‘KRS’芒果發(fā)育過程中糖積累與代謝相關(guān)酶活性的關(guān)系
2016-08-11 11:22:30武紅霞姚全勝王松標馬小衛(wèi)詹儒林
安徽農(nóng)業(yè)科學 2016年17期
武紅霞,姚全勝,王松標,馬小衛(wèi),詹儒林
(中國熱帶農(nóng)業(yè)科學院南亞熱帶作物研究所,農(nóng)業(yè)部熱帶果樹生物學重點實驗室,廣東湛江 524091)
?
‘KRS’芒果發(fā)育過程中糖積累與代謝相關(guān)酶活性的關(guān)系
武紅霞,姚全勝,王松標,馬小衛(wèi),詹儒林
(中國熱帶農(nóng)業(yè)科學院南亞熱帶作物研究所,農(nóng)業(yè)部熱帶果樹生物學重點實驗室,廣東湛江 524091)
[目的]探討芒果果實糖積累和轉(zhuǎn)化的生理機制。[方法]以‘KRS’芒為試材,研究芒果果實生長發(fā)育和成熟過程中的淀粉、蔗糖、葡萄糖和果糖含量變化,與淀粉酶、蔗糖代謝相關(guān)酶——酸性轉(zhuǎn)化酶(AI)、中性轉(zhuǎn)化酶(NI)、蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)活性的相關(guān)性。[結(jié)果]果實發(fā)育前期‘KRS’主要積累淀粉、葡萄糖和果糖,成熟時淀粉酶活性降至最低,淀粉水解快速積累蔗糖。在整個發(fā)育過程中AI活性維持最高,完熟時降低,SPS在果實發(fā)育中期略有降低,完熟時升至最高,SS和NI一直很低且較穩(wěn)定。相關(guān)性分析表明淀粉含量與酸性轉(zhuǎn)化酶活性呈顯著正相關(guān);蔗糖、葡萄糖含量與SPS、SS呈極顯著正相關(guān);果糖含量與SS、AI均呈極顯著正相關(guān)。[結(jié)論]芒果成熟時淀粉分解、AI活性降低和SPS、SS活性的增加是引起‘KRS’果實蔗糖積累的主要因子。
芒果;糖積累;轉(zhuǎn)化酶;蔗糖合成酶;蔗糖磷酸合成酶
芒果是著名的熱帶果樹。我國是主要的芒果生產(chǎn)國之一,芒果廣泛分布在我國海南、云南、廣西、廣東、福建、四川和貴州等熱帶亞熱帶區(qū)域。從每年2月(海南三亞)至10月(四川攀枝花和云南華坪),我國芒果果品基本可實現(xiàn)周年供應(yīng),極大地滿足了市場需求,芒果產(chǎn)業(yè)發(fā)展優(yōu)勢和潛力十分明顯。
蔗糖代謝相關(guān)酶如酸性轉(zhuǎn)化酶(AI)、中性轉(zhuǎn)化酶(NI)、蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)對果實中糖的代謝和積累具有重要影響。有關(guān)糖積累與蔗糖代謝相關(guān)酶的關(guān)系在荔枝[1-2]、龍眼[3]、香蕉[4-5]、菠蘿[6]等熱帶果樹中已有報道。在國外,F(xiàn)uchs等[7-8]報道,芒果果實發(fā)育過程中單果重和淀粉酶活性增加,成熟時淀粉降解其淀粉含量降低,還原糖增加,在蔗糖的快速積累階段SPS活性增加了10倍,AI活性降低。中國熱帶農(nóng)業(yè)科學院南亞熱帶作物研究所成員已報道芒果成熟階段的糖代謝及相關(guān)酶活性[9-10],果實成熟時淀粉水解,蔗糖含量快速增加,SPS和SS酶活性升高,AI活性降低。