畢赤酵母工程菌發(fā)酵木聚糖酶條件的響應(yīng)面優(yōu)化
2016-09-14 07:14:03王丹丹周晨妍朱新術(shù)李同彪新鄉(xiāng)醫(yī)學(xué)院生命科學(xué)技術(shù)學(xué)院河南新鄉(xiāng)453000新鄉(xiāng)醫(yī)學(xué)院三全學(xué)院河南新鄉(xiāng)453000
食品工業(yè)科技 2016年2期
王丹丹,周晨妍,朱新術(shù),李同彪(.新鄉(xiāng)醫(yī)學(xué)院生命科學(xué)技術(shù)學(xué)院,河南新鄉(xiāng)453000;2.新鄉(xiāng)醫(yī)學(xué)院三全學(xué)院,河南新鄉(xiāng)453000)
畢赤酵母工程菌發(fā)酵木聚糖酶條件的響應(yīng)面優(yōu)化
王丹丹1,2,周晨妍1,*,朱新術(shù)1,李同彪1
(1.新鄉(xiāng)醫(yī)學(xué)院生命科學(xué)技術(shù)學(xué)院,河南新鄉(xiāng)453000;2.新鄉(xiāng)醫(yī)學(xué)院三全學(xué)院,河南新鄉(xiāng)453000)
通過(guò)單因素實(shí)驗(yàn)研究了誘導(dǎo)溫度、種齡、pH、甲醇濃度以及誘導(dǎo)時(shí)間對(duì)畢赤酵母工程菌產(chǎn)木聚糖酶的影響。在此基礎(chǔ)上進(jìn)行響應(yīng)面優(yōu)化設(shè)計(jì)實(shí)驗(yàn),并根據(jù)結(jié)果擬合小二乘二次項(xiàng)回歸方程,探討了各因素對(duì)木聚糖酶比酶活力的影響。確定了最優(yōu)的發(fā)酵培養(yǎng)條件:種齡為31 h,誘導(dǎo)時(shí)間104 h,甲醇誘導(dǎo)濃度1%,發(fā)酵起始pH4.0,誘導(dǎo)溫度為30℃,在此條件下對(duì)實(shí)驗(yàn)結(jié)果進(jìn)行驗(yàn)證,得到木聚糖酶比酶活力為43526.3 U/mg。
畢赤酵母,木聚糖酶,發(fā)酵條件,響應(yīng)面
木聚糖酶是指專(zhuān)一降解木聚糖為寡木聚糖、低聚木糖、木二糖和木糖一類(lèi)酶的總稱(chēng)[1-3]。木聚糖酶在能源、食品、飼料、造紙等工業(yè)方面有著非常重要的應(yīng)用價(jià)值[4-5]。微生物發(fā)酵產(chǎn)木聚糖酶的過(guò)程中,不僅與微生物的種類(lèi)關(guān)系密切,還與培養(yǎng)基的組成以及培養(yǎng)條件關(guān)系也非常大。木聚糖酶的生產(chǎn)菌有很多,包括細(xì)菌[6-8]、霉菌[5,9-10]和放線菌等,但是這些野生型菌株生產(chǎn)木聚糖酶存在著產(chǎn)量不高,酶學(xué)性質(zhì)相對(duì)較差等缺點(diǎn),如周薇薇等[11]利用響應(yīng)面法優(yōu)化了黑曲霉(Aspergillus niger FIP-09-24)的固態(tài)發(fā)酵工藝,在最佳培養(yǎng)條件下培養(yǎng)酶活力只有66002 U/g;湯文晶等使用響應(yīng)面法優(yōu)化匍枝根霉最高酶活力為101.697 U/mL[12],故采用分子生物學(xué)手段對(duì)野生型木聚糖酶基因進(jìn)行改造,構(gòu)建工程菌是現(xiàn)在研究的熱點(diǎn),并在此基礎(chǔ)上對(duì)工程菌發(fā)酵產(chǎn)酶的條件進(jìn)行優(yōu)化,以期獲得更高產(chǎn)量的木聚糖酶。但是要實(shí)現(xiàn)工業(yè)生產(chǎn),使生產(chǎn)菌株的發(fā)酵產(chǎn)值達(dá)到一個(gè)更大的水平,必須采用更為經(jīng)濟(jì)合理的方法進(jìn)行研究。本研究在構(gòu)建畢赤酵母工程菌的基礎(chǔ)上,對(duì)其發(fā)酵條件進(jìn)行進(jìn)一步的優(yōu)化,旨在提高木聚糖酶的的產(chǎn)酶量,使其在工業(yè)上有更大的應(yīng)用價(jià)值。
