念珠菌生物被膜和抗真菌藥物選擇
2016-09-26 09:28:30李建國
中國感染與化療雜志 2016年2期
李建國
·綜述·
念珠菌生物被膜和抗真菌藥物選擇
李建國
念珠菌; 生物被膜; 抗真菌藥物; 耐藥機制
由于各種高效廣譜抗菌藥物的廣泛使用,病原微生物產生了多種耐藥機制以保持物種的延續,而形成生物被膜就是一種重要的耐藥機制[1-2]。生物被膜是指大量微生物黏附于某些物體的表面形成的一種膜樣聚集,具有復雜的三維結構和功能[2]。隨著現代醫學的發展,人體植入物和移植物的應用越來越廣泛,病原微生物可附著于植入性醫療器材(如各種人工假體、支架、血管內插管、各種類型導管等)的表面形成生物被膜,逃避宿主的免疫防御并抵制抗菌藥物的作用,目前認為人體80%微生物感染與病原微生物的生物被膜相關[3]。
念珠菌屬是人體內最大的正常真菌菌群,廣泛定植于機體與外界相通的各個器官,包括口咽部、鼻咽部、胃腸道、前尿道和陰道等部位[4]。念珠菌屬是造成人類侵襲性真菌感染最常見的條件致病真菌,73.4%侵襲性真菌感染由念珠菌引起[5]。可導致感染的念珠菌超過17種,其中90%以上的感染由白念珠菌、光滑念珠菌、近平滑念珠菌、熱帶念珠菌和克柔念珠菌引起,而白念珠菌就造成超過60%的感染[6]。念珠菌所致感染類型廣泛,從并不危及生命的皮膚黏膜感染,到多臟器的侵襲性感染[7]。念珠菌極易在各種植入性器材表面形成生物被膜,在人體免疫功能低下時造成機體感染,從而導致念珠菌血流感染及其他各種危及生命的侵襲性念珠菌病,病死率高達30%[8]。
生物被膜相關性感染會影響患者預后,臨床研究表明由產生生物被膜念珠菌造成的念珠菌血流感染其致死率是由不產生生物被膜念珠菌造成的血流感染的1.77倍[9],生物被膜產生也是念珠菌血流感染患者死亡的獨立危險因素[10]。動物實驗也表明,從生物被膜擴散的游離念珠菌較非生物被膜游離念珠菌具有更強的毒力,從而其感染可造成更差的結局[11]。生物被膜相關性感染很難控制,因為其具有更強的耐藥性,普通抗真菌藥物治療很難完全清除生物被膜[1]。目前臨床對念珠菌生物被膜尚無理想對策,一旦發生,只能取出植入器材,而這同樣嚴重影響患者預后[1]。如何防治和克服生物被膜耐藥問題,已成為臨床上極具挑戰性的課題。
1 念珠菌生物被膜的形成與耐藥機制
生物被膜是由微生物自身分泌的胞外多聚基質包裹的微生物聚集體,其中90%以上的干物質為胞外聚合物[2]。胞外多聚基質主要由胞外多糖、蛋白、核酸和脂類物質組成,形成穩定三維結構支架,介導生物被膜附著于物體表面[2],為微生物生長提供一個相對穩定、安全的微環境[3]。一般生物被膜形成有以下步驟:①微生物附著于物體表面;②形成離散菌群結構;③分泌胞外多聚基質,形成成熟三維結構;④生物被膜微生物聚合體擴散形成子代生物被膜[12]。體外研究表明,白念珠菌生物被膜形成要經過3個階段:①早期(約0~11 h):大部分白念珠菌細胞以芽生孢子的形式黏附于生物材料表面并形成明顯菌落;②中期(約12~30 h):細胞分泌胞外多聚基質覆蓋在菌落表面,形成由孢子、胚芽管和新菌絲樣細胞組成的雙層結構;③成熟期(約38~72 h):細胞外多聚基質包裹著芽生孢子和菌絲細胞組成網狀結構,厚度在2~450 μm[12]。念珠菌生物被膜在動物體內形成也有類似步驟,只是厚度一般都超過100 μm[13]。
