豬流感病毒基因池中的新成員:新甲型H1N1流感病毒基因片段的存在及潛在威脅
2016-10-24 03:41:09高靈茜崔夢一樊曉暉
中國人獸共患病學(xué)報 2016年9期
高靈茜,崔夢一,樊曉暉
?
豬流感病毒基因池中的新成員:新甲型H1N1流感病毒基因片段的存在及潛在威脅
高靈茜,崔夢一,樊曉暉
新甲型H1N1流感病毒(2009 pandemic H1N1 virus,pdm/09)于2009年在人群中暴發(fā)以后,迅速在全球范圍內(nèi)傳播,引起了21世紀(jì)的第一次流感大流行。pdm/09是由人的流感病毒、禽流感和豬流感病毒(swine influenza virus,SIV)經(jīng)過重配后形成的病毒,它的基因片段已經(jīng)進(jìn)入了豬流感病毒當(dāng)中并開始產(chǎn)生新的變異毒株,這些新的變異流感毒株在歐亞大陸、北美大陸及中國南部的各個地區(qū)被不斷報道和發(fā)現(xiàn),這表明豬源性pdm/09在人間流行后可返傳給豬,成為豬流感病毒基因池中的固有組成,獲得與SIV重組形成新的重配病毒的能力,并可能仍然具有感染人類的潛能。因此,必須關(guān)注新型重配病毒的進(jìn)化:包括其在豬群中的生長適應(yīng)、以及適應(yīng)性感染人的進(jìn)化過程。不僅如此,還必須加強(qiáng)對豬群及人群流感病毒的檢測,了解重配病毒在人和豬兩個種群中的進(jìn)化過程。
新甲型H1N1流感病毒;豬流感病毒基因池;基因重配病毒;跨種屬傳播
流感病毒是流行性感冒的病原體,屬于RNA病毒,分為甲乙丙3個型別,最容易發(fā)生變異的是甲型。甲型流感病毒的副鏈RNA有8個節(jié)段,每個節(jié)段為1個基因,這8個基因至少編碼 10種以上蛋白肽: RNA1節(jié)段編碼 PB2蛋白、RNA2編碼 PB1蛋白、RNA3 編碼 PA蛋白、RNA4 編碼 HA蛋白、RNA5編碼 NP蛋白、RNA6 編碼 NA蛋白、RNA7編碼 M1和M2蛋白、RNA8編碼兩個非結(jié)構(gòu)蛋白—— NS1和 NS2[1]。甲型流感病毒依據(jù)其表面血凝素(HA)和神經(jīng)氨酸酶(NA)的差異可分成不同的亞型,迄今為止,已有 17種HA亞型(H1~H17),10種NA亞型(N1~N10)被鑒定[1-3],任何一種HA與任何一種NA結(jié)合即為一種亞型。能夠引起人類感染的亞型包括新甲型H1N1、H2N2、H3N2、H5N1、H5N2、H5N6、H7N2、H7N3、H7N7、H7N9、H9N2和H10N8。歷史上H1N1、H2N2和H3N2引起幾次流感世界大流行,最近出現(xiàn)的一次流感大流行發(fā)生在2009年,由新甲型H1N1流感病毒(2009 pandemic H1N1 virus,pdm/09)引起。
pdm/09在人群中暴發(fā)以后,迅速在全球范圍內(nèi)傳播,引起了21世紀(jì)的第一次流感大流行。pdm/09是由人的流感病毒、禽流感和豬流感病毒(swine influenza virus,SIV)經(jīng)過重配后形成的病毒[4-5],它不僅能感染人,而且在人群流行之后很快回傳至豬群,與SIV進(jìn)一步發(fā)生重配[6-8]。pdm/09的流行和肆虐提示了兩個重要問題:流感病毒在人和豬之間的雙向穿梭感染頻繁的發(fā)生;豬作為中間宿主可以推動人源流感病毒和豬源流感病毒的基因片段發(fā)生重配并可能產(chǎn)生新的大流行株。而pdm/09的基因片段也已經(jīng)進(jìn)入了豬流感病毒當(dāng)中并開始產(chǎn)生新的變異毒株,這些新的變異流感毒株在歐亞大陸和北美大陸各個地區(qū)被不斷報道和發(fā)現(xiàn)[9-16],并有成為大流行株的可能性。本文擬對重配有pdm/09基因片段的SIV的流行情況和潛在危機(jī)做出討論。
1 pdm/09基因組成及流行情況
在兩次流感大范圍流行爆發(fā)之后,pdm/09已經(jīng)變?yōu)榧竟?jié)性流感毒株之一,(數(shù)據(jù)分析來源為WHO于2010年8月10日 的報到([http://www.who.int/mediacentre/news/statements/2010/h1n1_vpc_20100810/en/]).但是pdm/09對于流感病毒的流行學(xué)和生態(tài)學(xué)的影響卻在逐漸加大。pdm/09是由北美豬流感病毒、北美禽流感病毒、人類甲型流感H1N1亞型病毒和常見于歐亞大陸的豬流感病毒重配產(chǎn)生[5],這些病毒已經(jīng)在豬群中流行達(dá)到10年之久,是常見且穩(wěn)定的SIV基因池來源[17]。在2009年pdm/09爆發(fā)之前,世界范圍內(nèi)豬群中存在穩(wěn)定流通的SIV是 以基因重配H1N2亞型、三重重配H3N2亞型(triple reassortant,TR)和歐亞類禽H1N1亞型(European avian-like,ER)為主[18-22],即SIV基因池中三大供給亞型。而三重重配H3N2亞型病毒構(gòu)成了pdm/09除了NA和M基因之外的其他6個基因的主要來源。1979年歐洲一株禽源H1N1病毒進(jìn)入豬群中流通并成為了穩(wěn)定的來源,被命名為歐亞類禽H1N1[23-25],而這株病毒是pdm/09的NA和M基因的主要來源。