1 材料與方法
1.1 材料與儀器
畢赤酵母工程菌GS115/pPIC9K-xynZF-2 新鄉(xiāng)醫(yī)學(xué)院生命科學(xué)技術(shù)學(xué)院酶與發(fā)酵工程實(shí)驗(yàn)室構(gòu)建保存;無(wú)氨基酸酵母氮源(YNB)、G418 Amresco公司;樺木木聚糖(Birch wood xylan) Sigma公司;牛血清蛋白 上海生工生物有限公司。
恒溫振蕩器(HZQ-F160A) 上海一恒科學(xué)儀器有限公司;高速冷凍離心機(jī)(Neofuge 23R) 上海力申科學(xué)儀器有限公司;電子恒溫水浴鍋(DZKW-4)北京中興偉業(yè)有限公司;微量分光光度計(jì)Nanodrop 2000 美國(guó)Thermo Fisher Scientific公司;往復(fù)式水浴恒溫振蕩器ZHWY-110X、曲線控制十段編程恒溫培養(yǎng)箱ZGP-A2080 上海智誠(chéng)分析儀器制造有限公司;不銹鋼立式壓力蒸汽滅菌器LX-C35L 合肥華泰醫(yī)療設(shè)備有限公司。
1.2 實(shí)驗(yàn)方法
1.2.1 培養(yǎng)基的配制 YPD、BMGY(生長(zhǎng)富集培養(yǎng)基)、BMMY(誘導(dǎo)表達(dá)培養(yǎng)基)配制方法見(jiàn)參考文獻(xiàn)[13]。
1.2.2 畢赤酵母工程菌的搖瓶培養(yǎng)基誘導(dǎo)表達(dá) 挑取保存的木聚糖酶高產(chǎn)菌株GS115/pPIC9K-xynZF-2,接種于含有20 mL BMGY培養(yǎng)基的250 mL的三角瓶中,30℃,250 r/min培養(yǎng)至A600為6.0左右,3000 r/min離心收集菌體,轉(zhuǎn)接至裝有30 mL BMMY培養(yǎng)基的250 mL三角瓶中,相同條件下繼續(xù)培養(yǎng),每24 h補(bǔ)加100%甲醇至終濃度為0.5%,誘導(dǎo)表達(dá)3 d。
1.2.3 粗酶液的制備 取誘導(dǎo)表達(dá)培養(yǎng)后的發(fā)酵液于3000 r/min離心10 min,取上清。
1.2.4 木聚糖比酶活力的測(cè)定 采用改進(jìn)的DNS法[14]測(cè)定木聚糖酶活力。酶活力單位的定義:在50℃和pH4.6條件下,以0.5%樺木木聚糖作為底物,以每分鐘產(chǎn)生1 μmol木糖所需的酶量為1個(gè)酶活力單位。以木糖作為標(biāo)準(zhǔn)樣品的標(biāo)準(zhǔn)曲線為y=2.0890x-0.2079,R2=0.9999。
用Bradford法[15]測(cè)定蛋白質(zhì)濃度,以牛血清白蛋白作為標(biāo)準(zhǔn)蛋白標(biāo)準(zhǔn)曲線為y=5.4200x+0.0328,R2= 0.9997。
比酶活力(U/mg)=酶活力/蛋白濃度。
1.2.5 發(fā)酵條件的單因素優(yōu)化