他們收集的芒果種質(zhì)資源豐富,已超過200份。不同種質(zhì)資源果實所積累的糖的含量及比率是決定其品質(zhì)和商品價值的主要因素。前期研究中,人們根據(jù)果實成熟時單糖、雙糖比例將芒果種質(zhì)分為不同糖積累型,其中臺農(nóng)1號屬于蔗糖積累類型,‘KRS’屬于己糖積累型[11],同時對臺農(nóng)1號果實發(fā)育成熟過程中的糖分及相關(guān)代謝酶活性進行研究,結(jié)果表明芒果成熟時淀粉分解、AI活性降低,SPS和SS活性增加是引起果實蔗糖積累的主要因子[12]。‘KRS’芒果是20世紀90年代中國熱帶農(nóng)業(yè)科學院南亞熱帶作物研究所從澳大利亞引進的芒果品種,果實卵圓形,品質(zhì)優(yōu)良。但‘KRS’芒果實果實生長發(fā)育過程中糖積累及相關(guān)酶活性是否不同于臺農(nóng)1號糖積累尚未明確。鑒于此,中國熱帶農(nóng)業(yè)科學院南亞熱帶作物研究所成員以‘KRS’芒果品種為試材,研究了果實發(fā)育成熟過程中糖積累和代謝相關(guān)酶活性的關(guān)系,旨在為生產(chǎn)實踐中栽培措施的改良提供參考。
1 材料與方法
1.1材料試驗于2011年在中國熱帶農(nóng)業(yè)科學院南亞熱帶作物研究所芒果種質(zhì)資源圃進行,選用12年生的‘KRS’嫁接苗為材料,株距4 m,行距5 m,生長結(jié)果正常,常規(guī)田間管理。
1.2方法
1.2.1樣品采集。盛花時選擇樹冠大小、花期基本一致的健壯植株掛牌,從盛花后40 d開始,每隔10 d取樣1次,直到果實青熟采收,常溫放置成熟。每次隨機采10~15個果實,帶回實驗室,去除果皮后取果肉,混勻后用液氮速凍,置于-80 ℃超低溫冰箱中保存?zhèn)溆谩T囼炘O(shè)3次重復(fù)。
1.2.2測定指標與方法。葡萄糖、果糖、蔗糖的測定參照張秀梅等[6]的方法;蔗糖代謝相關(guān)酶活性的測定參照王惠聰?shù)萚2,6]的方法;淀粉酶活性的測定參照李雯等[13]的方法;淀粉含量的測定參照徐昌杰等[14]的方法。
2 結(jié)果與分析
2.1芒果果實發(fā)育期各組成糖分累積動態(tài)特征
2.1.1淀粉含量。由圖1可知,盛花后40 d,淀粉含量最低,僅17.90 mg/g,除果實發(fā)育100 d,淀粉含量略有降低,隨著果實的發(fā)育至120 d,淀粉含量一直呈上升趨勢,120 d淀粉含量最高,達419.60 mg/g FW,完熟時淀粉含量僅為5.60 mg/g FW,降至最低,說明隨著果實的發(fā)育進程,淀粉含量快速積累,果實完熟時淀粉分解轉(zhuǎn)化成糖。
2.1.2可溶性糖類含量。芒果可溶性糖包括葡萄糖、果糖和蔗糖,對于果實品質(zhì)起著重要作用。隨著果實發(fā)育成熟進程,蔗糖、葡萄糖和果糖表現(xiàn)出不同的變化規(guī)律(圖1)。‘KRS’果實發(fā)育至采收時,幾乎沒有蔗糖的積累,隨著果實的成熟,蔗糖快速積累,其含量達最高(64.45 mg/g FW)。在整個果實發(fā)育過程中,葡萄糖含量變化較平穩(wěn),在4.90~10.90 mg/g FW波動,僅在果實完熟時迅速升至最高,達21.60 mg/g FW。在果實發(fā)育前期(花后40~70 d),果糖含量變化平穩(wěn),果實發(fā)育70 d 至青熟時,果糖含量一直上升,完熟升至最高,達47.68 mg/g FW。可見,果實發(fā)育前期主要積累淀粉、葡萄糖和果糖,果實采后完熟階段,淀粉分解,蔗糖快速積累。

