長江中下游地區(qū)亞洲鐮刀菌NX-2毒素群體的檢測
2016-09-14 07:04:48許景升
植物保護 2016年2期
關(guān)鍵詞:檢測
羅 文, 張 昊, 許景升, 徐 進, 馮 潔
(中國農(nóng)業(yè)科學(xué)院植物保護研究所, 植物病蟲害生物學(xué)國家重點實驗室, 北京 100193)
?
長江中下游地區(qū)亞洲鐮刀菌NX-2毒素群體的檢測
羅文,張昊,許景升,徐進,馮潔*
(中國農(nóng)業(yè)科學(xué)院植物保護研究所, 植物病蟲害生物學(xué)國家重點實驗室, 北京100193)
由禾谷鐮刀菌復(fù)合種(Fusariumgraminearumspecies complex,FGSC)引起的麥類赤霉病,是農(nóng)業(yè)生產(chǎn)上的重要病害。為明確中國長江中下游冬小麥主產(chǎn)區(qū)小麥赤霉病菌種的構(gòu)成及其地理分布,對2008年從江蘇、浙江和湖北3省采集的656株小麥赤霉病菌株進行了分類鑒定。結(jié)果顯示,其中558個菌株為亞洲鐮刀菌(Fusariumasiaticum),98個菌株為禾谷鐮刀菌(Fusariumgraminearumsensu stricto),表明中國長江中下游冬小麥主產(chǎn)區(qū)小麥赤霉病的主要致病菌是亞洲鐮刀菌。選擇亞洲鐮刀菌(F.asiaticum)為研究對象,通過PCR-RFLP的方法對其進行產(chǎn)NX-2毒素菌株的檢測。結(jié)果沒有檢測到產(chǎn)NX-2毒素菌株,表明中國長江中下游冬小麥主產(chǎn)區(qū)并未出現(xiàn)NX-2毒素群體。本研究旨在了解NX-2毒素群體在中國長江中下游地區(qū)的地理分布,為進一步研究麥類赤霉病菌群體遺傳多樣性和演化趨勢奠定基礎(chǔ),為麥類赤霉病的防治和毒素污染的控制提供理論依據(jù)。
小麥赤霉病;亞洲鐮刀菌;NX-2
由禾谷鐮刀菌復(fù)合種(Fusariumgraminearumspecies complex,FGSC)引起的麥類赤霉病,是農(nóng)業(yè)生產(chǎn)上的重要病害,廣泛發(fā)生于世界濕潤和半濕潤地區(qū)[1-2]。近年來,隨著全球氣候變暖的加劇和麥田耕作制度的變化,赤霉病在亞洲、加拿大、歐洲和北美等地區(qū)頻繁發(fā)生,嚴(yán)重影響糧食產(chǎn)量,造成巨大的經(jīng)濟損失。
我國幅員遼闊,氣候多樣,再加上較為普遍的水稻、小麥和玉米輪作的種植制度,為赤霉病的發(fā)生提供了非常有利的條件。2012年,我國小麥赤霉病暴發(fā)程度歷史罕見,受害面積達到1 000萬hm2,約占全國小麥總面積的1/2。感病后的麥穗不僅籽粒皺縮,品質(zhì)下降[3],而且病菌產(chǎn)生的單端孢霉烯族毒素(trichothecenes)可抑制真核細(xì)胞蛋白質(zhì)的合成[4],破壞人和動物的免疫系統(tǒng),出現(xiàn)頭暈、嘔吐、口腔病損、炎癥、器官出血等癥狀,嚴(yán)重威脅人畜的健康安全[5]。
單端孢霉烯族毒素是結(jié)構(gòu)相似的倍半萜烯類化合物的總稱,根據(jù)化學(xué)結(jié)構(gòu)和產(chǎn)毒菌株來源不同,可分為A、B、C、D 4種毒素類型。其中B型單端孢霉烯族毒素(B-trichothecenes)是感染麥類赤霉病菌的病穗中積累的最主要的毒素,包括脫氧雪腐鐮刀菌烯醇(deoxynivalenol,DON)及其衍生物3-乙酰基脫氧雪腐鐮刀菌烯醇(3-acetyldeoxynivalenol,3-ADON)、15-乙酰基脫氧雪腐鐮刀菌烯醇(15-acetyldeoxynivalenol,15-ADON)和雪腐鐮刀菌烯醇(nivalenol,NIV)。