生物被膜的形成受附著基底層、營養、唾液或血液、可使用氧濃度、胞外多聚基質和念珠菌種屬等多種因素的影響[14]。很多材料如有機硅支架、有機玻璃、塑料和玻璃表面都可以形成生物被膜,這些材料的理化性質影響白念珠菌附著和生物被膜的形成。以葡萄糖、果糖或乳糖為碳源比蔗糖和麥芽糖更容易形成白念珠菌生物被膜。而一般植入物直接與身體體液,如唾液或是血液接觸后,會直接裹上一層薄膜,可能影響生物被膜形成。盡管念珠菌在有氧、無氧條件下均能生長,念珠菌生物被膜形成是否也如此目前尚無定論。胞外多聚基質的形成與營養流動相關,流動營養比靜止營養更容易增加胞外多聚基質的合成。不同念珠菌種屬形成的生物被膜也有差別,野生菌比實驗室菌形成的生物被膜更“健康光滑”。生物被膜具有高度耐藥性的特異表型,其耐藥濃度可以比單個個體微生物高1 000倍,其耐藥機制主要有:①生物被膜內細胞高密度導致細胞生長速率降低,代謝活性降低,對抗真菌藥物不敏感;②生物被膜胞外多聚基質的保護作用,阻礙藥物進入細胞發揮作用;③生物被膜細胞大量表達外排泵蛋白等與耐藥相關的蛋白;④一些抗真菌藥物靶點細胞膜麥角固醇表達降低;⑤滯留菌(persister cells)形成核心亞群抵抗真菌藥物;⑥生物被膜壓力應激反應。
1.1 細胞密度、群體感應和生長速率
生物被膜內細胞密度高也可能是其耐藥機制之一。研究表明高細胞密度比低細胞密度具有更高的耐藥性,而且生物被膜重新分散后,高密度細胞也比低密度細胞耐藥性更高[15]。白念珠菌有兩個功能相反的群體感應分子:酪醇和法尼醇,體外法尼醇可以和多種抗真菌藥物如氟康唑、兩性霉素B和米卡芬凈協同作用抗念珠菌生物被膜[16]。有些研究者還認為生物被膜內細胞密度太高,得不到充分的營養而導致的細胞生長速率低下可能引起其耐藥,這一觀點已經在一些細菌生物被膜中被證實[1]。不過有研究表明生物被膜內和游離培養的細胞生長速率并沒有差異[12],而且,不同生長速率的生物被膜細胞對兩性霉素B的耐藥性比較一致,表明念珠菌生物被膜的耐藥性可能與細胞生長速率無關[17]。
1.2 胞外多聚基質的作用
念珠菌形成的成熟生物被膜中胞外基質厚達450 μm,一些研究者認為,這些胞外基質可以通過形成物理屏障阻止抗菌藥物進入細胞內而產生耐藥作用[3]。有研究表明除去念珠菌生物被膜外基質后再用兩性霉素B處理,膜內真菌細胞的生存率降低20%,而且結果表明熱帶念珠菌生物被膜胞外多聚基質厚度與其耐藥程度相關[18]。生物被膜基質可以隔離放射性標記的氟康唑,這種耐藥機制可能與基質中的β-1,3-葡聚糖有關[19]。而且生物被膜可以與兩性霉素B、氟胞嘧啶和棘白菌素類藥物相互作用從而表現出耐藥性[20-21]。生物被膜胞外基質中的DNA也會影響其耐藥性,白念珠菌生物被膜用DNA酶處理后可以提高其對棘白菌素類和多烯類藥物的敏感性,但是不影響對氟康唑的敏感性[22-23]。
1.3 外排泵蛋白高表達
外排泵相關蛋白高表達可以將藥物排出細胞外,降低細胞內的抗菌藥物濃度。在無抗菌藥物存在時,ATP結合盒轉運蛋白CDR1和CDR2與易化擴散超家族轉運蛋白MDR1表達水平都很低[24],但在形成生物被膜時其表達明顯升高,可能是白念珠菌對唑類藥物耐藥的機制之一[25]。單基因或雙基因敲除突變株(CDRl,CDR2,MDR1, CDRl/ CDR2,MDR1/CDRl)在游離狀態下對氟康唑的敏感性增高,但形成生物被膜之后仍具有較高的耐藥性[25]。通過多基因敲除菌株實驗表明,在生物被膜形成早期(6 h),與無突變菌株相比,雙敲除或三敲除菌株形成的生物被膜對氟康唑敏感性明顯增加,但是在中后期(12、48 h),野生株和基因敲除株形成的生物被膜對氟康唑的敏感性沒有差異,表明生物被膜形成不同階段外排泵基因的表達有所差異,外排泵主要在生物被膜形成早期起作用[26]。
1.4 甾醇合成的影響
唑類藥物抗真菌的作用機制是抑制真菌細胞膜成分麥角固醇的生物合成,而兩性霉素B也是通過與細胞膜上的麥角固醇結合發揮作用,因此,甾醇合成變化也是念珠菌耐藥機制之一。研究表明,生物被膜在形成過程中細胞膜麥角固醇含量顯著降低,尤其是在中后期,其含量比早期下降了近50%[26]。因此成熟生物被膜對麥角固醇靶向藥物具有較好的耐藥性。還有研究表明生物被膜形成不同階段甾醇合成相關基因轉錄水平明顯變化[27]。ERG11編碼羊毛甾醇14α去甲基酶是唑類藥物的作用靶點,ERG25編碼C4甲基甾醇氧化酶,參與甾醇中間體合成,芯片分析表明ERG25 和ERG11在念珠菌體外生物被膜形成過程中轉錄水平升高,ERG25轉錄水平在體內生物被膜形成過程中也顯著升高[28]。