可見pdm/09是一種典型的基因重配病毒,一直以來,基因重配學(xué)說是被流感界廣泛接受的一個學(xué)說,該學(xué)說認(rèn)為流感病毒新亞型大流行株是人、禽、豬流感病毒通過基因重配而來,而豬是發(fā)生基因重配的重要場所。豬氣管存在能與禽流感病毒特異性結(jié)合的唾液酸α-2,3Gal受體,又存在能與人流感病毒特異性結(jié)合的唾液酸α-2,6Gal受體,禽流感病毒與人流感病毒均可感染豬,再加上SIV,3個不同來源的病毒易通過基因重配形成新病毒株,由于人群中普遍缺乏對新產(chǎn)生的重配病毒株的免疫力,一旦該病毒在豬中通過適應(yīng)性的變化而具備有效的人傳人能力,將有可能導(dǎo)致流感的大爆發(fā),故豬被認(rèn)為是“基因混合器“,而SIV的流行情況和基因組成更是需要得到關(guān)注和長時間監(jiān)測。
基因重配的病毒也是成為大流行株的重要條件,大量證據(jù)證明,基因重配的流感病毒具有明顯的選擇優(yōu)勢,傳播能力顯著增強(qiáng),從1957年的亞洲流感到1968年香港流感,再到2009年的pdm/09,都是由基因重配的病毒株所引發(fā)。在pdm/09爆發(fā)后的很短時間內(nèi)已經(jīng)可以檢測到其從人傳入豬[6-8,26-28],并在豬中不斷穿梭重配,即與SIV重組形成新的重組病毒,并可能仍然具有感染人類的潛能。因此,必須關(guān)注這一重組病毒株的進(jìn)化包括在豬群中的生長適應(yīng)、以及適應(yīng)感染人的進(jìn)化過程。特別的是,這些基因重配都是以pdm/09內(nèi)部基因的進(jìn)入為主,而表面基因只短暫或偶爾在重配病毒株中發(fā)現(xiàn),但這種趨勢在逐步增大。對帶有pdm/09內(nèi)部基因的重配毒株遺傳分析和系統(tǒng)進(jìn)化研究表明pdm/09的基因片段已經(jīng)開始取代現(xiàn)有歐亞大陸和北美SIV的基因來源(歐亞類禽H1N1、基因重配H1N2和三重重配H3N2病毒),從根本上改變了SIV的病毒池[18-19]。因此必須加強(qiáng)對豬群及人群流感病毒的檢測,了解重組病毒在人和豬兩個種群中的進(jìn)化過程。而這種不同來源的內(nèi)部基因和表面基因的重配組合是否意味著新一輪流感病毒重配的趨勢,還值得我們進(jìn)一步思考。
2 H1N1亞型重配毒株
在2009年4月pdm/09暴發(fā)后的不到一年時間內(nèi),D. Vijaykrishna等人[14]于2010年2月在對香港豬屠宰場的采樣中發(fā)現(xiàn)了一株重配有pdm/09基因片段的H1N1亞型的SIV,通過對同一頭豬進(jìn)行血液,咽拭子和鼻腔拭子3份標(biāo)本的采取,分離得到一株H1N1亞型的SIV—A/swine/Hong Kong/201/2010,遺傳進(jìn)化分析表明該毒株的一個表面基因NA來源于pdm/09,另一個表面基因HA則來源于歐亞類禽H1N1,其他6個內(nèi)部基因的來源是三重重配H3N2,可見pdm/09進(jìn)入到SIV和其他SIV發(fā)生基因重配的速度非常迅速,以先建立穩(wěn)定傳播,再進(jìn)行機(jī)會性重配的模式尋找擴(kuò)大化的感染機(jī)會。這也是最早的關(guān)于pdm/09和其他SIV發(fā)生重配新亞型毒株的報到。很快,在這之后3個月,Huachen Zhu等人[29]也在流感監(jiān)測網(wǎng)絡(luò)中篩查到了重配有pdm/09內(nèi)部基因和歐亞類禽H1N1表面基因的重配病毒A/swine/Guangdong/1361/2010,且毒株來源達(dá)到了10個樣品,說明有pdm/09基因片段的重配病毒有良好的豬群內(nèi)部傳播能力,事實(shí)也證明了這株病毒在豬和豬之間、豬和雪貂之間存在傳播能力并可以在人肺組織進(jìn)行有效復(fù)制,進(jìn)一步提升重配有pdm/09基因片段的SIV感染人的能力和危險性。更重要的是,帶有歐亞類禽H1N1亞型HA基因的病毒在近30年的時間內(nèi)幾乎很少感染到人,所以人群中普遍對于這個基因存在免疫缺陷或免疫空白,推測如果帶有這個基因的重配病毒在獲得pdm/09所賦予的傳播能力進(jìn)而感染到人,人群由于免疫缺陷就可能發(fā)生大規(guī)模的流行,可見豬群中出現(xiàn)帶有歐亞類禽H1N1和pdm/09基因片段的重配病毒,并伴隨大量多重病毒感染的情況不容小覷。在歐洲,Elke Starick[13]等人也于2010年在德國分離到一株重配病毒A/swine/Germany-NRW/R708/10,該病毒的HA和其他6個內(nèi)部基因的來源是pdm/09,NA基因來源于歐洲類禽H1N1,進(jìn)入2011年之后,全球各地關(guān)于重配有pdm/09基因片段的H1N1新亞型病毒的報道更加密集,地點(diǎn)也呈現(xiàn)散發(fā)狀態(tài),泰國、日本等地[11,13]均有出現(xiàn),更加說明pdm/09在豬群中發(fā)生重配的范圍已經(jīng)有在全世界擴(kuò)散甚至大量存在的可能性。
3 H1N2亞型重配病毒