圖1 ‘KRS’果實發(fā)育過程中淀粉和可溶性糖含量的變化Fig.1 The change of starch and soluble sugar content in ‘KRS’ fruit development process
2.2芒果果實發(fā)育期糖代謝相關(guān)酶活性動態(tài)特征
2.2.1淀粉酶活性。淀粉酶活性在果實發(fā)育過程中,花后40~70 d 時淀粉酶活性快速升高,70 d 時達428.3 mg/g·s,70~90 d 時淀粉酶活性降低,90~120 d 時淀粉酶活性變化較平穩(wěn),完熟時淀粉酶活性快速降低,僅為150.8 mg/g·s(圖2)。
2.2.2蔗糖代謝相關(guān)酶活性。由圖3可知,在果實發(fā)育過程中,在果實發(fā)育40~60 d 時蔗糖磷酸合成酶活性快速升高,60 d 時達最大,為10.60 mg/g·h,花后60~120 d 時基本呈降低的趨勢,100 d 時酶活性最低,為5.66 mg/g·h,至完熟時升至最高,達19.00 mg/g·h。花后40~110 d 時蔗糖合成酶活性變化較平穩(wěn),在2.30~4.90 mg/g·h,完熟時升至最高,為11.90 mg/g·h。整個果實發(fā)育過程中,蔗糖磷酸合成酶活性高于蔗糖合成酶,說明參與蔗糖合成的酶以蔗糖磷酸合成酶為主。

圖2 ‘KRS’果實發(fā)育過程中淀粉酶活性的變化Fig.2 The change of amylase activity in ‘KRS’ fruit development process
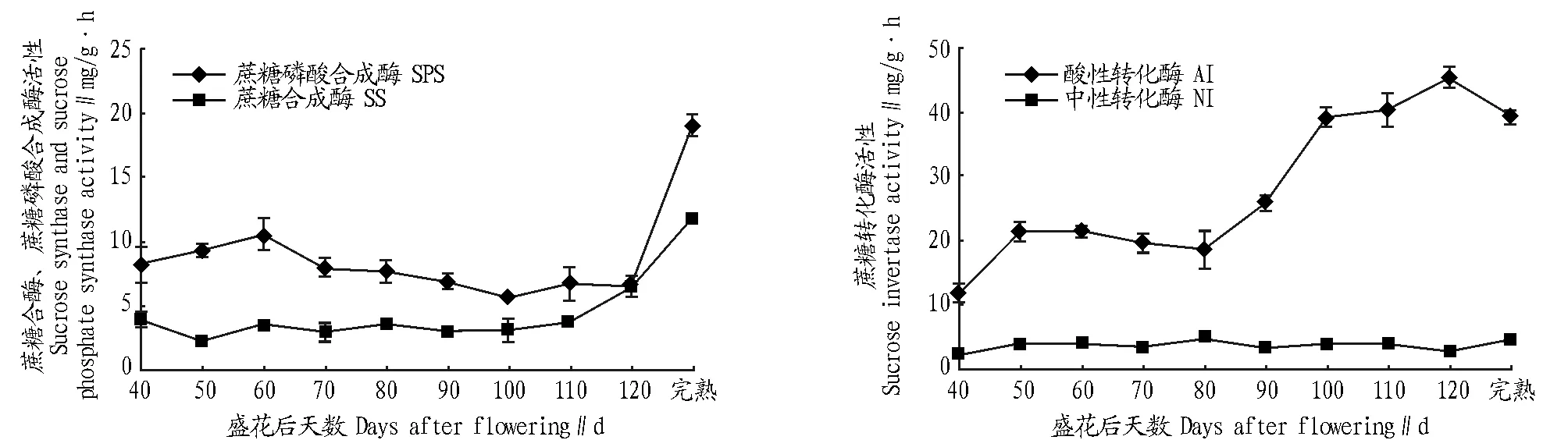
圖3 ‘KRS’果實發(fā)育中糖代謝相關(guān)酶活性的變化Fig.3 The change of metabolism related enzyme activity in ‘KRS’ fruit development process
在整個果實發(fā)育過程中,酸性轉(zhuǎn)化酶活性呈升高的趨勢,花后40~50 d 時快速升高,花后60~80 d 時變化較平穩(wěn),花后80~120 d 時快速升高,120 d 時活性達45.50 mg/g·h,果實完熟時降低至39.20 mg/g·h。在整個發(fā)育成熟過程中,中性轉(zhuǎn)化酶活性變化不大,維持在2.10~4.50 mg/g·h。總體來說,酸性轉(zhuǎn)化酶活性遠高于中性轉(zhuǎn)化酶,說明蔗糖分解酶類以酸性轉(zhuǎn)化酶為主。
2.2.3糖分與相關(guān)酶活性的關(guān)系。由表1可知,蔗糖含量與蔗糖磷酸合成酶和蔗糖合成酶活性均呈極顯著正相關(guān);葡萄糖含量與蔗糖磷酸合成酶和蔗糖合成酶活性均呈極顯著正相關(guān);果糖含量與蔗糖合成酶和酸性轉(zhuǎn)化酶活性均呈極顯著正相關(guān),淀粉含量與酸性轉(zhuǎn)化酶呈顯著正相關(guān)。

