特異性信號轉導阻滯劑AG490對人肝癌HepG2細胞增殖、凋亡的影響及機制
2016-12-27 04:51:46趙冀安趙洪正周小慧孫會鳳朱俊青胡毅平
山東醫藥 2016年41期
趙冀安,趙洪正,周小慧,孫會鳳,朱俊青,胡毅平
(河北醫科大學第一醫院,石家莊050013)
?
特異性信號轉導阻滯劑AG490對人肝癌HepG2細胞增殖、凋亡的影響及機制
趙冀安,趙洪正,周小慧,孫會鳳,朱俊青,胡毅平
(河北醫科大學第一醫院,石家莊050013)
目的 探討特異性信號轉導阻滯劑AG490對人肝癌HepG2細胞增殖、凋亡的影響及其作用機制。方法 將培養好的人肝癌細胞株HepG2用4、6、8 mg/kg的AG490(AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組)進行處理,另設對照組(僅加PBS)。AG490干預24 h,MTT法測算細胞增殖抑制率;流式細胞術測算細胞周期及凋亡率;采用Hoechst 33342染色法觀察細胞形態學變化;采用Western blotting法檢測HepG2細胞中P-JAK2、P-STAT3、Bcl-2、Bax、Survivin、Caspase-3蛋白表達。結果 AG490干預24 h,與對照組比較,AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組細胞增殖抑制率、G0/G1期細胞比例、細胞凋亡率逐漸升高(P均<0.05),且呈濃度依賴性(P均<0.05);HepG2細胞核染色質固縮濃集、碎裂,凋亡小體形成,藍色熒光增強,且AG490濃度越高以上變化越明顯;P-JAK2、P-STAT3、Bcl-2、Survivin蛋白表達降低,而Bax、Caspase-3蛋白表達升高(P均<0.05)。結論 AG490可能通過抑制JAK2/STAT3信號通路表達抑制人肝癌HepG2細胞增殖并促進其凋亡。
肝腫瘤;HepG2細胞;Janus激酶2/信號轉導子與轉錄激活子3信號通路;信號轉導阻滯劑AG490
肝癌的發生發展是多基因突變演化進展的過程,發病機制目前仍不明確。Janus激酶2/信號轉導子與轉錄激活子3(JAK2/STAT3)信號通路在細胞增殖、凋亡、分化和免疫功能等過程中發揮重要作用,當該信號通路發生異常活化時會導致細胞出現異常增殖甚至惡變[1]。STAT3作為JAK2/STAT3轉導通路的重要成員,可與相關轉錄因子相互作用,使JAK2/STAT3與其他信號通路相互關聯,在腫瘤發生發展中起重要作用[2,3]。研究證實,多種肝癌細胞內均有STAT3激活,JAK2/STAT3信號通路活化顯著促進了肝癌細胞增殖[4]。STAT3調控的靶基因編碼蛋白包括抗凋亡蛋白Bcl-xl、Bcl-2、Mcl-1、存活蛋白Survivin、促凋亡蛋白Bax及血管內皮生長因子(VEGF)[5]。JAK2/STAT3信號通路的特異性信號轉導阻滯劑AG490可影響肝癌細胞的多種生物學特性(抑制肝癌細胞增殖和誘導凋亡,降低侵襲力等)[6]。2015年8~12月,我們探討了AG490對人肝癌HepG2細胞增殖、凋亡的影響及作用機制。
1 材料與方法
1.1 材料 人肝癌細胞株HepG2(河北醫科大學腫瘤研究所),AG490、MTT(Sigma公司),Annexin V/PI細胞凋亡檢測試劑盒、Hoechst 33342(BioVision公司),P-JAK2、P-STAT3、Bcl-2、Bax、Survivin、Caspase-3和β-actin兔抗人多克隆抗體(Santa公司)。
1.2 細胞培養及分組 將人肝癌細胞株HepG2用含10% FBS的RPMI1640培養基培養,置于5% CO2、37 ℃孵箱中傳代培育。取對數生長期的HepG2細胞,接種于96孔培養板(含10% FBS的RPMI1640培養液稀釋后的懸液),貼壁孵育24 h分別加入不同濃度(4、6、8 mg/kg)的AG490溶液0.2 mL,根據AG490濃度將細胞分為AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組,另設對照組,不加藥物,僅加PBS。
1.3 細胞增殖抑制率測算 采用MTT法。分別用AG490干預8、12、24 h取各組細胞,每孔各加入DMSO溶液100 μL,振蕩5 min,用酶標儀測定490 nm波長的吸光度(A)值。每個實驗組設3個復孔。計算細胞增殖抑制率,細胞增殖抑制率=[(實驗組A-對照組A)/對照組A]×100%。
1.4 細胞周期及凋亡率測算 采用流式細胞術。AG490干預24 h取各組細胞,PBS清洗2次,棄上清液,用PBS洗滌后懸浮,加入Annexin V 5 μL、PI 5 μL,混勻,4 ℃避光反應30 min,用流式細胞儀檢測細胞周期和凋亡率。
1.5 細胞形態學變化觀察 采用Hoechst 33342染色法。AG490干預24 h取各組細胞,吸棄上清液,PBS清洗,每孔加入固定液100 μL,固定10 min。去除固定液,PBS洗2次,滴加Hoechst 33342工作液100 μL,室溫染色10 min。以340 nm波長紫外光激發,熒光顯微鏡下觀察細胞形態學變化。
