茅山丘陵地區茶園秋季雜草種群生態位研究
2016-08-09 10:15:05王虎琴孫國俊王哲明李粉華陳樂季忠鐘霽白潔瑞
蠶桑茶葉通訊 2016年3期
王虎琴 孫國俊 王哲明 李粉華 陳樂 季忠 鐘霽 白潔瑞
(1.常州市金壇區茶業技術指導站 213200;2.常州市金壇區植保植檢站 213200;3.揚州大學園藝與植物保護學院 225009;4.金壇市茅麓茶廠 213254;5.常州市金壇區農業廣播學校 213200; 6.常州市金壇區種子管理站 213200;7.常州市金壇區農業試驗站 213200;8.常州市金壇區土壤肥料技術指導站 213200)
?
茅山丘陵地區茶園秋季雜草種群生態位研究
王虎琴1孫國俊2、3王哲明4李粉華2陳樂5季忠6鐘霽7白潔瑞8
(1.常州市金壇區茶業技術指導站213200;2.常州市金壇區植保植檢站213200;3.揚州大學園藝與植物保護學院225009;4.金壇市茅麓茶廠213254;5.常州市金壇區農業廣播學校213200; 6.常州市金壇區種子管理站213200;7.常州市金壇區農業試驗站213200;8.常州市金壇區土壤肥料技術指導站213200)
摘要:對茅山丘陵茶園秋季雜草的發生種類、數量、高度等進行調查,將其轉換成相對多度和重要值數據,在此基礎上計測了茅山丘陵茶園22種主要雜草的生態位寬度和生態位重疊值。結果表明:茅山丘陵茶園秋季雜草共發現80種,隸屬36個科、74個屬。其中,發生頻率>20%的雜草有22種。22 種雜草中,馬唐、小飛蓬、小旱稗、牛筋草等4種雜草實際生態位較寬,為茅山丘陵地區茶園秋季優勢雜草;馬唐、小旱稗小飛蓬、牛筋草、酢漿草、泥胡菜、繁縷、鐵莧菜、族生卷耳、苦苣等雜草生態位重疊值較大,說明它們對資源的競爭較為激烈。
關鍵詞:茶園;秋季雜草; 生態位寬度;生態位重疊值
生態位是指在自然生態系統中生物種群在時間、空間上的位置及其與相關種群之間的功能關系,反映了物種對環境資源的需求[1~2]。近年來生態位理論已廣泛應用于多種生物的研究。
雜草是農田生態系統的生物組分,維持適當數量的雜草對保護農田生物多樣性發揮著重要作用[3~4],但雜草往往與作物競爭水、肥、光等自然資源,影響作物生長與產量[5]。生態位寬度可以作為雜草對農田環境資源利用多樣性的一種測度,反映了不同雜草在農田中的生態適應幅度。生態位重疊可以作為植物種間生態學相似性的測度[6],通過計測不同雜草間的實際生態位重疊值,能明確不同雜草對生態條件要求的相似性程度,可以預測雜草種群競爭結局和人為干擾下群落演替的方向[2、7]。故研究雜草生態位對預測雜草群落演替,特別是預測除草劑長期單一使用后農田雜草群落的演替有重要的指導意義[8]。雖然目前國內外對各類植物群落生態位的研究相當普遍[9],但對茶園雜草的生態位進行研究預測種群動態變化以揭示茶園雜草的生態特性的報道較少。筆者研究了茅山丘陵地區茶園秋季雜草生態位寬度和生態位重疊值,以期明確茶園秋季雜草對資源環境的生態適應性和雜草之間的生態相似性,為該區域茶園秋季雜草的綜合治理提供理論依據。