H1N2亞型同樣是SIV中常見的基因型,其來源有人源H1N2,基因重配H1N2和三重重配H3N2病毒,于1994年在英國最先被發(fā)現(xiàn)被開始逐步確立其在豬群中的傳播[30,31],最終成為SIV基因池中固有的基因來源,自從pdm/09爆發(fā)之后,H1N2也在發(fā)生變化。 2010年在德國發(fā)現(xiàn)了可以確立種系穩(wěn)定傳播的H1N2亞型重配病毒,并且從第一株重配病毒A/sw/Papenburg/IDT12653/2010(NA來源于三重重配H3N2亞型,其余7個基因來源于pdm/09)的檢出開始,一直持續(xù)到2012年12月,H1N2亞型重配病毒在德國北部的豬群中的檢出率大大增長,Ana Moreno[12]等人在2010年5月通過對意大利境內(nèi)某養(yǎng)豬場中出現(xiàn)有呼吸系統(tǒng)問題的豬的肺組織勻漿進(jìn)行熒光定量PCR檢測,而非傳統(tǒng)的咽拭子和血清學(xué)鑒定,發(fā)現(xiàn)了呈甲型流感病毒和pdm/09雙重陽性的流感病毒,進(jìn)而篩選出一株H1N2亞型重配毒株A/Sw/It/116114/2010,該病毒除了表面基因NA之外其余的7個基因均來自pdm/09。兩株新型重配病毒的發(fā)現(xiàn)遵循了相似的基因組成,可以認(rèn)為是pdm/09在歐洲豬群中建立穩(wěn)定傳播后,與其他病毒的初步重配行為。幾乎同時(2010年4月),同樣在歐洲的英國,Howard[9]等發(fā)現(xiàn)了命名為A/swine/England/1382/10的H1N2重配毒株,其基因組成變化為除HA和NA之外所有6個內(nèi)部基因均來源于pdm/09,說明pdm/09與其他病毒重配的多樣性在逐步增加。在美洲,情況也不容樂觀,美國發(fā)現(xiàn)的兩株H1N2重配毒株[32](A/swine/Ohio/FAH10-1/2010 和A/swine/Ohio/FAH11-1/2010)的基因組成非常有潛在的威脅性,其HA和NA基因的來源更傾向于類人季節(jié)性H1N2流感病毒,特別是HA基因在系統(tǒng)進(jìn)化重要氨基酸殘基位點(diǎn)96A、133S和183P及抗原決定簇位點(diǎn)91Y、134A、180H、187N和 225G[33]上與人流感病毒保持了高度的一致性。相對保守穩(wěn)定的NP和M基因則來源于pdm/09,導(dǎo)致這兩株病毒獲得了較高的關(guān)注度,直接把重配有pdm/09基因片段的豬流感病毒傳播到人的可能性提升到較高程度。而且有趣的是與其他國家和地區(qū)的重配趨勢不同,美國發(fā)現(xiàn)的至少6株H1N2和3株H3N2重配病毒更傾向于持有來源于pdm/09的M基因[10],而非其他地區(qū)的6個內(nèi)部基因或者2個表面基因,其揭示的不同重配趨勢是否有其他的可能性,值得繼續(xù)關(guān)注。亞洲地區(qū)在中國、日本、韓國均有發(fā)現(xiàn)H1N2重配毒株[15,34-35],其中中國發(fā)現(xiàn)的2株H1N2重配毒株中分別來自廣西及浙江,結(jié)合后面將闡述的H3N2亞型重配毒株的發(fā)現(xiàn)地區(qū),可以說明重配病毒的存在出現(xiàn)了有傾向性的地域問題,而且兩株病毒的發(fā)現(xiàn)地相距千里卻顯示出99.6%~100%的高度遺傳進(jìn)化相似性(PB2、 PB1、 PA、HA、NP、NA、 M和NS基因),提示它們在有共同祖先的前提下,某些陸禽鳥類(鷓鴣、鵪鶉、火雞等)在流感病毒的傳播和基因交流上發(fā)揮著某些不可忽視的重要作用[36-37],對于這些陸禽鳥類的流感病毒受體分布以及不同流感病毒感染后可以作為“放大器”的特點(diǎn),都非常值得進(jìn)一步關(guān)注。
4 H3N2亞型重配病毒
SIV基因池中的第3種供給是H3N2亞型流感病毒,2011年,在美國出現(xiàn)的帶有pdm/09片段的重配毒株H3N2亞型至今已有超過340個人感染[38],即H3N2重配毒株感染人的情況已經(jīng)發(fā)生。2009年8月—2012年9月,筆者所在課題組和香港大學(xué)聯(lián)合的課題組在香港、廣東及廣西得到的37 213份豬呼吸道(鼻粘膜和氣管)拭子標(biāo)本中分離到了387株流感病毒,其中鑒定為H3N2亞型的有94株,廣西地區(qū)的分離率為0.75%。值得注意的是,我們于2010年10月首次在國內(nèi)分離到了重組有pdm/09內(nèi)部基因片段的H3N2 SIV(SW/GX/NS2783/10,SW2783)[8],隨后2011年7—8月間在香港和廣東也分離到類似的重組有pdm/09內(nèi)部基因片段的H3N2,后者在廣東和香港兩地的分離率達(dá)到24.3%[10]。通過對SW2783廣西分離株等毒株的遺傳進(jìn)化分析,發(fā)現(xiàn)廣西病毒株與一些越南H3N2三重重組病毒(triple reassortant,TR,是在2000—2007年間由人季節(jié)性H3N2病毒進(jìn)入豬群后,經(jīng)過三次重配形成的)具有同一宗支來源。更值得關(guān)注的是越南研究同行從一名兩歲越南女孩體內(nèi)分離得到一株H3N2毒株(Vietnam/302/11)(http://thanhniennews.com/health/vietnam-reports-first-human-case-of-h3n2-swine-flu-8302.html),其基因序列和中國廣西分離到的H3N2 SIV高度相關(guān)。這提示在廣西分離得到的重組有pdm/09基因片段的H3N2 SIV可能來自越南,而且已具備了感染人的能力。
如前文提到,禽流感病毒在作為“基因混合器”的豬體內(nèi)長期不斷的增殖復(fù)制,逐漸改變其對受體的特異性,即同時特異性結(jié)合唾液酸α-2,3Gal和α-2,6Gal受體到傾向于結(jié)合唾液酸α-2,6Gal受體(人以此受體為主)而能感染人,提示禽流感病毒在向逐漸適應(yīng)人體的方向進(jìn)化[39]。同樣道理,重組有pdm/09基因片段的H3N2 SIV也可通過在作為潛在“基因混合器”的人體內(nèi)長期不斷的增殖復(fù)制,通過改變其對受體的特異性及其他特性,從而變得更加適應(yīng)于人群。