1.6 HepG2細胞中STAT3及下游相關蛋白表達檢測 采用Western blotting法。AG490干預24 h取各組細胞,去除培養液,PBS洗1次,刮下細胞提取蛋白并定量,取等量蛋白電泳分離,轉移到PVDF膜,轉移膜用1% BSA封閉。隨后加入1∶1 000的兔抗人P-JAK2、P-STAT3、Bcl-2、Bax、Survivin、Caspase-3和β-actin多克隆抗體,4 ℃孵育過夜,用TBST洗3次后再分別加入1∶5 000標記的抗兔二抗,室溫下孵育1 h,TBS洗3次。經ECL化學發光法檢測并記錄各目的條帶與內參條帶灰度值,以目的條帶與內參條帶灰度比值作為各目的蛋白相對表達量。
1.7 統計學方法 采用SPSS13.0統計軟件。計量資料以表示,多組間比較采用單因素方差分析,進一步兩兩比較采用LSD-t檢驗。P<0.05為差異有統計學意義。
2 結果
2.1AG490對HepG2細胞增殖的影響AG490對HepG2細胞增殖抑制作用呈濃度和時間依賴性,各組不同時間細胞增殖抑制率比較見表1。
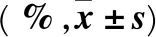
表1 各組不同時間細胞增殖抑制率比較
2.2 AG490對HepG2細胞周期和凋亡率的影響 對照組、AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組G0~G1期細胞百分比分別為(59.48±1.1)%、(64.59±2.6)%、(69.55±3.4)%、(73.28±3.9)%;與對照組比較,AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組G0~G1期細胞逐漸增多(P均<0.05),且呈濃度依賴性(P均<0.05)。對照組、AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組細胞凋亡率分別為(1.50±0.12)%、(33.32±1.16)%、(45.54±1.22)%、(52.22±1.06)%;與對照組比較,AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組細胞凋亡率逐漸升高(P均<0.05),且呈濃度依賴性(P均<0.05)。
2.3 細胞形態學變化 對照組細胞核在熒光顯微鏡下顯示為較弱的均勻藍色熒光,可見細胞核完整且染色均勻。AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組則可見凋亡細胞呈致密濃染,核染色質固縮濃集、碎裂,凋亡小體形成,藍色熒光增強,且AG490濃度越高以上變化越明顯。2.4 AG490對HepG2細胞STAT3及下游相關蛋白表達的影響 與對照組比較,AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組P-STAT3、P-JAK2、Bcl-2蛋白表達降低,Bax、Caspase-3蛋白表達升高(P均<0.05),且呈濃度依賴性(P均<0.05)。見表2。
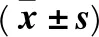
表2 各組STAT3及下游相關蛋白相對表達量比較
3 討論
肝癌的發生發展有眾多信號通路參與[7]。JAK2/STAT3信號通路廣泛參與體內多種重要生長因子信號轉導,參與調節正常細胞的增殖、分化、凋亡、免疫應答等過程[8,9]。STAT3可通過上調Bcl-2、Bcl-xl、Mcl-1、Survivin等抗凋亡基因的表達抑制細胞凋亡;同時抑制基因p53和Bax表達,抑制正常細胞分化增殖和腫瘤細胞凋亡,最終導致肝癌發生[10]。AG490作為酪氨酸激酶抑制劑通過阻斷JAK2激酶活化,使細胞中STAT3表達降低,從而發揮阻滯細胞信號傳導作用。
Yoshikawa等[11]發現,AG490通過細胞信號抑制因子與JAK2的關聯抑制STAT3的激活,從而使肝癌細胞貼壁生長受到抑制,同時促進細胞凋亡。AG490通過抑制STAT3與DNA結合的活性以抑制STAT3活化與凋亡,從而誘導骨髓瘤細胞U266凋亡,證明AG490有抗腫瘤作用[12]。梁斌等[13]實驗用AG490阻斷JAK2/STAT3信號通路后,人胃癌細胞的增殖受抑,且基質金屬蛋白酶2(MMP-2)、MMP-9表達下降,MMP-2、MMP-9抑制劑TIMP-1、TIMP-2表達上調。
本研究結果顯示,不同濃度的AG490均可使HepG2細胞生長受到抑制,同時誘導細胞發生凋亡,并且呈濃度和時間依賴性。AG490 4 mg/kg組、AG490 6 mg/kg組、AG490 8 mg/kg組P-JAK2、P-STAT3及下游抗凋亡蛋白(Bcl-2、Survivin)表達明顯減弱,而Bax和Caspase-3蛋白表達升高,與對照組比較差異有統計學意義。認為AG490通過與受體酪氨酸激酶競爭結合抑制JAK2磷酸化,使STAT3不能被JAK2磷酸化后再二聚化,有效阻斷STAT3活化,使凋亡相關蛋白表達受到影響,從而抑制腫瘤細胞的生長并促進凋亡。
綜上所述,AG490通過阻斷JAK2/STAT3信號通路的表達抑制腫瘤細胞增殖并促進其凋亡。
[1] Levy DE, Darnell JE. Stats: transcriptional control and biological impact[J]. Nat Rev Mol Cell Biol, 2002,3(9):651-662.