1研究地區與方法
1.1研究區概況
研究區金壇位于31°33′42″~31°53′22″N, 119°17′45″~119°44′59″E,地處長江下游江蘇省南部。茶葉生產面積2733hm2,主要分布在茅山丘陵地帶,海拔10~50m,土質以黃棕壤土、基性巖土、石灰巖土為主。常年年均氣溫15.3 ℃,年均降雨量1063.6mm,雨水多集中于春夏秋季。常年年均日照時數為2035.5h[10]。
1.2雜草生物多樣性調查
筆者于2013年的10月,在夏秋季多數雜草已開花結實時,根據茶葉生產布局,分別選擇不同地點、茶葉品種、種植時間、土質、土壤類型、坡度、農事管理措施等有代表性的茶園類型,每種類型各調查3~4塊茶園,共調查30塊田。每塊茶園采用“M”形9點取樣法進行調查[11~12],考慮到茶樹覆蓋度大,空行較小的特點,每個樣方0.25m2(0.5m×0.5m),分別記載雜草種類、數量、高度。并詳細記載調查地點、耕作方式、 土壤類型、土壤水分狀況、酸堿度、除草劑使用等情況。
1.3數據分析
按下列公式計算雜草生態重要值:
重要值(IV)=(RD%+RF%+RH%)/3;相對多度:RA%=RU%+RD%+RF%,其中RU、RD、RF、RH分別為某雜草相對均度、相對密度、相對頻度、相對高度。田間均度:某雜草在田塊中出現樣方數占調查田塊總樣方數百分比;田間頻度:某雜草出現的田塊數占總調查田塊數的百分比;田間密度:某雜草在各調查田塊平均密度和與田塊數之比;平均高度:樣方中某雜草的高度平均值;相對均度:某雜草均度與各種雜草均度和之比;相對密度:某雜草平均密度與各種雜草密度之和相比;相對頻度:某雜草頻度與各種雜草頻度之和相比;相對高度:某雜草的平均高度與所有種的平均高度之和相比。相對多度較大的雜草將被視為當地的主要優勢雜草,體現雜草豐富程度[13~14]。
運用Excel 2003數據處理系統進行田間均度、頻度、密度、高度、重要值、相對多度等參數的處理分析,并以重要值為指標,建立原始數據矩陣,生態位寬度采用Levins計算方法,生態位重疊值采用Schoener計算方法。
Levins[15]生態位寬度:
式中Bi為物種i的Levins生態位寬度,Pij為物種i對第j資源水平的利用占對全部資源利用的百分率,Pij=nij/Ni,而Ni=∑nij,nij為物種i在資源水平j的數量特征值(如蓋度、重要值、密度等),本文中為物種i在第j個樣方內的重要值。r為資源水平總數,即樣方數。
Schoener[16]生態位重疊值:
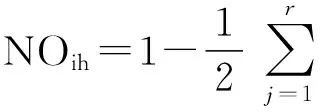
式中NOih為種i與種h生態位重疊值,r為資源等級數,Pij和Phj分別為種i與種h在第j個資源單位中所占的比例[17]。
2結果與分析
2.1茶園秋季雜草物種組成分析