表1 芒果果實糖和酶活性的相關(guān)性
注:*表示P<0.05;**表示P<0.01。
Note:* stands forP<0.05;** stands forP<0.01.
3 討論
芒果屬于呼吸躍變型果實,其糖積累類型為淀粉轉(zhuǎn)化型[15],芒果果實發(fā)育S模式的快速生長階段以積累淀粉為主。果實接近青熟時,芒果淀粉酶活性增加,淀粉水解作用加強,使成熟時芒果果實蔗糖、葡萄糖和果糖含量增加。果實成熟時蔗糖快速增加,占總糖的60%~75%。芒果果實發(fā)育過程中單果質(zhì)量和淀粉酶活性增加,成熟時淀粉含量降低,還原糖和非還原糖增加[7]。Ito等[16]報道在果實發(fā)育過程中葉綠體中的淀粉含量增加,成熟時幾乎完全水解為糖,未成熟階段Alphonso 芒果淀粉含量為14%,成熟時僅為0.3%。成熟的Irwin芒果實幾乎未檢測到淀粉含量,而蔗糖含量顯著增加。Haden芒成熟過程中,淀粉降解、還原糖增加,在蔗糖的快速積累階段SPS活性增加了10倍,SS活性較穩(wěn)定,AI活性在完熟時降低[8],這與筆者所在研究所之前報道的粵西1號、Irwin芒[9-10]結(jié)果相似。該研究中,在‘KRS’芒果果實快速生長期,AI活性一直較高使果實發(fā)育過程中蔗糖積累很少,果糖和葡萄糖積累較多,果實快速積累淀粉,在采收青熟時淀粉含量達最高,成熟時淀粉水解,淀粉酶活性和淀粉含量降低轉(zhuǎn)化為蔗糖,使蔗糖快速積累,與蔗糖合成酶和蔗糖磷酸合成酶活性升高相一致。這與Keitt芒的研究結(jié)果相似[17-18]。該研究結(jié)果與臺農(nóng)1號芒果的糖積累規(guī)律相似,筆者推測不同糖積累類型的品種其糖的動態(tài)變化與相關(guān)酶活性相似。
高活性的轉(zhuǎn)化酶與果實的快速膨大相關(guān)[19],‘KRS’果實從幼果期至快速膨大期,較高的酸性轉(zhuǎn)化酶活性使果實中蔗糖的含量很低,蔗糖轉(zhuǎn)化酶活性增強,是由于該時期運輸?shù)焦麑嵵械恼崽侵饕糜诜纸猓a(chǎn)生能量以供果實膨大生長的需要。這與南瓜[20]、香瓜[21]和葡萄[22]等植物中的研究結(jié)果相似。轉(zhuǎn)化酶的活性下降是蔗糖積累的前提條件,在果實成熟期‘KRS’芒存在明顯的蔗糖積累轉(zhuǎn)折點,蔗糖積累的同時伴隨著蔗糖合成酶和蔗糖磷酸合成酶活性的升高及酸性轉(zhuǎn)化酶活性的下降。此時運輸?shù)焦麑嵵械恼崽潜毁A存起來,這與桃[23]、芒果[18]等果實上的試驗結(jié)果相似。
4 結(jié)論
芒果是淀粉轉(zhuǎn)化型果實,隨著果實的生長發(fā)育,‘KRS’芒果快速積累淀粉,青熟時達最高,完熟時顯著降低;在果實快速生長階段蔗糖磷酸合成酶活性略有降低,完熟時升至最高,蔗糖合成酶活性變化較小,果實成熟時快速升高;在果實發(fā)育過程中酸性轉(zhuǎn)化酶活性一直維持較高,完熟時顯著降低;中性轉(zhuǎn)化酶變化平穩(wěn)且一直較低。果實發(fā)育過程中酸性轉(zhuǎn)化酶對蔗糖的影響較大,蔗糖的積累很少,果糖和葡萄糖積累較多,完熟時淀粉酶活性降低,淀粉水解轉(zhuǎn)化成蔗糖,這與蔗糖合成酶和蔗糖磷酸合成酶活性升高一致。綜上所述,芒果成熟時淀粉分解、酸性轉(zhuǎn)化酶活性降低,蔗糖合成酶和蔗糖磷酸合成酶活性增加,是引起‘KRS’果實蔗糖積累的主要因子。