1.2.5.1 生長(zhǎng)曲線的繪制 挑取保存的木聚糖酶高產(chǎn)菌株GS115/pPIC9K-xynZF-2,接種于含有20 mL BMGY培養(yǎng)基的250 mL的三角瓶中,30℃,250 r/min培養(yǎng)48 h,每隔4 h取樣一次,將菌液稀釋后在波長(zhǎng)為600 nm條件下以空白培養(yǎng)基為對(duì)照進(jìn)行比色,測(cè)定吸光值A(chǔ)600,繪制細(xì)胞生長(zhǎng)曲線,其中A600=讀數(shù)×稀釋倍數(shù)。
1.2.5.2 誘導(dǎo)溫度對(duì)發(fā)酵產(chǎn)酶的影響 將GS115/ pPIC9K-xynZF-2,接種于含有20 mL的BMGY培養(yǎng)基中,30℃,250 r/min培養(yǎng)后接入30 mL的BMMY培養(yǎng)基中,于26、28、30、32、34、36℃條件下培養(yǎng),每隔24 h補(bǔ)加0.5%甲醇一次,培養(yǎng)3 d后制備粗酶液,測(cè)量比酶活力,確定最優(yōu)的誘導(dǎo)溫度。
1.2.5.3 種齡對(duì)發(fā)酵產(chǎn)酶的影響 將GS115/pPIC9K-xynZF-2,接種于含有20 mL BMGY培養(yǎng)基的250 mL的三角瓶中,30℃,250 r/min培養(yǎng)12、16、20、24、28、 32、36 h后離心轉(zhuǎn)接入30 mL的BMMY培養(yǎng)基中,最優(yōu)溫度下培養(yǎng)3 d,每隔24 h補(bǔ)加甲醇0.5%,制備粗酶液,測(cè)定比酶活力,確定最佳的種齡。
1.2.5.4 甲醇誘導(dǎo)時(shí)間對(duì)發(fā)酵產(chǎn)酶的影響 將GS115/ pPIC9K-xynZF-2接種于含有20 mL BMGY培養(yǎng)基的250 mL的三角瓶中,30℃,250 r/min培養(yǎng),按照最佳種齡接入30 mL BMMY培養(yǎng)基,最適溫度下繼續(xù)培養(yǎng),每隔24 h補(bǔ)加0.5%的甲醇一次,24、48、72、96、120、144、168 h后,制備粗酶液,測(cè)量比酶活力,確定最佳的誘導(dǎo)時(shí)間。
1.2.5.5 誘導(dǎo)起始pH對(duì)發(fā)酵產(chǎn)酶的影響 將GS115/ pPIC9K-xynZF-2接種于含有20 mL pH分別由4.5、5.0、5.5、6.0、6.5、7.0、7.5的緩沖液配制的BMGY培養(yǎng)基中,30℃,250 r/min培養(yǎng),按照最佳種齡接入30 mL 的BMMY培養(yǎng)基中,最適溫度下培養(yǎng),每隔24 h補(bǔ)加0.5%的甲醇一次,培養(yǎng)最優(yōu)誘導(dǎo)時(shí)間后,制備粗酶液,測(cè)量比酶活力,確定最優(yōu)的誘導(dǎo)表達(dá)起始pH。
1.2.5.6 甲醇誘導(dǎo)濃度對(duì)發(fā)酵產(chǎn)酶的影響 將GS115/ pPIC9K-xynZF-2接種于含有20 mL BMGY培養(yǎng)基的250 mL的三角瓶中,30℃,250 r/min培養(yǎng),按照最佳種齡接入30 mL的BMMY培養(yǎng)基中最適溫度下培養(yǎng),每隔24 h補(bǔ)加甲醇0.25%、0.5%、1.0%、1.5%、2.0%、2.5%、3.0%、3.5%,培養(yǎng)最優(yōu)誘導(dǎo)時(shí)間后,制備粗酶液,測(cè)量比酶活力,確定甲醇最優(yōu)的誘導(dǎo)濃度。