1.5 滯留菌
滯留菌是菌群中所占比例不到0.1%的一類表型異化的亞群,具有與種群內普通菌、抗性突變菌所不同的特征,可通過“休眠-生長-增殖”應對逆境的脅迫,維持自身生存和菌體結構穩定[29]。其生長速度不如正常生物被膜細胞,但能夠在高濃度抗菌藥物下存活并且形成菌落[30],藥物撤除之后,這些滯留菌可形成新的生物被膜[31]。這些細胞和野生株在甾醇和β-1,6-葡聚糖相關基因轉錄上有差異[32]。研究顯示,在不同種念珠菌形成的生物被膜中,有的可發現滯留菌,但有些菌種未發現滯留菌的存在,提示滯留菌并不是念珠菌生物被膜耐藥的唯一機制[33]。
1.6 生物被膜壓力應激
生物被膜形成過程中一些壓力應激激活可能會引起耐藥性。細胞膜壓力激活相關信號通路相關蛋白MKC1基因突變會影響菌絲形成、生物被膜形成并且增加其對氟康唑的敏感性[34]。鈣調磷酸酶參與調控真菌生理結構,UPPULURI等[35]研究發現鈣調磷酸酶增加白念珠菌生物被膜對氟康唑的敏感性。熱休克蛋白HSP90可以穩定MKC1蛋白和鈣調磷酸酶因而可以維持生物被膜的耐藥性,而且,抑制HSP90還可以降低基質葡聚糖合成,抑制HSP90功能可以明顯提高生物被膜對各種抗真菌藥物的敏感性[36]。
2 抗念珠菌生物被膜的藥物或材料
抗念珠菌生物被膜藥物主要也是目前臨床上使用的抗真菌藥物,主要作用于2個靶點:細胞壁或質膜(如棘白菌素類、唑類和多烯類)和核酸或蛋白合成(如氟胞嘧啶)。不過如前所述,念珠菌生物被膜具有較高的耐唑類藥物能力,唑類藥物在抗生物被膜治療中不建議使用,見表1。
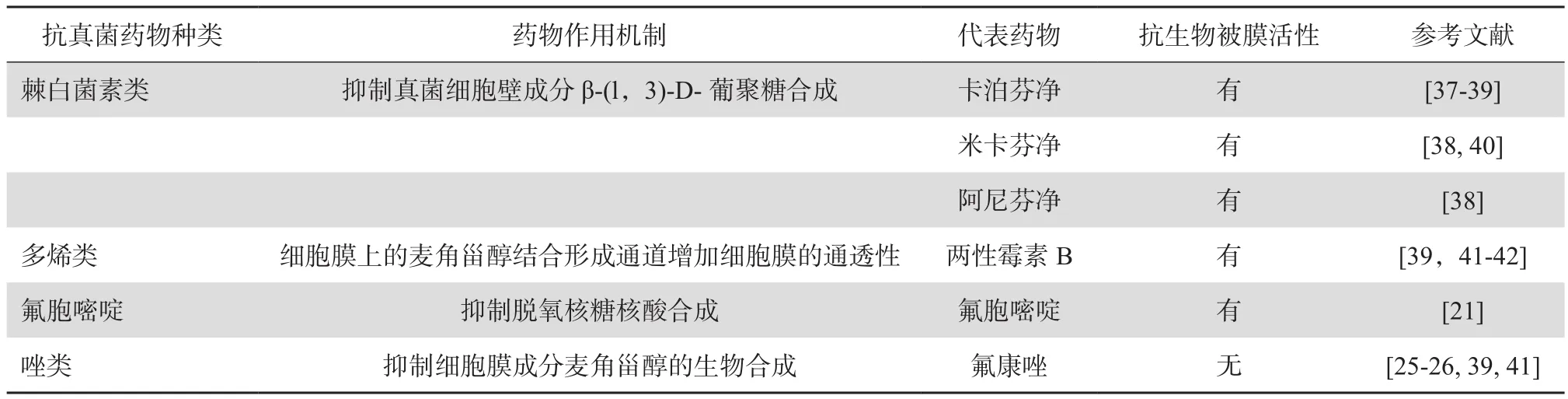
表1 常用抗真菌藥物的抗念珠菌生物被膜活性
2.1 棘白菌素類
真菌細胞壁成分包括幾丁質、β-(l,3)-D-葡聚糖和甘露糖蛋白。棘白菌素類抗真菌藥物可以抑制β-(1,3)-D-葡聚糖合成酶活性從而抑制葡聚糖的合成,導致細胞壁缺損,最終導致真菌細胞的溶解[43]。目前臨床使用的此類藥物主要有卡泊芬凈、米卡芬凈和阿尼芬凈等。棘白菌素類藥物在體外具有較好的抗念珠菌生物被膜活性,卡泊芬凈可以抑制細胞黏附于材料表面、改變被膜內細胞的形態和代謝狀態,生理血液濃度的米卡芬凈體外具有抗念珠菌生物被膜活性[40]。它們還可以通過影響葡聚糖影響胞外多聚基質降低生物被膜的耐藥性而發揮作用[19]。中心靜脈導管念珠菌病小鼠模型中的研究表明卡泊芬凈可以抑制白念珠菌被膜形成[37],KUCHARIKOVA等[38]使用大鼠支架模型研究也表明所有的棘白菌素類藥物都可以有效抑制白念珠菌被膜形成,說明棘白菌素類藥物在臨床中對念珠菌生物被膜可能會有比較好的療效。
2.2 多烯類
多烯類藥物抗菌機制是通過與細胞膜上的麥角甾醇結合形成通道增加細胞膜的通透性,使胞內一些小分子外泄從而造成細胞死亡。這類藥物的代表有兩性霉素和制霉菌素。多個體外研究表明高濃度兩性霉素B脂質體形式具有較好的抗念珠菌生物被膜活性[39],不過對近平滑念珠菌生物被膜作用較差[42]。體內實驗也表明兩性霉素B脂質體可以清除兔體內支架上形成的白念珠菌生物被膜[43]。
2.3 氟胞嘧啶
氟胞嘧啶是作用于核酸合成的抗真菌藥物,其作用機制是藥物通過細胞的滲透酶系統進入細胞內轉化為氟尿嘧啶,競爭尿嘧啶進入真菌的核糖核酸中,從而阻斷真菌RNA的合成,最終抑制蛋白質的合成[1]。氟胞嘧啶單獨使用容易產生耐藥性,所以一般與其他抗真菌藥物聯用。