在中國流行危害的禾谷鐮刀菌復(fù)合種(FGSC)主要是禾谷鐮刀菌(Fusariumgraminearumsensu stricto)和亞洲鐮刀菌(Fusariumasiaticum),其中黃河流域以北地區(qū)以禾谷鐮刀菌為主,長江中下游地區(qū)以亞洲鐮刀菌為主[6-8]。Zhang等研究發(fā)現(xiàn)長江中下游麥區(qū)的亞洲鐮刀菌主要產(chǎn)生3-ADON,而禾谷鐮刀菌只產(chǎn)生15-ADON毒素[8]。
2014年,Liang等在美國的明尼蘇達州分離到一種禾谷鐮刀菌[9],這種禾谷鐮刀菌早在2010年Gale等人就報道過,發(fā)現(xiàn)它不產(chǎn)生B型單端孢霉烯族毒素中的任何一種類型的毒素[10],而產(chǎn)生一種新的毒素(3α-乙酰基-7α,15-二羥基-12,13-環(huán)氧單端孢霉烯-9烯),Varga等人將其命名為NX-2[11]。Liang等將NX-2與B型單端孢霉烯族毒素中的DON、3-ADON、15-ADON和NIV的化學(xué)結(jié)構(gòu)進行比較,發(fā)現(xiàn)NX-2與3-ADON的化學(xué)結(jié)構(gòu)有很高的一致性,只在C-8位缺了一個酮基[9](圖1)。
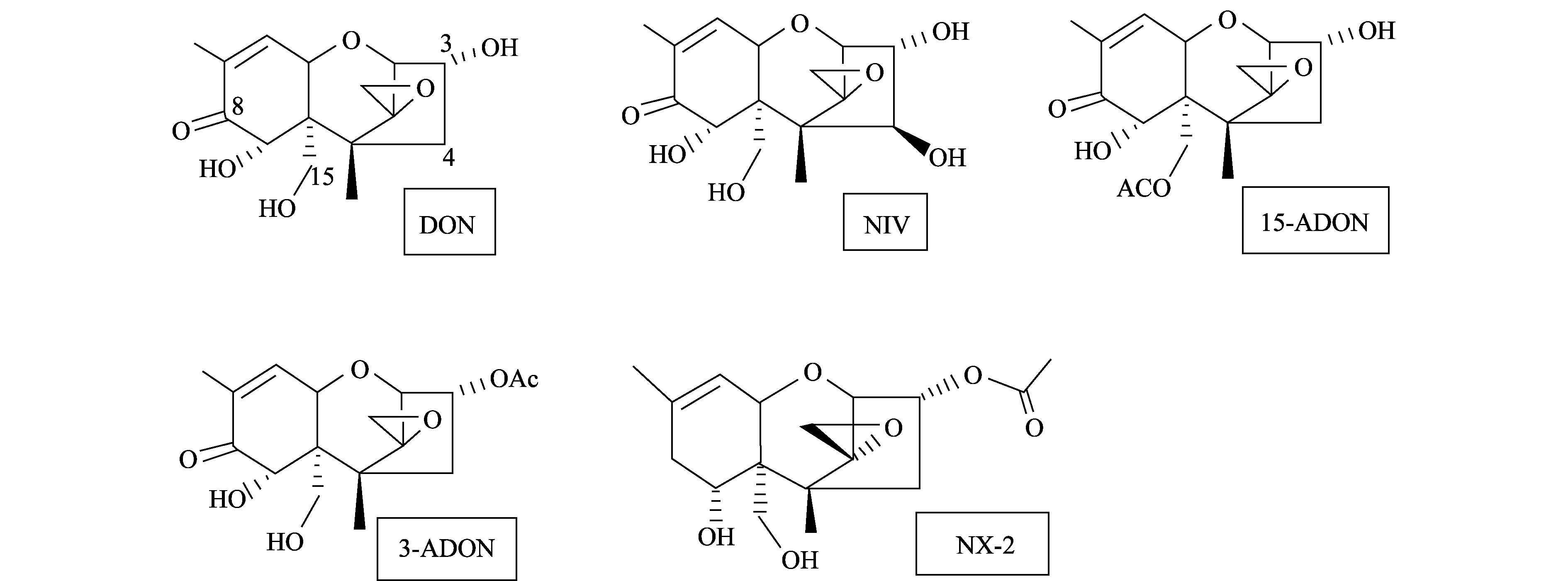
圖1 B型單端孢霉烯族毒素和NX-2毒素的化學(xué)結(jié)構(gòu)Fig.1 Chemotype of toxins B-trichothecenes and NX-2
McCormick等研究發(fā)現(xiàn)擬枝孢鐮刀菌(Fusariumsporotrichioides)和禾谷鐮刀菌(F.graminearums. str.)的Tri1基因都能編碼催化單端孢霉烯族毒素羥基化的酶[12]。FsTri1編碼細(xì)胞色素P450單氧酶,催化A型單端孢霉烯族毒素(A-trichothecenes)C-8位羥基化;而FgTri1編碼細(xì)胞色素P450加氧酶,催化B型單端孢霉烯族毒素(B-trichothecenes)C-7位和C-8位羥基化,使C-7位生成一個羥基,C-8位生成一個酮基[13-14]。新發(fā)現(xiàn)的NX-2型毒素僅在C-7位生成一個羥基,推測產(chǎn)NX-2型毒素菌株的FgTri1等位基因可能編碼一種細(xì)胞色素P450單氧酶,只催化C-7位羥基化。Liang等將產(chǎn)生NX-2的菌株與產(chǎn)生DON的菌株的Tri1序列進行比對分析,設(shè)計出1對引物Tri1F/Tri1R[9],采用PCR-RFLP檢測禾谷鐮刀菌中產(chǎn)NX-2毒素的菌株,結(jié)果共檢測到產(chǎn)NX-2毒素菌株13株,占總數(shù)的2.8%,并發(fā)現(xiàn)若按照原來基于Tri3和Tri12基因的分子檢測方法,這些產(chǎn)生NX-2的菌株均被誤認(rèn)為是3-ADON化學(xué)型[9]。Liang等對3-ADON毒素群體的地理分布及出現(xiàn)頻率進行分析發(fā)現(xiàn),NX-2毒素群體與3-ADON群體表現(xiàn)出相似的動態(tài)變化規(guī)律[9],推測在NX-2毒素群體與3-ADON毒素群體間可能存在一定的聯(lián)系。