1.2.6 響應(yīng)面優(yōu)化設(shè)計(jì) 根據(jù)單因素實(shí)驗(yàn)結(jié)果,選用種齡、誘導(dǎo)起始pH、誘導(dǎo)時(shí)間、甲醇濃度為因素,采用Box-Behnken中心組合實(shí)驗(yàn)設(shè)計(jì),設(shè)計(jì)四因素五水平的響應(yīng)面實(shí)驗(yàn),因素水平表見(jiàn)表1。
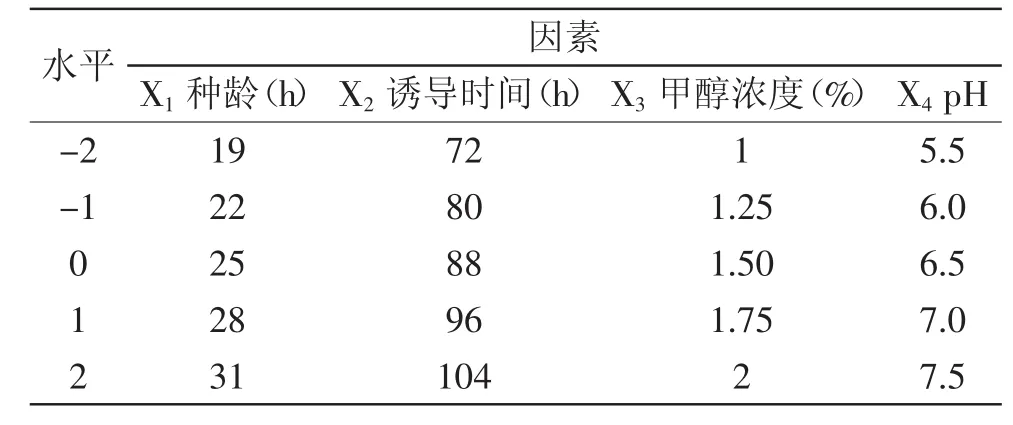
表1 實(shí)驗(yàn)因素水平表Table1 Experimental factors and levels table
1.2.7 統(tǒng)計(jì)學(xué)處理 每個(gè)實(shí)驗(yàn)進(jìn)行3次重復(fù),取其平均值,最終實(shí)驗(yàn)數(shù)據(jù)均用SPSS 11.0和Matlab-R2014a-Windows軟件進(jìn)行處理,并繪制出相應(yīng)的圖形。
2 結(jié)果與分析
2.1 畢赤酵母工程菌生長(zhǎng)曲線的繪制
以培養(yǎng)時(shí)間為橫坐標(biāo),A600為縱坐標(biāo)做GS115/ pPIC9K-xynZF-2生長(zhǎng)曲線圖。如圖1所示,16~36 h為對(duì)數(shù)生長(zhǎng)期,在28 h左右菌體達(dá)到對(duì)數(shù)生長(zhǎng)中期,對(duì)數(shù)生長(zhǎng)期以前由于菌體剛剛開(kāi)始生長(zhǎng),菌體含量過(guò)少,屬于生長(zhǎng)延遲期;超過(guò)對(duì)數(shù)生長(zhǎng)期,由于菌株的代謝產(chǎn)物增加以及營(yíng)養(yǎng)物質(zhì)的消耗,不適宜菌體生產(chǎn)大量目的產(chǎn)物。對(duì)數(shù)生長(zhǎng)期的菌株呈指數(shù)形式增長(zhǎng),繁殖速度快,菌體數(shù)量多,新陳代謝活躍,有利于進(jìn)行菌株的最優(yōu)化生產(chǎn),所以本實(shí)驗(yàn)先暫定選取對(duì)數(shù)生長(zhǎng)中期28 h菌體作為發(fā)酵用種子。

圖1 畢赤酵母工程菌生長(zhǎng)曲線圖Fig.1 Growth curve of recombinant yeast
2.2 單因素實(shí)驗(yàn)
2.2.1 誘導(dǎo)溫度對(duì)發(fā)酵產(chǎn)酶的影響 圖2顯示,在低于30℃時(shí),隨著溫度的上升,該工程菌的產(chǎn)酶量依次增加,并且在30℃時(shí)產(chǎn)酶量達(dá)到最大,溫度再次升高時(shí),產(chǎn)酶量成下降趨勢(shì)。溫度是微生物菌體生長(zhǎng)的重要環(huán)境條件之一,菌體的發(fā)酵過(guò)程就是菌體內(nèi)酶反應(yīng)的一個(gè)過(guò)程,酶反應(yīng)需要一定的溫度。溫度是通過(guò)影響微生物膜的液晶結(jié)構(gòu)、酶和蛋白質(zhì)的合成與活性,以及RNA的結(jié)構(gòu)和轉(zhuǎn)錄等影響微生物的生命活動(dòng)[16]。溫度過(guò)低,微生物的生長(zhǎng)素受到限制,表達(dá)過(guò)程也會(huì)受到影響;溫度過(guò)高會(huì)破壞微生物的生長(zhǎng)和表達(dá)過(guò)程,因此選擇合適的溫度非常重要,對(duì)于該工程菌選擇誘導(dǎo)溫度為30℃。