2.4 其他潛在的生物被膜治療藥物
2.4.1 抗菌肽 抗菌肽是一類具有廣譜抗菌活性的堿性多肽物質,由15~30個氨基酸組成,可以和微生物表面帶負電荷的結構分子結合從而抑制微生物生長,抗菌肽還可以通過修飾一些氨基酸基團增強其抗菌效果和穩定性[3]。修飾后的特定靶向性的抗菌肽STAMPs可以有效抑制變異鏈球菌生物被膜的形成[44]。
2.4.2 一些天然產物 很多中草藥活性成分表現出一定的抗生物被膜活性,體外研究表明黃芩素、甘草查爾酮A、苦參堿、黃根醇、麝香草酚、提琴形凹唇姜和灰羅勒中的揮發油、茶樹油、桉葉素等都有較好的抗白念珠菌生物被膜的活性,可以抑制生物被膜的形成,有些對成熟的生物被膜也有很強的抑制作用[43]。
2.4.3 協同作用藥物 有些藥物具有協同作用,可以增強一些臨床一線抗真菌藥物的抗真菌活性。最新研究表明枸櫞酸托瑞米芬可以增強卡泊芬凈和兩性霉素B體內外抗生物被膜活性[45],這也為我們提供了生物被膜治療新思路,也許我們可以通過一些具有協同作用的化合物來增強現有抗真菌藥物的抗生物被膜活性。
2.5 具有抗生物被膜活性的生物材料
生物被膜的產生主要是微生物附著于植入的生物器材,因此如何從源頭上防治生物被膜的產生也是防治中的重要一環。目前很多研究者致力于研究開發具有一定抗生物被膜形成的生物材料,比如納米金屬顆粒和離子包被、抗菌藥物固定嵌入到生物材料中、對生物材料聚合物進行一些特殊修飾等,這些措施都已經在臨床上取得了一定的成效[46-47]。
3 問題和展望
念珠菌生物被膜是當前臨床感染疾病治療的一大難題。盡管很多體外研究表明一些化合物具有體外抗真菌或抗生物被膜活性,但只是為藥物開發提供了一些研究思路,距離進入臨床應用還十分遙遠。目前對念珠菌生物被膜感染相關疾病要從防和治兩方面著手,在早期以預防為主,包括使用各種具有抗真菌活性的植入器材,盡量避免生物被膜的發生。治療方面,盡量優先選用針對念珠菌生物被膜有效的抗真菌藥物,最大程度發揮藥物作用,達到最佳的治療效果。
致謝
美捷登生物科技有限公司的廖慶嬌博士為本文提供了醫學編輯的輔助工作,默沙東(中國)投資有限公司為這些輔助工作提供了資助。
[1]TAFF HT, MITCHELL KF, EDWARD JA, et al. Mechanisms of Candida biofilm drug resistance[J]. Future Microbiol, 2013,8(10):1325-1337.
[2]FLEMMING HC, WINGENDER J. The biofilm matrix[J]. Nat Rev Microbiol, 2010, 8(9):623-633.
[3]SUN F, QU F, LING Y, et al. Biofilm-associated infections:antibiotic resistance and novel therapeutic strategies[J]. Future Microbiol, 2013, 8(7):877-886.
[4]張嬰元,汪復. 念珠菌病診斷與治療:專家共識[J]. 中國感染與化療雜志, 2011, 11(2):81-95.
[5]AZIE N, NEOFYTOS D, PFALLER M, et al. The PATH (Prospective Antifungal Therapy) Alliance(R) registry and invasive fungal infections: update 2012[J]. Diagn Microbiol Infect Dis, 2012, 73(4):293-300.