Ward等的研究發(fā)現(xiàn),與15-ADON毒素群體相比,3-ADON群體能產(chǎn)生更多的毒素,表現(xiàn)出更強的繁殖能力和更快的生長速率[15]。說明3-ADON毒素群體具有更強的生態(tài)適應(yīng)性,有取代15-ADON群體的趨勢。因此,盡管Liang等在美國發(fā)現(xiàn)的NX-2毒素群體所占比例很小(2.8%),但是它的發(fā)現(xiàn)是否意味著一種具有更強生態(tài)適應(yīng)性的種群的出現(xiàn),是否會對農(nóng)業(yè)生產(chǎn)帶來更多的威脅,需要我們進一步研究。
目前,在中國尚未有NX-2毒素群體的相關(guān)報道。本研究于2008年從中國冬小麥主產(chǎn)區(qū)江蘇、浙江和湖北采集大量菌株,對這些菌株進行了分類鑒定。選擇其中主要產(chǎn)生3-ADON毒素的亞洲鐮刀菌(F.asiaticum)為研究對象,檢測其是否為產(chǎn)NX-2毒素的菌株,旨在了解NX-2毒素群體在中國長江中下游地區(qū)的地理分布,為揭示小麥赤霉菌群體演化趨勢,研發(fā)新的綜合控制途徑等提供理論依據(jù)。
1 材料與方法
1.1小麥赤霉病菌的分離和培養(yǎng)
2008年,從江蘇、浙江和湖北3省的冬小麥田中采集感染小麥赤霉病的病穗,進行病原真菌的單孢分離[16],并對分離的菌株進行純化及初步的表型鑒定,共獲得菌株656個。將供試菌株接種于PDA平板上,28℃條件下培養(yǎng)。

表1 供試菌株信息Table 1 Information of strains used in this study
1.2真菌基因組DNA的提取
供試菌株長滿平皿后,刮取菌絲,用真菌基因組DNA提取試劑盒D3390- 02(Omega,美國)提取其基因組DNA。
1.3菌株的鑒定
利用引物Fg16F(5′-CTCCGGATATGTTGCGTCAA-3′)和Fg16R(5′-GGTAGGTATCCGACATGGCAA-3′)[17]進行PCR擴增,PCR反應(yīng)體系為:真菌DNA 約20 ng、2×TaqMix 10 μL、10 μmol/L Fg16F/Fg16R 各1 μL、加ddH2O至20 μL;PCR反應(yīng)程序為: 95℃預(yù)變性 3 min;95℃變性30 s,52℃退火30 s,72℃延伸30 s,30個循環(huán);72℃延伸10 min。擴增產(chǎn)物采用1.5%的瓊脂糖凝膠電泳檢測,其中禾谷鐮刀菌擴增片段大小為400 bp;亞洲鐮刀菌為500 bp。
1.4亞洲鐮刀菌(F.asiaticum)Tri1基因測序
利用引物Tri16-IF1(5′-GCCTSATAGCGACGATCTTGC-3′)和Fg_Tri1-R1(5′-AACAAGTGGCGAGATCAAACC-3′)[9],以亞洲鐮刀菌(F.asiaticum)DNA為模板,進行PCR擴增,獲得含有FaTri1基因序列的PCR產(chǎn)物。