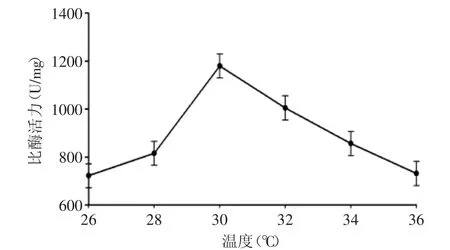
圖2 溫度對(duì)工程菌發(fā)酵產(chǎn)酶的影響Fig.2 Effect of temperature on xylanase production
2.2.2 種齡對(duì)發(fā)酵產(chǎn)酶的影響 將GS115/pPIC9K-xynZF-2進(jìn)行甲醇誘導(dǎo)表達(dá),并對(duì)其發(fā)酵條件進(jìn)行優(yōu)化。在發(fā)酵過(guò)程中,種子的選擇非常關(guān)鍵,種齡太短,發(fā)酵過(guò)程會(huì)出現(xiàn)前期生長(zhǎng)緩慢,致使周期延長(zhǎng),甚至造成異常發(fā)酵;種齡過(guò)長(zhǎng),會(huì)引起菌體過(guò)早自溶,導(dǎo)致生產(chǎn)能力下降。圖3結(jié)果顯示種齡選擇在對(duì)數(shù)生長(zhǎng)期的28 h(A600=16.0)時(shí),比酶活力最高。
2.2.3 甲醇誘導(dǎo)時(shí)間對(duì)發(fā)酵產(chǎn)酶的影響 圖4結(jié)果表明,誘導(dǎo)表達(dá)前期隨著時(shí)間的延長(zhǎng),表達(dá)量增大,在誘導(dǎo)96 h時(shí)達(dá)到最大;之后隨著時(shí)間的延長(zhǎng),產(chǎn)酶量依次遞減。在畢赤酵母中蛋白表達(dá)后被分泌到細(xì)胞外,可能影響培養(yǎng)液中營(yíng)養(yǎng)物質(zhì)的pH進(jìn)而影響木聚糖酶的表達(dá)。誘導(dǎo)時(shí)間過(guò)短時(shí),誘導(dǎo)劑還未充分發(fā)揮作用,培養(yǎng)基中營(yíng)養(yǎng)物質(zhì)并未大量消耗,啟動(dòng)子啟動(dòng)轉(zhuǎn)錄,蛋白表達(dá)量較少,比酶活力較低;誘導(dǎo)時(shí)間過(guò)長(zhǎng),培養(yǎng)液pH會(huì)發(fā)生明顯改變,并且隨著營(yíng)養(yǎng)物質(zhì)的大量消耗,目的蛋白的表達(dá)將會(huì)受阻,導(dǎo)致發(fā)酵液比酶活力下降。因此選擇96 h為該工程菌的最佳誘導(dǎo)時(shí)間。
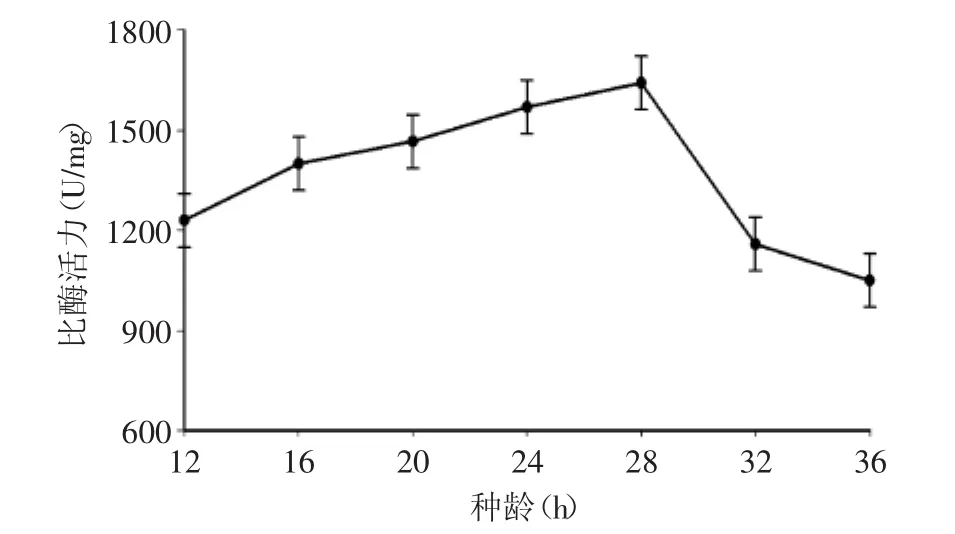
圖3 種齡對(duì)產(chǎn)酶的影響Fig.3 Effect of inoculation time on xylanase production
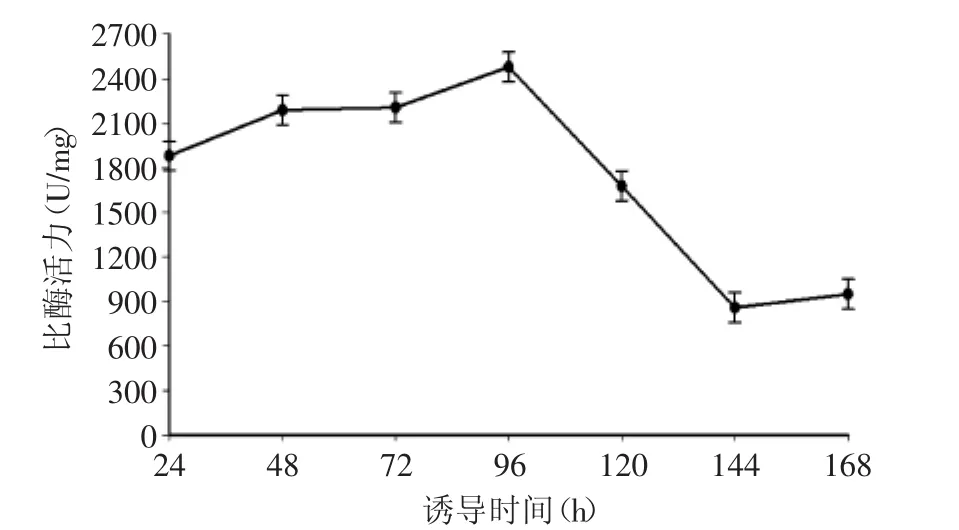
圖4 甲醇誘導(dǎo)時(shí)間對(duì)產(chǎn)酶的影響Fig.4 Effect of induction time on xylanase production
2.2.4 發(fā)酵起始pH對(duì)產(chǎn)酶的影響 圖5結(jié)果顯示,發(fā)酵液偏酸偏堿都會(huì)影響工程菌的表達(dá)。GS115/ pPIC9K-xynZF-2工程菌在pH為6.5時(shí)產(chǎn)酶量最高,pH偏高時(shí),畢赤酵母自身分泌的蛋白酶活力較高,可以分解產(chǎn)生的木聚糖酶[17];pH偏低時(shí),酸性太強(qiáng),重組木聚糖酶非常的不穩(wěn)定,所以該工程菌的發(fā)酵pH應(yīng)選擇在pH6.5。
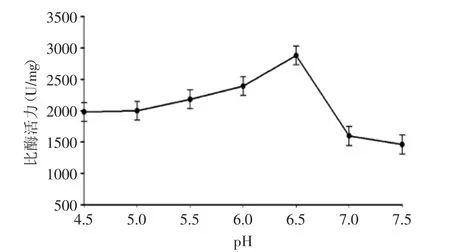