5 pdm/09基因片段產(chǎn)生的威脅
2009年到2012年間,筆者所在課題組和香港大學(xué)合作分離得到的387份SIV中,有60%的病毒帶有pdm/09基因片段,并形成了至少17種不同的重配基因型。對這些重配毒株進(jìn)行分析后發(fā)現(xiàn):pdm/09的PB2、PB1、PA、NP、M和NS等6個內(nèi)部基因大量存在于重配毒株中(表1)。這一發(fā)現(xiàn)說明來自于人大流行毒株的pdm/09基因片段,可能有利于豬源流感病毒的生存、適應(yīng)和傳播。由于這些新基因型的毒株同時具有豬和人病毒基因,因此在兩種宿主中都有可能復(fù)制、感染,甚至進(jìn)一步傳開。這對流感病毒的進(jìn)化具有深遠(yuǎn)的影響,并有可能對人類構(gòu)成更多樣性的威脅。
表1廣西和香港分離的3株帶有新甲型H1N1流感病毒的H3N2重組型豬流感病毒
Tab.1Three Guangxi or Hong Kong reassortment H3N2 swine influenza viruses carrying genes from the pandemic H1N1/2009
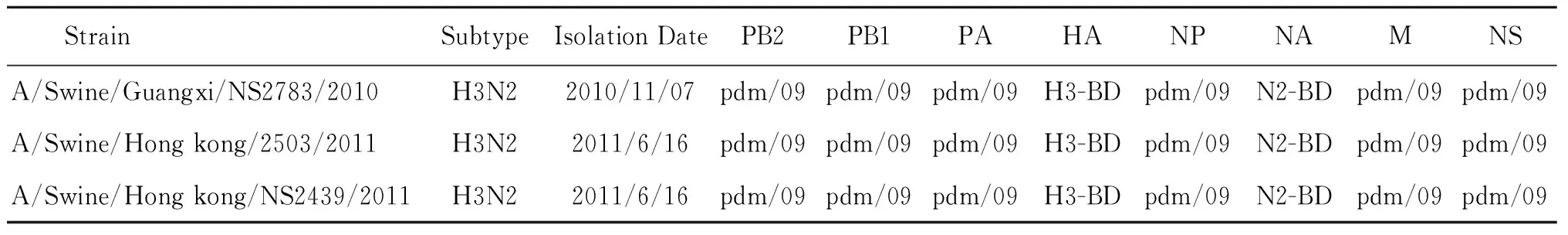
StrainSubtypeIsolationDatePB2PB1PAHANPNAMNSA/Swine/Guangxi/NS2783/2010H3N22010/11/07pdm/09pdm/09pdm/09H3-BDpdm/09N2-BDpdm/09pdm/09A/Swine/Hongkong/2503/2011H3N22011/6/16pdm/09pdm/09pdm/09H3-BDpdm/09N2-BDpdm/09pdm/09A/Swine/Hongkong/NS2439/2011H3N22011/6/16pdm/09pdm/09pdm/09H3-BDpdm/09N2-BDpdm/09pdm/09
注:pdm/09: pandemic H1N1/2009 lineage ,2009年引起世界大流行的新甲型H1N1流感病毒;H3-BD,N2-BD: A/Swine/Binh Duong/03_06/10-like H3N2 lineage,2010年在越南Binh Duong分離到的H3N2三重重組豬流感病毒
考慮到在亞洲(中國東南部)已經(jīng)發(fā)現(xiàn)了H5N1和H9N2亞型流感病毒在豬群中存在流通的情況[40],這些禽源的流感病毒及其基因通過感染豬獲得哺乳動物的適應(yīng)性,那么出現(xiàn)禽源和pdm/09或帶有pdm/09片段的重配毒株發(fā)生進(jìn)一步重配的可能性就時刻存在。我們必須警惕、避免和干預(yù)重組有pdm/09基因片段的H3N2 SIV和禽流感病毒(H5N1、H7N9等能感染哺乳動物和人的病毒)的進(jìn)一步重配,尤其是經(jīng)過長期適應(yīng)后的pdm/09重組病毒或H3N2 SIV,可能以一種更能適應(yīng)人體生長的方式,為禽流感病毒提供有效的哺乳動物適應(yīng)性基因。目前已經(jīng)知道,禽流感病毒可以通過所編碼蛋白的單個或多個氨基酸的突變,增強(qiáng)其對哺乳動物的致病性,例如HA基因的多個堿性氨基酸裂解位點(diǎn)的存在是禽流感病毒最重要的分子毒力基礎(chǔ)。除此之外,HA影響病毒毒力的因素大多與宿主的唾液酸α-2,3Gal和α-2,6Gal受體有關(guān),因?yàn)樗鼈冎饕獩Q定了病毒的宿主范圍和組織親嗜性。對于H5N1禽流感病毒來說,L129V/A134V、N182/KQ222L/G224S、S227N、182/192、G228S這些位點(diǎn)的突變,能增強(qiáng)病毒對α-2,6Gal受體的結(jié)合能力,而對于H1和H3亞型流感病毒,影響受體結(jié)合特性最重要的因素在190至225區(qū)域(H1 numbering)以及226和228區(qū)域(H3 numbering)。對于pdm/09而言,D222G的變化使其受體結(jié)合特性從α-2,6Gal受體轉(zhuǎn)變?yōu)榻Y(jié)合α-2,3Gal和α-2,6Gal兩種受體。那么文中所總結(jié)的重配有pdm/09基因片段的新SIV病毒,其HA是否也遵循上述這些規(guī)律或者有新的變化可循,值得進(jìn)一步探討。
中國具有最為多樣和復(fù)雜的SIV流感生態(tài)學(xué)和基因庫,長期研究只揭示了歐洲類禽和三重重配病毒在豬中存在流通,是SIV基因池的重要供給病毒,直到2009年流感大暴發(fā)之后,才在中國豬群中發(fā)現(xiàn)了帶pdm/09基因片段的重配毒株[29],這些重配毒株的發(fā)現(xiàn)引起了注意,前期對這些重配毒株分析無不證明pdm/09基因片段進(jìn)入豬群并建立了穩(wěn)定大量的傳播(例如帶pdm/09基因片段的重配毒株比帶TR或者EA片段的重配病毒的基因多樣性的水平低,說明其專注在豬群傳播),以往的經(jīng)驗(yàn)說明帶有pdm/09片段的毒株都有成為大流行株的潛力,那么前期研究中發(fā)現(xiàn)的這些重配毒株是否具有傳染人的威脅?或者pdm/09基因片段的出現(xiàn)是否意味著新的流感大暴發(fā)有可能在中國發(fā)生?均是考慮的重點(diǎn),可以通過對這些重配毒株適應(yīng)性感染實(shí)驗(yàn)后的遺傳進(jìn)化分析來探討pdm/09在中國豬群基因池的地位及可能存在的威脅,還可以探討感染后某些位點(diǎn)的變化是否具有標(biāo)記性作用,進(jìn)而為有成為大流行株潛力的毒株提供某些“分子證明”。