[6]PFALLER MA, DIEKEMA DJ, RINALDI MG, et al. Results from the ARTEMIS DISK Global Antifungal Surveillance Study: a 6.5-year analysis of susceptibilities of Candida and other yeast species to fluconazole and voriconazole by standardized disk diffusion testing[J]. J Clin Microbiol, 2005,43(12):5848-5859.
[7]PAPPAS PG, KAUFFMAN CA, ANDES D, et al. Clinical practice guidelines for the management of candidiasis: 2009 update by the Infectious Diseases Society of America[J]. Clin Infect Dis, 2009, 48(5):503-535.
[8]SARDI JC, SCORZONI L, BERNARDI T, et al.Candida species: current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options[J]. J Med Microbiol, 2013, 62(Pt 1):10-24.
[9]TUMBARELLO M, FIORI B, TRECARICHI EM, et al. Risk factors and outcomes of candidemia caused by biofilm-forming isolates in a tertiary care hospital[J]. PLoS One, 2012, 7(3):e33705.
[10]TUMBARELLO M, POSTERARO B, TRECARICHI EM,et al. Biofilm production by Candida species and inadequate antifungal therapy as predictors of mortality for patients with candidemia[J]. J Clini Microbiol, 2007, 45(6):1843-1850.
[11]UPPULURI P, CHATURVEDI AK, SRINIVASAN A, et al. Dispersion as an important step in the Candida albicans biofilm developmental cycle[J]. PLoS Pathogens, 2010, 6(3):e1000828.
[12]CHANDRA J, KUHN DM, MUKHERJEE PK, et al. Biofilm formation by the fungal pathogen Candida albicans:development, architecture, and drug resistance[J]. J Bacteriol,2001, 183(18):5385-5394.
[13]ANDES D, NETT J, OSCHEL P, et al. Development and characterization of an in vivo central venous catheter Candida albicans biofilm model[J]. Infect Immun, 2004, 72(10):6023-6031.