PCR反應(yīng)體系為:DNA模板約20 ng、PrimeSTAR HS (Premix)25 μL、10 μmol/L Tri16-IF1 1 μL、10 μmol/L Fg_Tri1-R1 1 μL、加ddH2O至50 μL。PCR反應(yīng)程序為:96℃預(yù)變性2 min;94℃變性 30 s;53℃退火30 s,68℃延伸2 min,35個循環(huán);72℃延伸10 min、產(chǎn)物4℃保存。擴增產(chǎn)物采用1%的瓊脂糖凝膠電泳檢測,目的條帶送上海生物工程有限公司測序。
將測序反饋的含有FaTri1基因的序列與FgTri1基因序列進行比對,發(fā)現(xiàn)存在少數(shù)SNP位點,因此,已報道的引物Tri1F(5′-ATGGCTCTCATCACCAG-3′)和Tri1R(5′-CAATTCCAATCGCAGACAA-3′)[9]無法成功擴增F.asiaticum菌株。根據(jù)FaTri1基因序列設(shè)計亞洲鐮刀菌特異的FaTri1基因正向引物FaTri1F(5′-ATGGCTATCATCAGCAAC-3′),反向引物仍為FgTri1基因序列的反向引物Tri1R。采用F.asiaticum標(biāo)準(zhǔn)菌株進行驗證。
1.5產(chǎn)NX-2毒素菌株的檢測
采用PCR-RFLP檢測亞洲鐮刀菌中產(chǎn)NX-2毒素的菌株。PCR反應(yīng)體系為:模板DNA約20 ng、2×TaqMix 10 μL、10 μmol/L FaTri1F 1 μL、10 μmol/L Tri1R 1 μL,加ddH2O至20 μL。PCR反應(yīng)程序為:94℃預(yù)變性2 min;94℃變性1 min,56℃退火1 min,72℃延伸1 min 45 s,25個循環(huán);72℃延伸10 min。產(chǎn)物4℃保存。擴增產(chǎn)物采用1%的瓊脂糖凝膠電泳檢測,檢測到目的條帶后,對擴增產(chǎn)物進行酶切驗證。酶切體系為:擴增產(chǎn)物10 μL、ApoⅠ(10U/μL)0.15 μL、100×BSA 0.15 μL、10×NEB buffer 1.5 μL、加ddH2O至15 μL,反應(yīng)液置于50℃,水浴2 h。酶切產(chǎn)物采用1.5%的瓊脂糖凝膠電泳檢測。
2 結(jié)果與分析
2.1菌株的鑒定結(jié)果
利用引物Fg16F/Fg16R對656個菌株進行了PCR擴增,其中558個菌株能夠穩(wěn)定地擴增出500 bp的片段,屬于亞洲鐮刀菌(F.asiaticum)。另有98個
菌株能夠穩(wěn)定地擴增出400 bp的片段,屬于禾谷鐮刀菌(F.graminearums.str.)(表2,圖2)。其中從江蘇省分離的189個菌株中,有166個為F.asiaticum,23個為F.graminearums.str;從浙江省分離的188個菌株中,有172個為F.asiaticum,16個為F.graminearums.str;從湖北省分離的279個菌株中,有220個為F.asiaticum,59個為F.graminearums.str,表明長江中下游地區(qū)以F.asiaticum為主。
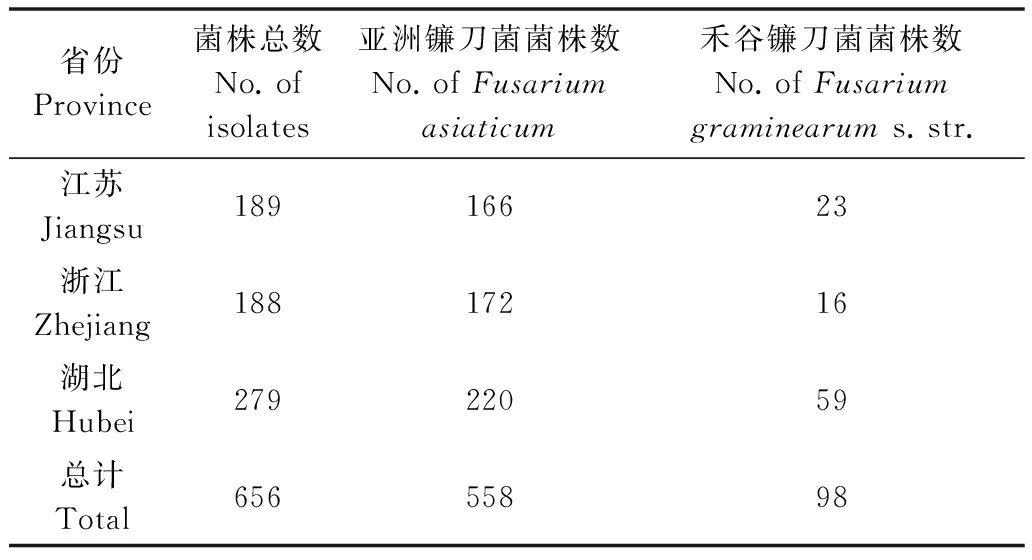
表2 利用引物Fg16F/Fg16R對供試菌株的鑒定結(jié)果Table 2 Identification result of strains using primers Fg16F/Fg16R in this study