[1] Webster RG,Bean WJ,Gorman OT,et al. Evolution and ecology of influenza A viruses[J]. Curr Topics Microbiol Immunol,1992,385(9): 235-235. DOI: 10.1111/j.1751-0813.1992.tb09940.x
[2] Murphy BR. Orthomyxoviruses[J]. Fields Virol,1996: 1397-1445.
[3] Fouchier RAM,Vincent M,Anders W,et al. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from Black-Headed Gulls[J]. J Virol,2005,79(5): 2814-2822. DOI: 10.1128/jvi.79.5.2814-2822.2005
[4] Garten RJ,Davis CT,Russell CA,et al. Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans[J]. Science,2009,325(5937): 197-201. DOI: 10.1126/science.1176225
[5] Smith GJD,Dhanasekaran V,Justin B,et al. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic[J]. Nature,2009,459(7250): 1122-1125. DOI: 10.1038/nature08182
[6] Howden KJ,Brockhoff EJ,Caya FD,et al. An investigation into human pandemic influenza virus (H1N1) 2009 on an Alberta swine farm[J]. Canad Vet J La Revue Veterinaire Canadienne,2009,50(11): 1153-1161.
[7] Ariel P,Javier C,Quiroga MA,et al. Pandemic (H1N1) 2009 outbreak on pig farm,Argentina[J]. Emerg Infect Dis,2010,16(2): 304-307. DOI: 10.3201/eid1602.091230
[8] Weingartl HM,Yohannes B,Tamiko H,et al. Genetic and pathobiologic characterization of pandemic H1N1 2009 influenza viruses from a naturally infected swine herd[J]. J Virol,2009,84(5): 2245-2256. DOI: 10.1128/JVI.02118-09
[9] Howard WA,Essen SC,Strugnell BW,et al. Reassortant pandemic (H1N1) 2009 virus in pigs,United Kingdom[J]. Emerg Infect Dis,2011,17(6): 1049-1052. DOI: 10.3201/eid/1706.101886
[10] Ducatez MF,Ben H,Evelyn SR,et al. Multiple reassortment between pandemic (H1N1) 2009 and endemic influenza viruses in pigs,United States[J]. Emerg Infect Dis,2011,17(9): 1624-1629. DOI: 10.3201/eid1709.110338
[11] Kitikoon P,Sreta D,Ayudhya SNN,et al. Brief report: Molecular characterization of a novel reassorted pandemic H1N1 2009 in Thai pigs[J]. Virus Genes,2011,43(1): 1-5. DOI: 10.1007/s11262-011-0597-5
[12] Moreno A,Di Trani L,F(xiàn)accini S,et al. Novel H1N2 swine influenza reassortant strain in pigs derived from the pandemic H1N1/2009 virus[J]. Vet Microbiol,2011,149(3/4): 472-477. DOI: 10.1016/j.vetmic.2010.12.011