圖5 發(fā)酵起始pH對(duì)產(chǎn)酶的影響Fig.5 Effect of pH on xylanase production
2.2.5 誘導(dǎo)濃度對(duì)產(chǎn)酶的影響 甲醇能夠誘導(dǎo)重組畢赤酵母表達(dá),但是并不是隨著甲醇濃度的增加表達(dá)量就隨之增加,圖6結(jié)果表明,甲醇濃度在1.5%時(shí)誘導(dǎo)表達(dá)量最大,濃度過(guò)大對(duì)誘導(dǎo)表達(dá)反而起抑制作用。畢赤酵母的表達(dá)是以醇氧化酶啟動(dòng)子啟動(dòng)外源基因表達(dá)的,在只有甘油作為碳源的BMGY培養(yǎng)基中,甲醇氧化酶AOX1根本不會(huì)產(chǎn)生,外源基因也不會(huì)表達(dá)[18]。在以甲醇作為碳源的BMMY培養(yǎng)基中,一方面,畢赤酵母會(huì)表達(dá)醇氧化酶來(lái)利用甲醇,而木聚糖酶基因在醇氧化酶基因的下游,所以木聚糖酶的表達(dá)與醇氧化酶的表達(dá)成正比關(guān)系;另一方面,甲醇既可以作為酵母的碳源也可以作為誘導(dǎo)物,因此,在一定范圍內(nèi)提高甲醇的濃度可能會(huì)增加外源蛋白的表達(dá)量,但是,添加過(guò)量,甲醇對(duì)酵母生長(zhǎng)也會(huì)產(chǎn)生毒害作用,由圖6可以選擇1.5%為該工程菌的最優(yōu)甲醇濃度。

圖6 甲醇濃度對(duì)產(chǎn)酶的影響Fig.6 Effect of methanol concentration on xylanase production
2.3 響應(yīng)面實(shí)驗(yàn)設(shè)計(jì)優(yōu)化結(jié)果
2.3.1 方差分析 如表2所示是響應(yīng)面的結(jié)果,使用Matlab軟件對(duì)結(jié)果進(jìn)行統(tǒng)計(jì)學(xué)分析,對(duì)表中數(shù)據(jù)我們進(jìn)行了方差分析,見(jiàn)表3,在誤差允許范圍內(nèi),F(xiàn)值大于臨界值,四個(gè)因素的影響是顯著的,具有統(tǒng)計(jì)學(xué)意義。
對(duì)表中結(jié)果選用偏最小二乘回歸分析程序進(jìn)行分析,得到畢赤酵母工程菌發(fā)酵產(chǎn)木聚糖酶比酶活力(Y)對(duì)種齡(X1)、誘導(dǎo)時(shí)間(X2)、甲醇濃度(X3)、pH (X4)的二次多項(xiàng)式回歸模型:Y=6672.3+1663.8X1-1222.3X3-1017.2X4+876.9X+1540X+759.2X-1670X1X3-1043.8X1X4-1560.2X2X3。可以從誤差平方和看出該二次多項(xiàng)式回歸模型的擬合效果,當(dāng)潛變量個(gè)數(shù)為1時(shí),數(shù)據(jù)標(biāo)準(zhǔn)化后模型誤差平方和為0.7526,并且可以得出相應(yīng)組分時(shí)的模型擬合決定系數(shù)R2為0.9465,回歸模型的擬合度較好。
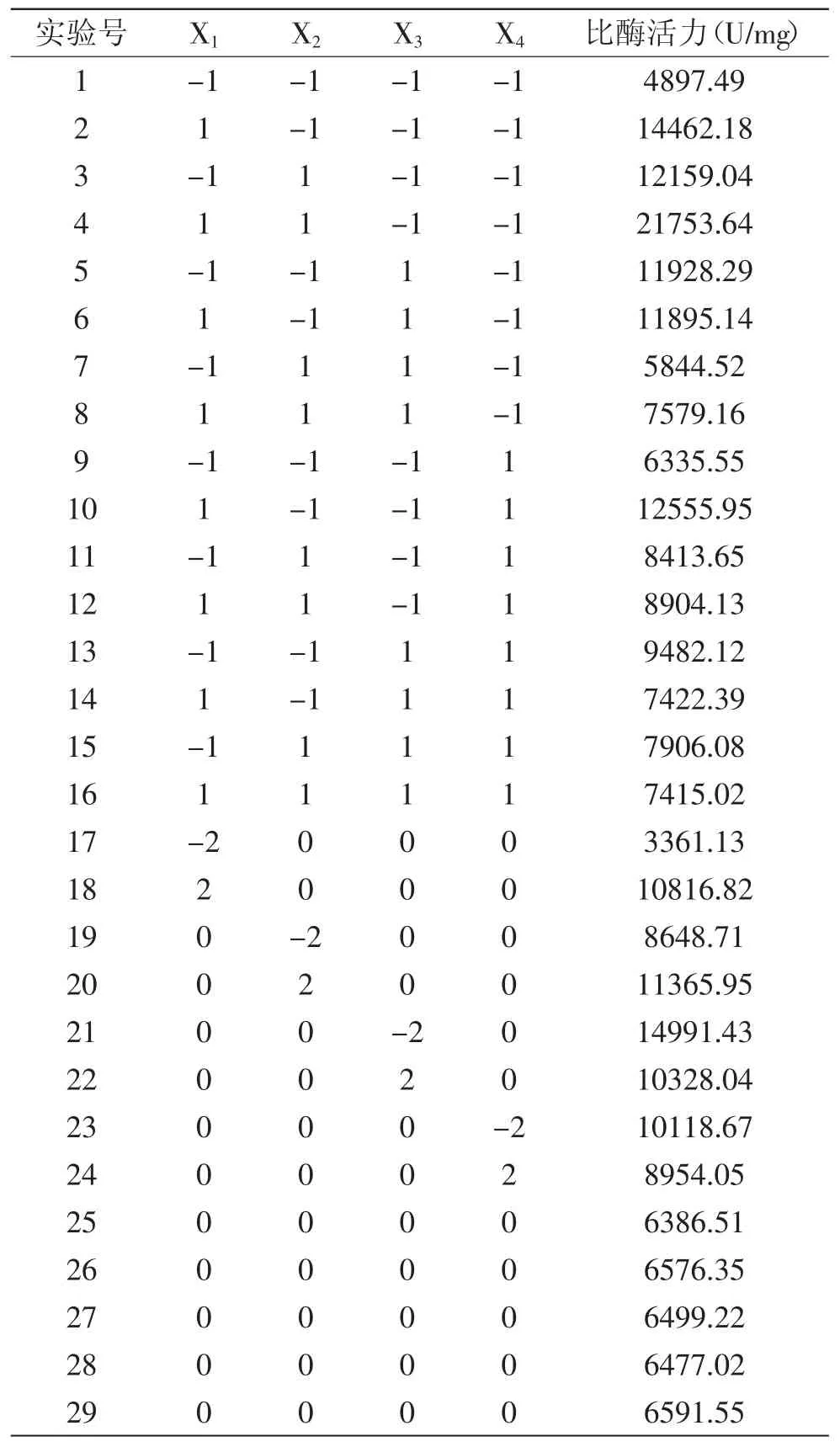
表2 響應(yīng)面分析實(shí)驗(yàn)結(jié)果Table2 Results of response surface analysis test
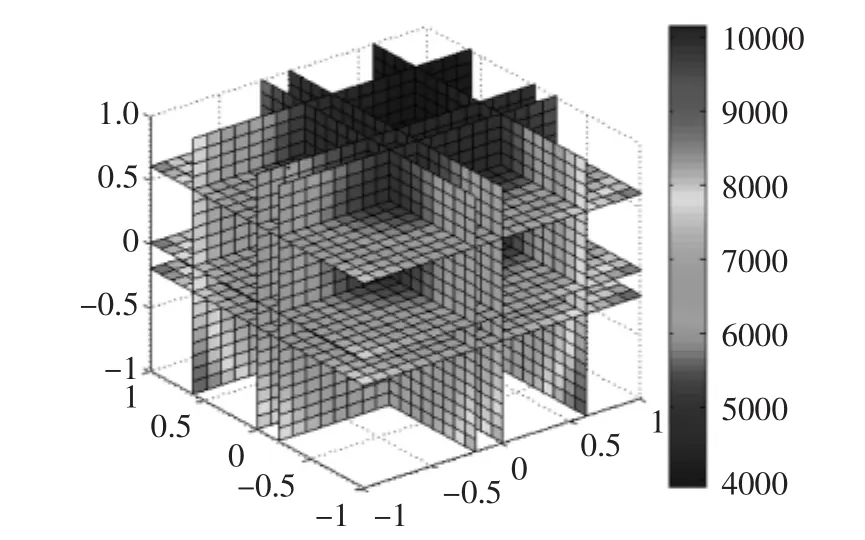