圖2 表1中部分菌株的Fg16F/Fg16R擴增產(chǎn)物Fig.2 PCR products of part of strains in table 1 using primers Fg16F/Fg16R
2.2NX-2毒素菌株的檢測結(jié)果
采用PCR-RFLP,用引物FaTri1F和Tri1R對558個亞洲鐮刀菌(F.asiaticum)、3-ADON型F.asiaticum標(biāo)準(zhǔn)菌株NRRL6101和NIV型F.asiaticum標(biāo)準(zhǔn)菌株NRRL13818的Tri1基因進行PCR擴增,擴增產(chǎn)物經(jīng)ApoⅠ酶切。產(chǎn)3-ADON和NIV毒素的菌株,其擴增產(chǎn)物酶切后產(chǎn)生888 bp和851 bp 2個片段,但由于這2個片段長度只相差37 bp,無法被瓊脂糖凝膠分離,因此電泳后只顯示出一條帶;而產(chǎn)NX-2毒素的菌株,則會出現(xiàn)407、482和851 bp 3個片段[9]。酶切結(jié)果顯示,標(biāo)準(zhǔn)菌株和558個被測菌株均能夠穩(wěn)定的擴增出一條接近1 740 bp的片段(圖3~4),但經(jīng)ApoⅠ酶切后標(biāo)準(zhǔn)菌株和558個被測菌株均產(chǎn)生888 bp和851 bp的片段(圖5),表明江蘇、浙江和湖北3省未出現(xiàn)產(chǎn)NX-2毒素的菌株。

圖3 采用引物FaTri1F和Tri1R檢測供試 Fusarium asiaticum菌株Tri1基因Fig.3 Detection result of Tri1 in Fusarium asiaticum strains using primers FaTri1F and Tri1R

圖4 采用FaTri1F和Tri1R引物檢測部分湖北Fusarium asiaticum菌株的Tri1基因Fig.4 Detection result of Tri1 in Fusarium asiaticum strains from Hubei using primers FaTri1F and Tri1R

圖5 Fusarium asiaticum菌株Tri1基因的PCR-RFLP結(jié)果Fig.5 PCR-RFLP result of Tri1 gene in Fusarium asiaticum strains
3 討論
NX-2是最近在美國明尼蘇達州發(fā)現(xiàn)的由禾谷鐮刀菌產(chǎn)生的一種新的毒素,其結(jié)構(gòu)與3-ADON毒素有很高的一致性,只在C-8位缺了一個酮基[9]。研究發(fā)現(xiàn),Tri1基因在不同鐮刀菌種中的序列差異很大,在擬枝孢鐮刀菌(F.sporotrichioides)中,FsTri1編碼的酶催化A型單端孢霉烯族毒素(A-trichothecenes)C-8位羥基化,而在禾谷鐮刀菌(F.graminearums. str.)中,FgTri1編碼的酶催化B型單端孢霉烯族毒素(B-trichothecenes)C-7位和C-8位羥基化[13-14]。Liang等發(fā)現(xiàn)的NX-2型毒素菌株的Tri1基因編碼的酶僅催化C-7位羥基化,而不作用于C-8位,推測Tri1基因的多態(tài)性可能與NX-2毒素合成有關(guān)。而原有毒素化學(xué)型檢測方法無法區(qū)分NX-2與3-ADON化學(xué)型,并且NX-2毒素群體波動起伏的頻率與3-ADON毒素群體相似,推測NX-2毒素群體和3-ADON毒素群體間可能存在某種聯(lián)系[9]。