[13] Elke S,Elke L,Sasan F,et al. Reassorted pandemic (H1N1) 2009 influenza A virus discovered from pigs in Germany[J]. J General Virol,2011,92(3): 1184-1188. DOI: 10.1099/vir.0.028662-0
[14] Vijaykrishna D,Poon LLM,Zhu HC,et al. Reassortment of pandemic H1N1/2009 influenza a virus in swine[J]. Science,2010,328(5985): 1529. DOI: 10.1126/science.1189132
[15] Kirisawa R,Ogasawara Y,Yoshitake H,et al. Genomic reassortants of pandemic A (H1N1) 2009 virus and endemic porcine H1 and H3 viruses in swine in Japan[J]. J Vet Med Sci,2014,76: 1457-1470. DOI: 10.1292/jvms.14-0194
[16] Pravina K,Vincent AL,Gauger PC,et al. Pathogenicity and transmission in pigs of the novel A(H3N2)v influenza virus isolated from humans and characterization of swine H3N2 viruses isolated in 2010-2011[J]. J Virol,2012,86(12): 6804-6814. DOI: 10.1128/JVI.00197-12
[17] Zhu H,Webby R,Lam TTY,et al. History of swine influenza viruses in Asia[J]. Curr Topics Microbiol Immunol,2011,370: 57-68. DOI: 10.1007/82_2011_179
[18] Lorusso A,Vincent AL,Gramer MR,et al. Contemporary epidemiology of North American lineage triple reassortant influenza A viruses in pigs[J]. Curr Topics Microbiol Immunol,2011,370: 113-131. DOI: 10.1007/82_2011_196
[19] Webby RJ,Swenson SL,Krauss SL,et al. Evolution of swine H3N2 influenza viruses in the United States[J]. J Virol,2000,74(18): 8243-8251. DOI: 10.1128/jvi.74.18.8243-8251.2000
[20] Brown IH. The epidemiology and evolution of influenza viruses in pigs[J]. Vet Microbiol,2000,74(1): 29-46. DOI: 10.1016/s0378-1135(00)00164-4
[21] Naomi K,Jodie MV,Robert H,et al. A historical perspective of influenza A(H1N2) virus[J]. Emerg Infect Dis,2014,20(1): 6-12. DOI: 10.3201/eid2001.121848
[22] Karasin A,Olsen CG. Genetic characterization of an H1N2 influenza virus isolated from a pig in Indiana[J]. J Clin Microbiol,2000,38(6): 2453-2456.
[23] Brown IH. History and epidemiology of swine influenza in Europe. Swine Influenza[M]. Berlin: Springer. 2013:133-146. DOI: 10.1007/82_2011_194
[24] Castrucci MR,Donatelli I,Sidoli L,et al. Genetic reassortment between avian and human influenza A viruses in Italian pigs[J]. Virology,1993,193(1): 503-506. DOI: 10.1006/viro.1993.1155
[25] Ludwig S,Stitz L,Planz O,et al. European swine virus as a possible source for the next influenza pandemic?[J]. Virology,1995,212(2): 555-561. DOI: 10.1006/viro.1995.1513