[14]SENEVIRATNE CJ, JIN L, SAMARANAYAKE LP. Biofilm lifestyle of Candida: a mini review[J]. Oral Dis, 2008, 14(7):582-590.
[15]PERUMAL P, MEKALA S, CHAFFIN WL. Role for cell density in antifungal drug resistance in Candida albicans biofilms[J]. Antimicrob Agents Chemother, 2007, 51(7):2454-2463.
[16]KATRAGKOU A, MCCARTHY M, ALEXANDER EL,et al. In vitro interactions between farnesol and fluconazole,amphotericin B or micafungin against Candida albicans biofilms[J]. J Antimicrob Chemother, 2015, 70(2):470-478.
[17]BAILLIE GS, DOUGLAS LJ. Effect of growth rate on resistance of Candida albicans biofilms to antifungal agents[J]. Antimicrob Agents Chemother, 1998, 42(8):1900-1905.
[18]AL-FATTANI MA, DOUGLAS LJ. Biofilm matrix ofCandida albicans and Candida tropicalis: chemical composition and role in drug resistance[J]. J Med Microbiol, 2006, 55(Pt 8):999-1008.
[19]NETT J, LINCOLN L, MARCHILLO K, et al. Putative role of beta-1,3 glucans in Candida albicans biofilm resistance[J]. Antimicrob Agents Chemother, 2007, 51(2):510-520.
[20]VEDIYAPPAN G, ROSSIGNOL T, d'ENFERT C. Interaction of Candida albicans biofilms with antifungals: transcriptional response and binding of antifungals to beta-glucans[J]. Antimicrob Agents Chemother, 2010, 54(5):2096-2111.
[21]NETT JE, CRAWFORD K, MARCHILLO K, et al. Role of Fks1p and matrix glucan in Candida albicans biofilm resistance to an echinocandin, pyrimidine, and polyene[J]. Antimicrob Agents Chemother, 2010, 54(8):3505-3508.
[22]MARTINS M, UPPULURI P, THOMAS DP, et al. Presence of extracellular DNA in the Candida albicans biofilm matrix and its contribution to biofilms[J]. Mycopathologia, 2010, 169(5):323-331.
[23]MARTINS M, HENRIQUES M, LOPEZ-RIBOT JL, et al. Addition of DNase improves the in vitro activity of antifungal drugs against Candida albicans biofilms[J]. Mycoses, 2012, 55 (1):80-85.
[24]SCHUBERT S, BARKER KS, ZNAIDI S, et al. Regulation of efflux pump expression and drug resistance by the transcription factors Mrr1, Upc2, and Cap1 in Candida albicans[J]. Antimicrob Agents Chemother, 2011, 55(5):2212-2223.
[25]RAMAGE G, BACHMANN S, PATTERSON TF, et al. Investigation of multidrug efflux pumps in relation to fluconazole resistance in Candida albicans biofilms[J]. J Antimicrob Chemother, 2002, 49(6):973-980.
[26]MUKHERJEE PK, CHANDRA J, KUHN DM, et al. Mechanism of fluconazole resistance in Candida albicans biofilms: phase-specific role of efflux pumps and membrane sterols[J]. Infect Immun, 2003, 71(8):4333-4340.
[27]FINKEL JS, MITCHELL AP. Genetic control ofCandida albicans biofilm development[J]. Nat Revi Microbiol, 2011, 9 (2):109-118.
[28]NETT JE, LEPAK AJ, MARCHILLO K, et al. Time course global gene expression analysis of an in vivo Candida biofilm[J]. J Infect Dis, 2009, 200(2):307-313.
[29]龍冬艷, 胡少平, 陳新才, 等. 持留菌及其對微生物膜的耐受性影響研究進展[J]. 應用生態學報, 2010, 21(10):2707-2714.
[30]KEREN I, KALDALU N, SPOERING A, et al. Persister cells and tolerance to antimicrobials[J]. FEMS Microbiol Letters,2004, 230(1):13-18.
[31]LAFLEUR MD, KUMAMOTO CA, LEWIS K.Candida albicans biofilms produce antifungal-tolerant persister cells[J]. Antimicrob Agents Chemother, 2006, 50(11):3839-3846.
[32]KHOT PD, SUCI PA, MILLER RL, et al. A small subpopulation of blastospores in Candida albicans biofilms exhibit resistance to amphotericin B associated with differential regulation of ergosterol and beta-1,6-glucan pathway genes[J]. Antimicrob Agents Chemother, 2006, 50(11):3708-3716.