[1] 李建國,羅詩,袁煒群.荔枝果實成熟期間糖積累和糖代謝相關(guān)酶活性變化[J].華南農(nóng)業(yè)大學學報,2003,24(2):87-88.
[2] 王惠聰,黃輝白,黃旭明.荔枝果實的糖積累與相關(guān)酶活性[J].園藝學報,2003,30(1):1-5.
[3] 劉麗琴,李偉才,舒波,等.蔗糖代謝相關(guān)酶在‘石硤’龍眼假種皮糖積累中的作用[J].果樹學報,2015(4):653-657.
[4] HUBBARD N L,PHARR D M,HUBER S C.Role of sucrose phosphate synthase in sucrose biosynthesis in ripening bananas and its relationship to the respiratory climacteric[J].Plant physiol,1990,94:201-208.
[5] 袁揚靜,胡玉林,謝江輝.溫度對采后香蕉果實糖代謝及其酶活性的影響[J].熱帶作物學報,2011,32(1):66-70.
[6] 張秀梅,杜麗清,謝江輝,等.蔗糖代謝相關(guān)酶在卡因菠蘿果實糖積累中的作用[J].果樹學報,2006,23(5):707-710.
[7] FUCHS Y,PESIS E,ZAUBERMAN G.Changes in amylase activity,starch and sugars contents in mango fruit pulp[J].Scientia horticulturae,1980,13(2):155-160.
[8] CASTRILLO M,KRUGER N J,WHATLEY F R.Sucrose metabolism in mango fruit during ripening[J].Plant science,1992,84(1):45-51.
[9] 魏長賓,武紅霞,馬蔚紅,等.芒果成熟階段蔗糖代謝及其相關(guān)酶類研究[J].西南農(nóng)業(yè)學報,2008,21(4):972-974.
[10] 魏長賓,武紅霞,馬蔚紅,等.粵西1號芒果成熟階段的蔗糖代謝[J].熱帶作物學報,2009,30(6):735-739.
[11] 武紅霞,邢姍姍,王松標,等.芒果種質(zhì)資源糖積累的基因型差異分析[J].中國農(nóng)學通報,2011,27(10):266-270.
[12] 武紅霞,邢姍姍,王松標,等.‘臺農(nóng)1號’芒果果實發(fā)育過程中的糖分積累與相關(guān)酶活性研究[J].西北植物學報,2011,31(9):1811-1815.
[13] 李雯,邵遠志,陳維信.淀粉酶活性測定方法的改進[J].植物生理學通訊,2005,41(5):655-656.
[14] 徐昌杰,陳文峻,陳昆松,等.淀粉含量測定的一種簡便方法:碘顯色法[J].生物技術(shù),1998,8(2):41-43.