[26] Welsh MD,Baird PM,Guelbenzu-Gonzalo MP,et al. Initial incursion of pandemic (H1N1) 2009 influenza A virus into European pigs[J]. Vet Rec,2010,166(21): 642-645. DOI: 10.1136/vr.4851
[27] Deng YM,Iannello P,Smith I,et al. Transmission of influenza A(H1N1) 2009 pandemic viruses in Australian swine[J]. Influenza & Other Respir Viruses,2012,6(3): e42-e47. DOI: 10.1111/j.1750-2659.2012.00337.x
[28] Min-Suk S,Jun Han L,Pascua PNQ,et al. Evidence of human-to-swine transmission of the pandemic (H1N1) 2009 influenza virus in South Korea[J]. J Clin Microbiol,2010,48(9): 3204-3211. DOI: 10.1128/jcm.00053-10
[29] Huachen Z,Boping Z,Xiaohui F,et al. Novel reassortment of Eurasian avian-like and pandemic/2009 influenza viruses in swine: infectious potential for human[J]. J Virol,2011,85(20): 10432-10439. DOI: 10.1128/jvi.05352-11
[30] Brown IH,Chakraverty P,Harris PA,et al. Disease outbreaks in pigs in Great Britain due to an influenza A virus of H1N2 subtype[J]. Vet Rec,1995,136(13): 328-329. DOI: 10.1136/vr.136.13.328
[31] Brown IH,Harris PA,Mccauley JW,et al. Multiple genetic reassortment of avian and human influenza A viruses in European pigs,resulting in the emergence of an H1N2 virus of novel genotype[J]. J General Virol,1998,79 (pt 12)(3): 2947-2955. DOI: 10.1099/0022-1317-79-12-2947
[32] Ali A,Khatri M,Wang L,et al. Identification of swine H1N2/pandemic H1N1 reassortant influenza virus in pigs,United States[J]. Vet Microbiol,2012,158(1/2): 6068. DOI: 10.1016/j.vetmic.2012.02.014
[33] Alessio L,Vincent AL,Harland ML,et al. Genetic and antigenic characterization of H1 influenza viruses from United States swine from 2008[J]. J General Virol,2011,92(4): 919-930. DOI: 10.1099/vir.0.027557-0
[34] Chuanling Q,Liping L,Huanliang Y,et al. Novel triple reassortant H1N2 influenza viruses bearing six internal genes of the pandemic 2009/H1N1 influenza virus were detected in pigs in China[J]. J Clin Virol,2014,61(4): 529-534. DOI: 10.1016/j.jcv.2014.10.014
[35] Han JY,Park SJ,Kim HK,et al. Identification of reassortant pandemic H1N1 influenza virus in Korean pigs[J]. J Microbiol Biotechnol,2012,22(5): 699-707. DOI: 10.4014/jmb.1106.05062
[36] Yang L,Zhang ZF,F(xiàn)an XH,et al. Distribution characteristics of avian influenza virus and human influenza virus sialic acid receptors in chukar[J]. Chin J Zoonoses,2014,30(6): 545-550. DOI: 10.3969/cjz.j.issn.1002-2694.2014.06.001 (in Chinese)
楊利,張?jiān)龇澹畷詴?等. 鷓鴣禽流感病毒和人流感病毒唾液酸受體分布特征[J]. 中國人獸共患病學(xué)報,2014,30(6): 545-550.