Ward等研究發(fā)現(xiàn),北美產(chǎn)3-ADON毒素的F.graminearums. str.群體由東向西擴張,逐漸取代15-ADON群體[15]。這說明NX-2群體也可能通過同樣的方式進行群體擴張。在中國也發(fā)現(xiàn)了類似的情況,在長江中下游地區(qū)小麥赤霉病菌的優(yōu)勢種群是產(chǎn)3-ADON毒素的亞洲鐮刀菌(F.asiaticum),其具有取代NIV群體的擴張趨勢[7]。中國是否存在NX-2型菌株,其與3-ADON群體有何關(guān)系,目前尚無相關(guān)報道。由于NX-2毒素群體與3-ADON群體在基因型和流行趨勢方面十分相似,對主要由3-ADON化學(xué)型組成的亞洲鐮刀菌(F.asiaticum)群體進行篩選鑒定具有更強的針對性。通過F.asiaticum與F.graminearums. str.的Tri1基因序列比對分析,發(fā)現(xiàn)存在少數(shù)SNP位點,因此已報道的引物無法成功擴增F.asiaticum菌株。我們重新設(shè)計了FaTri1基因特異性引物FaTri1F,改進了Liang等的PCR-RFLP的方法。并對長江中下游地區(qū)江蘇、浙江、湖北3省是否存在NX-2群體進行了檢測。
本研究對2008年分離自中國冬小麥主產(chǎn)區(qū)江蘇、浙江、湖北3省的656株赤霉病菌株進行了鑒定和分析。研究結(jié)果表明,長江中下游地區(qū)江蘇、浙江、湖北3省的赤霉病菌由亞洲鐮刀菌(F.asiaticum)和禾谷鐮刀菌(F.graminearums. str.)組成,其中亞洲鐮刀菌是優(yōu)勢群體,與之前的報道一致。
在產(chǎn)NX-2毒素菌株的檢測中,江蘇、浙江、湖北3省的亞洲鐮刀菌(F.asiaticum)中都未檢測出產(chǎn)NX-2毒素菌株,表明這3省都還沒有出現(xiàn)產(chǎn)NX-2毒素菌株。但是不少研究都表明,與其他B型單端孢霉烯族毒素比較,3-ADON毒素群體能夠產(chǎn)生更多的毒素,具有更強的繁殖能力和更快的生長速率,并且它的群體正逐漸擴張。而Liang等的研究表明,NX-2毒素群體的出現(xiàn)和3-ADON毒素群體間有一定相關(guān)性[9]。目前,NX-2毒素群體的生長特性,產(chǎn)毒量和致病力并沒有相關(guān)的報道。但是若NX-2毒素群體與3-ADON毒素群體一樣,表現(xiàn)出更強的生態(tài)適應(yīng)性,那么它將在基因組水平上具有潛在的擴張趨勢,可能給世界麥類作物造成嚴(yán)重的危害。因此,加強NX-2毒素群體時空動態(tài)變化的監(jiān)測對小麥赤霉病菌防治十分重要。
本研究采用PCR-RFLP對長江中下游地區(qū)的江蘇、浙江、湖北3省是否存在產(chǎn)NX-2毒素菌株進行了檢測,明確了我國長江中下游冬小麥主產(chǎn)區(qū)小麥赤霉病菌種的構(gòu)成及其地理分布,發(fā)現(xiàn)了需要重點監(jiān)控的群體,為進一步研究麥類赤霉病菌群體遺傳多樣性和演化趨勢奠定了基礎(chǔ),也為麥類赤霉病的防治和毒素污染的控制提供了理論依據(jù)。