調查發現,金壇茶園秋季雜草區系豐富,常見雜草有80種,分別隸屬36個科、74個屬。菊科、禾本科雜草為金壇茶園秋季雜草優勢種群,分別有14個屬、14種和8個屬、8種雜草,分別占總屬數、總種數的18.92%、17.50%和10.96%、10.00%;大戟科雜草有3屬5種,分別占總屬數、總種數的4.11%、6.25%;唇形科雜草有4屬4種,分別占總屬數、總種數的5.48%、5.00%;莧科雜草分別有3屬4種,分別占總屬數、總種數的4.11%、5.00%;豆科、百合科雜草有3屬3種,分別占總屬數、總種數的4.11%、3.75%;茄科雜草有2屬3種,分別占總屬數、總種數的2.74%、3.75%;莎草科雜草有1屬3種,分別占總屬數、總種數的1.37%、3.75%;蓼科、茜草科、薔薇科、傘形科、玄參科、石竹科雜草分別有2屬2種,分別占總屬數、總種數的2.74%、2.50%;堇菜科、車前科、酢漿草科、番杏科、防己科、風尾蕨科、海金沙科、葫蘆科、夾竹桃科、爵床科、鱗毛蕨科、蘿藦科、馬齒莧科、牻牛兒苗科、毛茛科、葡萄科、商陸科、、梧桐科、十字花科、旋花科、鴨跖草科、紫草科雜草等21科雜草各有1屬1種。其中,發生頻率大于20%的雜草有22種(表1),22種主要雜草中禾本科和菊科最多,分別是4種,5種,大戟科3種,石竹科2種,其它各科均為1種。
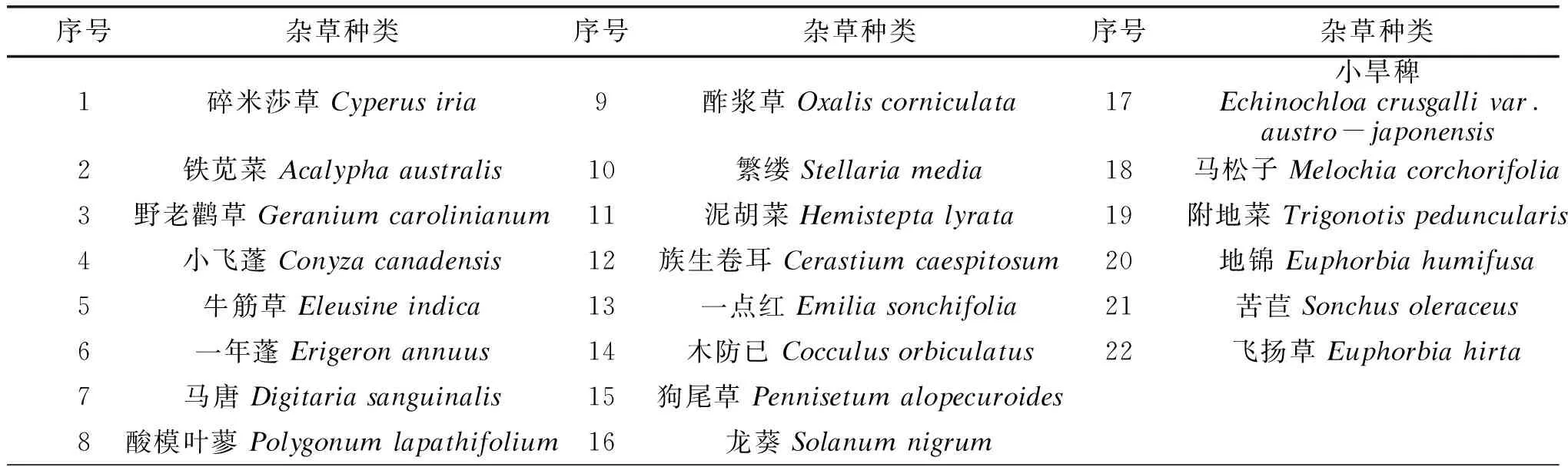
表1 金壇秋季茶園22種主要雜草名錄
2.2茶園秋季雜草生態位寬度分析
生態位寬度是一個生物所利用的各種資源的總和,是度量植物種群對環境資源利用狀況的尺度,其大小體現了物種在群落中的競爭地位, 物種生態位寬度越大, 則它對環境的適應能力越強[18]。茅山丘陵地區茶園秋季雜草中(表2)生態位寬度最寬的分別是馬唐(Digitaria sanguinalis)、小飛蓬(Conyza canadensis)、小旱稗(Echinochloa crusgalli var. austro-japonensis)、牛筋草(Eleusine indica)等,它們對環境資源利用的多樣性程度較高,是茅山丘陵地區秋季茶園最為重要的雜草;野老鸛草(Geranium carolinianum)、酢漿草(Oxalis corniculata)、狗尾草(Pennisetum alopecuroides)等雜草生態位寬度次之,是茅山丘陵地區秋季茶園的常見雜草。
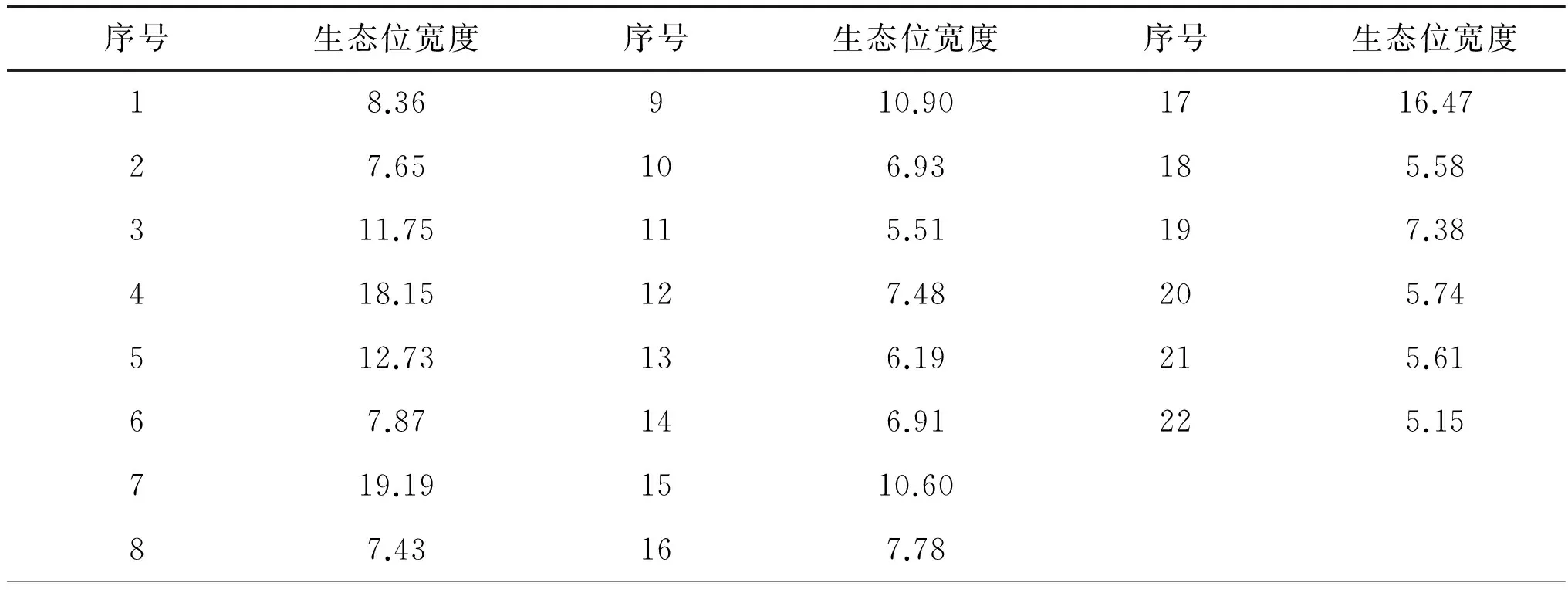
表2 金壇茶園秋季22種主要雜草生態位寬度
注:雜草物種編號1~22同表1。下同。
2.3茶園秋季雜草生態位重疊值