圖7 響應(yīng)面結(jié)果四維圖Fig.7 Four dimensional graph of response surface results

表3 方差分析Table3 Analysis of variance
基于該模型,可以以四維圖的形式顯示出來(lái),如圖7所示,從圖7中可以看出,四個(gè)因素在不同的水平對(duì)比酶活力均能產(chǎn)生不同的影響,最大值處于四維圖的邊緣,這給后期做進(jìn)一步的優(yōu)化提供了很好的基礎(chǔ)。根據(jù)設(shè)定的條件對(duì)回歸模型進(jìn)行優(yōu)化,在一定的實(shí)驗(yàn)范圍內(nèi)可以通過(guò)尋優(yōu)得到最優(yōu)的發(fā)酵工藝條件為:種齡為31 h,誘導(dǎo)時(shí)間104 h,甲醇誘導(dǎo)濃度1%,發(fā)酵起始pH4.0,模擬結(jié)果如圖8所示。根據(jù)此回歸方程可以得到最優(yōu)比酶活力可達(dá)44279 U/mg。
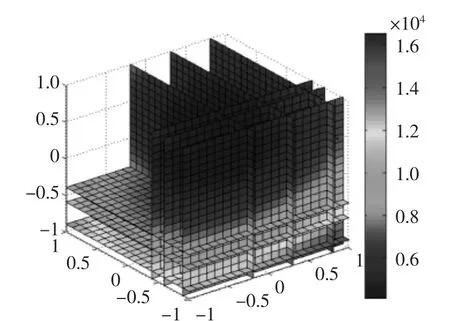
圖8 模擬結(jié)果四維圖Fig.8 The four dimensional map of analog result
2.3.2 驗(yàn)證性實(shí)驗(yàn) 根據(jù)所得到的最優(yōu)發(fā)酵工藝條件進(jìn)行驗(yàn)證性實(shí)驗(yàn),重復(fù)3次,得到實(shí)際比酶活力為43526.3 U/mg,大概占預(yù)測(cè)值的98.3%,說(shuō)明此回歸模型很好地反應(yīng)了實(shí)際情況。
3 結(jié)論
通過(guò)對(duì)畢赤酵母工程菌發(fā)酵條件單因素的優(yōu)化,確定各個(gè)因素的最優(yōu)水平,在此基礎(chǔ)上進(jìn)行Box-Behnken實(shí)驗(yàn)設(shè)計(jì),對(duì)工程菌發(fā)酵產(chǎn)木聚糖酶的發(fā)酵條件進(jìn)行優(yōu)化,以小二乘模型對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行優(yōu)化,得到影響木聚糖酶比酶活力的二次多項(xiàng)式回歸方程。最終的最優(yōu)發(fā)酵工藝條件為種齡為31 h,誘導(dǎo)時(shí)間104 h,甲醇誘導(dǎo)濃度1%,發(fā)酵起始pH4.0,預(yù)測(cè)值為44279 U/mg,對(duì)實(shí)驗(yàn)結(jié)果進(jìn)行驗(yàn)證性實(shí)驗(yàn),最終實(shí)際數(shù)值為43526.3 U/mg,與理論數(shù)值相差不大說(shuō)明該回歸模型合理可靠,與優(yōu)化前比酶活力1180 U/mg(圖2,30℃條件下培養(yǎng))相比提高了36倍多。本實(shí)驗(yàn)是在實(shí)驗(yàn)室水平上進(jìn)行的,后續(xù)研究可以考慮對(duì)發(fā)酵罐中試等擴(kuò)大培養(yǎng)條件進(jìn)行優(yōu)化,以使該結(jié)果在工業(yè)上得到更大的利用。
[1]吳萍,李正鵬,何慶元,等.金針菇固態(tài)發(fā)酵產(chǎn)木聚糖酶的純化及其性質(zhì)研究[J].藥用生物技術(shù),2012,19(1):31-34.
[2]符丹丹,李?lèi)?ài)江,謝慧,等.宇佐美曲霉木聚糖酶的純化及其性質(zhì)[J].食品科學(xué),2006,27(2):116-120.