[15] 張上隆,陳昆松.果實品質(zhì)形成與調(diào)控的分子生理[M].北京:中國農(nóng)業(yè)出版社,2007:1-2.
[16] ITO T,SASAKI,K,YOSHIDA Y.Changes in respiration rate,saccharide and organic acid content during the development and ripening of mango fruit (MangiferaindicaL.cv.‘Irwin’)cultured in a plastic house[J].Journal of the Japanese society for horticultural science,1997,66:629-635.
[17] MEDLICOTT A P,THOMPSON A K.Analysis of sugar and organic acids in ripening mango fruit(MangiferaindicaL.cv.‘Keitt’)by high performance liquid chromatography[J].Journal of the science of food and agriculture,1985,36:561-566.
[18] CORDENUNSI B R.Starch mobilization and sucrose accumulation in the pulp ofKeittmangoesduring postharvest ripening[J].Journal of food biochemistry,2008:32(3):384-395.
[19] KRISHNAN H B,PUEPPKE S G.Cherry fruit invertase:Partial purification,characterization and activity during fruit development[J].Journal of plant physiology,1990,135(6):662-666.
[20] 孫守如,楊子琴,張菊平,等.南瓜果實發(fā)育過程中糖代謝及相關(guān)酶活性的變化[J].西北農(nóng)林科技大學學報(自然科學版),2008,36(6):159-164.
[21] SCHAFFER A A,ALINI B,F(xiàn)OGELMAN E.Sucrose metabolism and accumulation in developing fruit ofCucumis[J].Phytochem,1987,26:1883-1887.
[22] 閆梅玲,王振平,范永,等.蔗糖代謝相關(guān)酶在赤霞珠葡萄果實糖積累中的作用[J].果樹學報,2010(5):703-707.
[23] MORIGUCHI T,SANADA T,YAMAKI S.Seasonal fluctuation of some enzymes relating to sucrose and sorbitol metabolism in peach fruit[J].Amer Soc Hort Sci,1990,115:278-281.
The Relationship between Sugar Accumulation and Related Enzyme Activities during the Development Process of ‘KRS’Mango Fruits
WU Hong-xia, YAO Quan-sheng, WANG Song-biao et al
(Key Laboratory of Tropical Fruit Biology of Ministry of Agriculture, South Subtropical Crops Research Institute, CATAS, Zhanjiang,Guangdong 524091)
[Objective] The aim was to discuss physiological mechanism of sugar accumulation and transformation in mango fruit. [Method]The relationship between the contents of starch, sucrose, glucose and fructose, and the activities of amylase, sucrose metabolizing enzymes including acid invertase (AI), neutral invertase (NI), sucrose synthase(SS) and sucrose phosphate synthase(SPS) during fruit development process in KRS mango were studied. [Result] The results showed that starch, fructose and glucose accumulated at the early stage of fruit development, amylase activity dropped to minimum along with the starch decomposition and sucrose accumulated at the ripening stage. The activities of AI maintained higher throughout the developmental stage, dropped slightly at the ripening stage. The SPS activities dropped slightly in middle stage of fruit development, then raised to the highest at the ripening stage. The activities of SS and NI maintained minimum and changed very little throughout the development process. The starch content had positive correlation with amylase. The contents of sucrose and glucose had significantly positive correlation with the activities of SPS and SS. The content of fructose had significantly positive correlation with SS and AI. [Conclusion] Decomposition in starch, reduction in AI, increment in SS and SPS activities are responsible for sucrose accumulation in developing fruits of mango ‘KRS’.
Mango; Sugar accumulation; Invertase; Sucrose synthetase; Sucrose phosphate synthase
農(nóng)業(yè)行業(yè)科研專項子項目(201203092-3);廣東省自然科學基金粵東西北創(chuàng)新人才聯(lián)合培養(yǎng)項目(2016A030307006);科研院所基本科研業(yè)務(wù)費專項(1630062015001,1630062015007)。
武紅霞(1975- ),女,山西萬榮人,副研究員,博士,從事果樹品質(zhì)生理與栽培技術(shù)研究。
2016-04-29
S 601
A
0517-6611(2016)17-024-03