22種茶園秋季主要雜草對田間資源的利用是有差異的,主要表現為不同的生態位重疊值(表3)。在231對生態位重疊值中生態位重疊值在0.6~0.7的有1對,分別為馬唐-小旱稗;生態位重疊值在0.5~0.6的有7對,分別為馬唐-小飛蓬、馬唐-牛筋草、酢漿草-泥胡菜、繁縷-鐵莧菜、小旱稗-小飛蓬、小旱稗-牛筋草、族生卷耳-苦苣;生態位重疊值在0.4~0.5的有20對,占總數的8.66%;生態位重疊值在0.3~0.4 的有45對,占總數的19.48%;其余的158對的生態位重疊值較小,在0~0.3之間,其中有4對生態位重疊值為0。由此可知,多數雜草種對間存在較小的生態位重疊,只有少數種對間生態位重疊值較大,這些種對間利用資源的生態學特性比較相近。而生態位重疊值為0或近似為0的種對,多為生態學習性差異較大的種對。

表3 金壇茶園秋季22種主要雜草生態位重疊值
3討論
生態位是物種的屬性特征表現,它定量地反映物種與生境的相互作用關系[19]。在自然狀況下,生態位接近的兩個種,很少能長期穩定地共存,這涉及到生態位重疊和資源分享的數量問題。從資源利用的角度來看,生態位重疊較大的種間,要么對資源的利用有較大的相似性,要么在資源的利用上有著互補的需求[20]。從競爭的角度看,在資源相對不足的情況下,生態位重疊較大的種間可能會產生較為激烈的利用性競爭[21]。
研究結果表明,茅山丘陵地區秋季茶園中的馬唐、小飛蓬、小旱稗等雜草的生態位寬度值較大,說明它們對環境資源利用的多樣性比較高,是該地區秋季茶園中生態適應性強、發生危害相對較重的雜草種類,應該成為秋季茶園雜草防除的主要對象。馬唐、小飛蓬、小旱稗等雜草有較強的生態適應性和耐貧瘠特性。研究發現,馬唐-小旱稗,馬唐-小飛蓬、牛筋草、酢漿草、泥胡菜,繁縷-鐵莧菜,小旱稗-小飛蓬、牛筋草,族生卷耳-苦苣等雜草間的生態位重疊值較大,對資源的競爭較為激烈,在防除實踐中如果僅防除其中的一類雜草,這些生態位相近的雜草會替代成為優勢雜草。因此,制訂防除措施時需要考慮由它們構成的雜草群落。應用 DCA排序結合生態位特征分析的方法,可以更全面地研究各種群之間資源利用和空間分布的相互關系。由 DCA排序圖可以看出,馬唐、小飛蓬、小旱稗等幾種雜草組成的群落,對該地區的生態環境適應性較強,常作為茶園秋季生境中的優勢種,因此雜草防除應選擇禾闊雙除除草劑或復配劑。單一除草劑的施用易導致雜草群落演替為多年生難防除的雜草[22]。不同的雜草控制措施(如化感生物控制、耕作方式、人工及化學除草措施等)通過形成特殊的環境生態條件,常常限制了某些雜草的生長,同時又適合另外一些雜草的生長,對田間雜草群落的物種組成進行著篩選,起著過濾雜草的作用[23]。茶樹種群生態元的生態位擴充可認為是對各類雜草種群生態位的入侵,可能導致某些雜草種群在資源競爭中處于劣勢,從而使之釋放出其所占有的資源空間。在茶園生態系統中,雜草與茶樹競爭光照、水分和養分時,雜草處于劣勢,有機物積累相對較少。實踐表明茶園封行,可抑制茶園中雜草數量的發生,促進作物良好生長。另外,農業生態系統被人為控制,存在大量潛在生態位[24],可采取一些措施,使生態元的潛在生態位變為實際生態位,以降低理想生態位離差。例如隨著施氮的增加,茶樹生長旺盛,可有效地截斷雜草的光資源,減少雜草的光合作用,從而導致雜草競爭力下降,茶樹在競爭中逐漸占據主要地位,雜草長勢減弱。