[33]AL-DHAHERI RS, DOUGLAS LJ. Absence of amphotericin B-tolerant persister cells in biofilms of some Candida species[J]. Antimicrob Agents Chemother, 2008, 52(5):1884-1887.
[34]KUMAMOTO CA. A contact-activated kinase signalsCandida albicans invasive growth and biofilm development[J]. Proc Nati Acad Sci USA, 2005, 102(15):5576-5581.
[35]UPPULURI P, NETT J, HEITMAN J, et al. Synergistic effect of calcineurin inhibitors and fluconazole against Candida albicans biofilms[J]. Antimicrob Agents Chemother, 2008, 52 (3):1127-1132.
[36]ROBBINS N, UPPULURI P, NETT J, et al. Hsp90 governs dispersion and drug resistance of fungal biofilms[J]. PLoS Pathogens, 2011, 7(9):e1002257.
[37]LAZZELL AL, CHATURVEDI AK, PIERCE CG, et al. Treatment and prevention of Candida albicans biofilms with caspofungin in a novel central venous catheter murine model of candidiasis[J]. J Antimicrob Chemother, 2009, 64(3):567-570.
[38]KUCHARIKOVA S, SHARMA N, SPRIET I, et al. Activities of systemically administered echinocandins against in vivo mature Candida albicans biofilms developed in a rat subcutaneous model[J]. Antimicrob Agents Chemother, 2013,57(5):2365-2368.
[39]UPPULURI P, SRINIVASAN A, RAMASUBRAMANIAN A,et al. Effects of fluconazole, amphotericin B, and caspofungin on Candida albicans biofilms under conditions of flow and on biofilm dispersion[J]. Antimicrob Agents Chemother, 2011, 55 (7):3591-3593.
[40]GUEMBE M, GUINEA J, MARCOS-ZAMBRANO LJ, et al. Micafungin at physiological serum concentrations shows antifungal activity against Candida albicans and Candida parapsilosis biofilms[J]. Antimicrob Agents Chemother, 2014,58(9):5581-5584.
[41]MELO AS, BIZERRA FC, FREYMULLER E, et al. Biofilm production and evaluation of antifungal susceptibility amongst clinical Candida spp. isolates, including strains of the Candida parapsilosis complex[J]. Med Mycol, 2011, 49(3):253-262.
[42]MUKHERJEE PK, LONG L, KIM HG, et al. Amphotericin B lipid complex is efficacious in the treatment of Candida albicans biofilms using a model of catheter-associated Candida biofilms[J]. Int J Antimicrob Agents, 2009, 33(2):149-153.
[43]趙蘭雪, 李德東, 王彥, 等. 抗白念珠菌生物被膜藥物的研究進展[J]. 中國真菌學雜志, 2013, 8(2):124-128.
[44]LI LN, GUO LH, LUX R, et al. Targeted antimicrobial therapy against Streptococcus mutans establishes protective noncariogenic oral biofilms and reduces subsequent infection[J]. Internat J Oral Sci, 2010, 2(2):66-73.
[45]DELATTIN N, DE BRUCKER K, VANDAMME K, et al. Repurposing as a means to increase the activity of amphotericin B and caspofungin against Candida albicans biofilms[J]. J Antimicrob Chemother, 2014, 69(4):1035-1044.
[46]CUELLAR-CRUZ M, VEGA-GONZALEZ A, MENDOZANOVELO B, et al. The effect of biomaterials and antifungals on biofilm formation by Candida species: a review[J]. Eur J Clin Microbiol Infect Dis, 2012, 31(10):2513-2527.
[47]BOUZA E, GUINEA J, GUEMBER M. The Role of Antifungals against Candida biofilm in catheter-related candidemia[J]. Antibiotics, 2015, 4(1):1-17.
Candida biofilm and antifungal strategies
LI Jianguo. (Intensive Care Unit, Zhongnan Hospital of Wuhan University, Wuhan 430071, China)
R379
A
1009-7708(2016)02-0235-06
10.16718/j.1009-7708.2016.02.020
2015-08-05
2015-10-12
武漢大學中南醫院重癥監護室; 武漢 430071。
李建國(1952—),男,碩士,主任醫師,教授,主要從事重癥醫學臨床和科研工作。
李建國,E-mail:drljg1817@163.com。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
科學大眾(2020年12期)2020-08-13 03:22:22
航空世界(2020年10期)2020-01-19 14:36:20