[1]Miedaner T, Cumagun C J R, Chakraborty S. Population genetics of three important head blight pathogensFusariumgraminearum,F.pseudograminearumandF.culmorum[J]. Journal of Phytopathology, 2008, 156(3): 129-139.
[2]Nganje W E, Bangsund D A, Leistritz F L, et al. Estimating the economic impact of a crop disease: the case ofFusariumhead blight in U.S [C]∥Wheat and Barley. National Fusarium Head Blight Forum Proceedings, 2002:275-281.
[3]Goswami R S, Kistler H C. Heading for disaster:Fusariumgraminearumon cereal crops [J]. Molecular Plant Pathology, 2004, 5(6): 515-525.
[4]Ueno Y, Nakajima M, Sakai K, et al. Comparative toxicology of trichothec mycotoxins: inhibition of protein synthesis in animal cells[J]. Journal of Biochemistry, 1973, 74(2): 285-296.
[5]Pestka J J, Smolinski A T. Deoxynivalenol toxicology and potential effects on humans [J]. Journal of Toxicology and Environmental Health, Part B Critical Reviews, 2005, 8(1): 39-69.
[6]Yang L, van der Lee T, Yang X, et al.Fusariumpopulations on Chinese barley show a dramatic gradient in mycotoxin profiles [J]. Phytopathology, 2008, 98(6): 719-727.
[7]Zhang Hao, Van der Lee T, Waalwijk C, et al. Population analysis of theFusariumgraminearumspecies complex from wheat in China show a shift to more aggressive isolates[J]. PLoS ONE, 2012, 7(2): e31722.
[8]Zhang Jingbo, Li Heping, Dang Fujun, et al. Determination of the trichothecene mycotoxin chemotypes and associated geographical distribution and phylogenetic species of theFusariumgraminearumclade from China[J]. Mycological Research, 2007, 111: 967-975.
