697.82(1/s Pa).G′ values were higher than G"of fermented milk from three strains and elastic modulus was preferred,fermented milk of R5 has higher elastic and viscous. By comparing their fermentability and rheological properties,strain of R5 had higher exopolysaccharide content,its fermented milk revealed the highest apparent viscosity,viscoelasticity and strongest stability,strain of R5 had potential applications.The result illustrated that exopolysaccharide producing strains could give good texture for fermented milk.
Tibetan kefir;exopolysaccharide;Leuconostocpseudomesenteroides;fermentability;rheological properties
2016-08-16
楊希娟(1980-),女,博士研究生,副研究員,研究方向:食品功能化學(xué)與發(fā)酵工程,E-mail:156044169@qq.com。
*通訊作者:黨斌(1980-),男,副研究員,研究方向:食品功能化學(xué)與營養(yǎng),E-mail:dangbin811@tom.com。
國家自然科學(xué)基金項目(31260395)。
TS201.7
A
:1002-0306(2017)04-0106-06
10.13386/j.issn1002-0306.2017.04.012