[3]Coughlan M P,Hazlewood G P.β-1,4-D-xylan-degrading enzyme system:biochemistry,molecularbiology and applications [J].Biotechnol Apply Bioche,1993,17:259.
[4]孫振濤,趙祥穎,劉建軍,等.微生物木聚糖酶及其應(yīng)用[J].生物技術(shù),2007,17(2):93-97.
[5]劉波,鄔應(yīng)龍,張霞,等.紅曲霉固態(tài)發(fā)酵產(chǎn)木聚糖酶培養(yǎng)基的響應(yīng)面優(yōu)化[J].食品工業(yè)技,2014,35(1):254-258.
[6]Bajaj B K,Manhas K.Production and characterization of xylanase from Bacillus licheniformis P11(C)with potential for fruit juice and bakery industry[J].Biocatalysis and Agricultural Biotechnology,2011,1(4):330-337.
[7]Ko C H,Tsai C H,Tu J,et al.Identification of Paenibacillus sp.2S-6 and application of its xylanase on bio-bleaching[J].International Biodeterioration&Biodegradation,2011,65(2):334-339.
[8]Jiang Z Q,Cong Q Q,Yan Q J,et al.Characterisation of a thermostable xylanase from Chaetomium sp.and its application in Chinese steamed bread[J].Food Chemistry,2010,120(2):457-462.
[9]曹云鶴,陳小玲,賀平麗,等.硫色曲霉木聚糖酶基因xynA的克隆、表達(dá)及酶學(xué)性質(zhì)分析[J].生物技術(shù)通訊,2006,17(6):878-881.
[10]Shi H L,Yin X,Wu M C,et al.Cloning and bioinformatics analysis of an endoglucanase gene(Aucel12A)from Aspergillus usamii and its functional expression in Pichia pastoris[J].J Ind Microbiol Biotechnol,2012,39:347-357.
[11]周薇薇,尹亞輝,趙長(zhǎng)新.響應(yīng)面法優(yōu)化黑曲霉固態(tài)發(fā)酵產(chǎn)木聚糖酶工藝[J].大連工業(yè)大學(xué)學(xué)報(bào),2013,32(3):176-179.
[12]湯文晶,謝志皓,張慶慶,等.匍枝根霉液態(tài)發(fā)酵產(chǎn)木聚糖酶培養(yǎng)基優(yōu)化研究[J].安徽工程大學(xué)學(xué)報(bào),2014,29(1):1-5.
[13]Zhang H L,Yao B,Wang Y R.Expression of xylanase gene xynA from Stretomyces olivaceoviridis A1 in Escherichia coli and Pichia pastoris[J].China Biotechnology,2003,19(1):76-80.
[14]Yao D,Wang Y,Yang J,et al.Optimization of extraction process for xylan from corncob by response surface methodology [J].Food Science,2011,32(8):111-115.
[15]Bailey M J,Biely P,Poutanen K.Interlaboratory testing of methods for assay of xylanase activity[J].Journal of Biotechnology,1992,23:257-270.
[16]董悅安.溫度變化對(duì)地下水中微生物影響的研究[J].勘察科學(xué)技術(shù),2008(2):15-18.
[17]王濤,萬(wàn)紅貴,蔡恒,等.畢赤酵母發(fā)酵產(chǎn)木聚糖酶條件的研究[J].中國(guó)釀造,2009,20(8):86-89.
[18]周晨妍.宇佐美曲霉木聚糖酶基因的克隆、表達(dá)及定向誘變研究[D].無(wú)錫:江南大學(xué),2008.
Optimization of fermentation condition of Pichia pastoris for xylanase production by response analysis
WANG Dan-dan1,2,ZHOU Chen-yan1,*,ZHU Xin-shu1,LI Tong-biao1
(1.School of Life Science and Technology,Xinxiang Medical University,Xinxiang 453000,China;2.San Quan Medlcal College,Xinxiang 453000,China)
The influence of the temperature,inoculum time,pH,methanol concentration and induction time on the xylanase production of pichia recombinant bacteria were studied.The response surface optimazation was designed on the basis of the single factor experiment,and the small squares quadratic regression equation was fitted out according to the results to discuss the influence of the various factors on the xylanase enzyme activity.The optimum fermentation culture conditions were as follows:inoculum age 31 h,induction time 104 h,methanol concentration induced by 1%,fermentation starting pH4.0,induction temperature 30℃.On this condition,the result of the experiment for xylanase enzyme activity was 43526.3 U/mg.
Pichia pastoris;xylanase;fermentation conditions;response surface
TS201.1
A
1002-0306(2016)02-0194-05
10.13386/j.issn1002-0306.2016.02.031
2015-05-06
王丹丹(1988-),女,碩士研究生,助教,研究方向:微生物酶工程,E-mail:wangdan2988@126.com。
*通訊作者:周晨妍(1979-),女,副教授,研究方向:酶工程與發(fā)酵工程,E-mail:zhouchenyan2008@163.com。
河南省教育廳科學(xué)技術(shù)研究重點(diǎn)項(xiàng)目(13A180861);河南省高等學(xué)校青年骨干教師資助計(jì)劃項(xiàng)目(2011GGJS-125);新鄉(xiāng)醫(yī)學(xué)院科研項(xiàng)目培育基金(2013ZD113);新鄉(xiāng)醫(yī)學(xué)院研究生科研創(chuàng)新支持計(jì)劃項(xiàng)目資助(YJSCX201327Y)。