[9]Liang J M, Xayamongkhon H, Broz K, et al. Temporal dynamics and population genetic structure ofFusariumgraminearumin the upper Midwestern United States [J]. Fungal Genetics and Biology, 2014, 73: 83-92.
[10]Gale L R, Ward T J, Kistler H C. A subset of the newly discovered Northland population ofFusariumgraminearumfrom the U.S does not produce the B-type trichothecenes DON, 15ADON, 3ADON or NIV in U.S [C]∥Wheat and Barley Scab Initiative. Proceedings of the 2010 National Fusarium Head Blight Forum,Session 2:Pathogen Biology & Genetics, Hyatt Regency Milwaukee, WI USA, 2010: 48-49.
[11]Varga E, Wiesenberger G, Hametner C, et al. New tricks of an old enemy: Isolates ofFusariumgraminearumproduce a novel type A trichothecene mycotoxin [J]. Environmental Microbiology, 2015, 17(8):2588-2600.
[12]McCormick S P, Alexander N J, Proctor R H. Heterologous expression of two trichothecene P450 genes inFusariumverticillioides[J]. Canadian Journal of Microbiology, 2006, 52(3): 220-226.
[13]Alexander N J, McCormick S P, Waalwijk C, et al. The genetic basis for 3-ADON and 15-ADON trichothecene chemotypes inFusarium[J].Fungal Genetics and Biology,2011,48(5):485-495.
[14]McCormick S P, Harris L J, Alexander N J, et al.Tri1 inFusariumgraminearumencodes a P450 oxygenase [J].Applied and Environmental Microbiology,2004,70(4):2044-2051.
[15]Ward T J, Clear R M, Rooney A P, et al. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenicFusariumgraminearumin North America [J]. Fungal Genetics and Biology, 2008, 45(4): 473-484.
[16]張昊, 張爭, 許景升,等. 一種簡單快速的赤霉病菌單孢分離方法-平板稀釋畫線分離法[J]. 植物保護, 2008, 34(6): 134-136.
[17]Nicholson P, Simpson D R, Weston G, et al. Detection and quantification ofFusariumculmorumandFusariumgraminearumin cereals using PCR assays [J]. Physiological and Molecular Plant Pathology, 1998, 53(1): 17-37.
(責(zé)任編輯:楊明麗)
Detection of NX-2 producingFusariumasiaticumstrains in the Middle and Lower Reaches of Yangtze River
Luo Wen,Zhang Hao,Xu Jingsheng,Xu Jin,Feng Jie
(State Key Laboratory for Biology of Plant Diseases and Insect Pests,Institute of Plant Protection,Chinese Academy of Agricultural Sciences, Beijing100193, China)
Fusariumhead blight (FHB)caused byFusariumgraminearumspecies complex is a very important disease in agricultural production. To analyze the composition and geographic distribution ofF.graminearumspecies complex in the main areas of wheat production in the Middle and Lower Reaches of Yangtze River in China, wheat spikes with FHB symptoms were collected from 3 provinces including Jiangsu, Zhejiang and Hubei in 2008. A total of 656 strains were isolated. The results showed that 558 strains wereF.asiaticumand 98 strains wereF.graminearums. str among the 656 isolates. Population ofF.asiaticumis predominant.F.asiaticumwas selected to detect NX-2 producing strains through the PCR-RFLP method. The results showed that there were no NX-2 producing strains in the main areas of wheat production in the Middle and Lower Reaches of Yangtze River in China. The aim of this research is to get a detailed geographic distribution of NX-2 producing strains in the Middle and Lower Reaches of Yangtze River in China, lay the foundation for further study of the population genetic diversity, and provide theoretical basis for disease and mycotoxin control.
Fusariumhead blight;Fusariumasiaticum;NX-2
2015-01-14
2015-03-12
國家自然科學(xué)基金(31201477);國家國際合作專項(2013DFG31930);公益性行業(yè)(農(nóng)業(yè))科研專項(201303016)
E-mail: jfeng@ippcaas.cn
S 435.12
A
10.3969/j.issn.0529-1542.2016.02.035
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