[37] Zhang ZF,Luo DZ,F(xiàn)an XH,et al. Distribution characteristics and roles of sialic acid receptor of quail in the ecosystem of influenza A virus[J]. Guangxi Med J,2010,32(11): 1313-1317. DOI: 10.3969/j.issn.0253-4304.2010.11.001 (in Chinese)
張?jiān)龇澹_殿中,樊曉暉. 鵪鶉流感病毒SA受體的分布特點(diǎn)及其在流感病毒生態(tài)系統(tǒng)中的作用[J]. 廣西醫(yī)學(xué),2010,32(11): 1313-1317.
[38] Jhung MA,Epperson S,Biggerstaff M,et al. Outbreak of variant influenza A(H3N2) virus in the United States[J]. Clin Infect Dis,2013,57(12): 1703-1712. DOI: 10.1093/cid/cit649
[39] Makarova NV,Ozaki HH,Webster RG,et al. Replication and transmission of influenza viruses in Japanese quail[J]. Virology,2003,310(1): 8-15. DOI: 10.1016/s0042-6822(03)00094-1[40] Peiris JS,Guan Y,Markwell D,et al. Cocirculation of avian H9N2 and contemporary "human" H3N2 influenza A viruses in pigs in southeastern China: potential for genetic reassortment?[J]. J Virol,2001,75(20): 9679-9686. DOI: 10.1128/jvi.75.20.9679-9686.2001
The new member of the swine influenza virus gene pool: the existence of the 2009 pandemic H1N1 virus gene fragments and potential threats
GAO Ling-xi,CUI Meng-yi,F(xiàn)AN Xiao-hui
(DepartmentofMicrobiology,SchoolofPreclinicalMedicine,GuangxiMedicalUniversity,Nanning530021,China)
The pandemic 2009 H1N1 influenza virus (pdm/09) has caused the 2009 pandemic and the first influenza pandemic of the 21st century. It was a swine-origin virus that was generated via multiple reassortments of the swine,human and avian influenza viruses. Pdm/09 can not only infect human but also transmit back into swine and further reassortments with the enzootic swine influenza virus (SIV). New gene reassortment viruses with gene segment of pdm/09 and other SIVs have repeated occurred in Eurasia,North America and southern China,which indicate swine-origin pdm/09 can transmit back into swine after human epidemic and become one of composition in the swine influenza virus gene pool. So the gene reassortment viruses may still have the potential to infect humans. Therefore,we must focus on the evolution of the new gene reassortment viruses,including swine epidemic and adaptable infect humans. Not only that,it need to strengthen the detection of influenza viruses in swine and human,to understand the evolution and cross-species infection of new reassortment viruses in both human and swine.
2009 pandemic H1N1 virus; swine influenza virus gene pool; gene ressortment viruses; cross-species infection
Fan Xiao-hui,Email: fanxiaohui63@163.com
10.3969/j.issn.1002-2694.2016.09.013
樊曉暉,Email:fanxiaohui63@163.com
廣西醫(yī)科大學(xué)微生物學(xué)教研室,南寧530021
R373.1
A
1002-2694(2016)09-0832-06
2015-12-21;
2016-04-19
廣西科學(xué)研究與技術(shù)開發(fā)計(jì)劃(No.桂科合1347004-27),廣西研究生教育創(chuàng)新計(jì)劃(No.YCBZ2014027),廣西醫(yī)科大學(xué)青年科學(xué)基金項(xiàng)目(No. GXMUYSF201525)以及“廣西特聘專家”專項(xiàng)經(jīng)費(fèi)聯(lián)合資助
Supported by the Guangxi Scientific Research and Technological Development Fund (No. 1347004-27),the Guangxi Graduate Education Innovation Fund (No. YCBZ2014027),the Guangxi Medical Youth Science Fund (No. GXMUYSF201525) and
"Guangxi Distinguished experts" Fund