[2] Booz GW, Day JN, Baker KM. Interplay between the cardiac rennin angiotensin system and JAK-STAT signaling: role in cardiac hypertrophy, ischemia/reperfusion dysfunction, and heart failure[J]. J Mol Cell Cardiol, 2002,34(11):1443-1453.
[3] Wung BS , Hsu MC, Wu CC, et al. Resveratrol suppresses IL-6-induced ICAM-1 gene expression in endothelial cells: effects on the inhibition of STAT3 phosphorylation[J]. Life Sci, 2005,78(4):389-397.
[4] Mueller KM, Kornfeld JW, Friedbichler K, et al. Impairment of hepatic growth hormone and glucocorticoid receptor signaling causes steatosis and hepatocellular carcinoma in mice[J]. Hepatology, 2011,54(4):1398-1409.
[5] 龔代鵬,張竹青,張妍,等.p-STAT3、Survivin和Mcl-1蛋白在胃癌組織中的表達及意義[J].中華臨床醫師雜志,2015,9(6): 915-920.
[6] 劉波,王洪林,宋立文.Janus激酶抑制劑AG490對人肝癌SMMC-7721細胞侵襲轉移的影響[J].第三軍醫大學學報,2011,33(18):1960-1963.
[7] Brivanlou AH, Darnell JE Jr. Signal transduction and the control of gene expression[J]. Science, 2002,295(5556):813-818.
[8] Kim MS, Lee WS, Jeong J, et al. Induction of metastatic potential by TrkB via activation of IL6/JAK2/STAT3 and PI3K/AKT signaling in breast cancer[J]. Oncotarget, 2015,6(37):40158-40171.
[9] Li L, Li H, Li M. Curcumin protects against cerebral ischemia-reperfusion injury by activating JAK2/STAT3 signaling pathway in rats[J]. Int J Clin Exp Med, 2015,8(9):14985-14991.
[10] Hsieh FC, Cheng G, Lin J. Evaluation of potential Stat3-regulated genes in human breast cancer[J]. Biochem Biophys Res Commun, 2005,335(2):292-299.
[11] Yoshikawa H, Matsubara K, Qian GS, et al. SOCS-1, a negative regulator of the JAK/STAT pathway, is silenced by methylation in human hepatocellular carcinoma and shows growth-suppression activity[J]. Nat Genet, 2001,28(1):29-35.
[12] Catlett-Falcone R, Landowski TH, Oshiro MM, et al. Constitutive activation of Stat3 signaling confers resistance to apoptosis in human U266 myeloma cells[J]. Immunity, 1999,10(1):105-115.
[13] 梁斌,王杉,葉穎江,等.JAK2激酶抑制劑AG490對胃癌細胞侵襲的影響[J].中華普通外科雜志,2004,19(10):627-630.
河北省醫學科學研究重點課題(20150634)。
胡毅平(E-mail:171955999@qq.com)
10.3969/j.issn.1002-266X.2016.41.011
R735.7
A
1002-266X(2016)41-0038-03
2016-05-30)
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中國生殖健康(2019年3期)2019-02-01 06:12:26
中成藥(2016年8期)2016-05-17 06:08:14
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