由于雜草生活史短,種群之間的競爭結局容易在短時間內反映出來,故應用生態位理論來研究特定區域茶園秋季雜草之間的競爭關系, 除可以闡明生態位研究中的一些理論問題,也能明確不同雜草對生態條件要求的相似程度,同時,還可以了解各種雜草對所用除草劑的敏感性,為茶園秋季雜草的綜合防治提供更為明確的理論依據。
參考文獻
[1]付必謙. 生態學實驗原理與方法(第一版)[M]. 北京:科學出版社,2006:1~327.
[2]吳海榮,強勝,林金成. 南京市春季外來雜草調查及生態位研究[J]. 西北植物學報,2004,24(11):2061~2068.
[3]Fenn M E, Poth M A, Aber J D, et al. Nitrogen excess in north American ecosystems: Predisposing factors, ecosystem responses, and management strategies[J] .Ecological Applications, 1998, 8(3): 706~733.
[4]Anderson R L, Tanaka D L, Black A L, et al. Weed community and species response to crop rotation, tillage, and nitrogen fertility[J]. Weed Technology, 1998, 12(3): 531~536.
[5]李儒海,強勝,邱多生,等. 長期不同施肥方式對稻油輪作制水稻田雜草群落的影響[J]. 生態學報,2008,28(7):3236~3243.
[6]王剛. 植物群落中生態位的計測[J]. 植物生態學與地植物學叢刊,1984,8(4)∶330~335.
[7]郭水良,李揚漢. 金華地區秋旱作物田雜草生態相似關系研究[J]. 武漢植物學研究,1998,16(1):39~46.
[8]郭水良, 李揚漢, 趙鐵橋. 浙江金華地區雜草群落中雜草生態位的研究[J]. 植物生態學報, 1998, 22( 1 ): 76~84.
[9]王正文, 祝廷成. 松嫩草原主要草本植物的生態位關系及其對水淹干擾的響應[J]. 草業學報, 2004, 13( 3): 27~34.
[10]金壇年鑒編纂委員會. 金壇年鑒(2000~2002)[M]. 北京: 中央文獻出版社, 2003.
[11]Thomas A C. Weed survey system used in saskatchewan for cereal and oilseed crops[J]. Weed Science, 1985, 33: 34~43.
[12]張朝賢,胡祥恩,錢益新,等. 江漢平原麥田雜草調查[J]. 植物保護,1998,24(3):14~16.
[13]強勝. 雜草學[M]. 北京: 中國農業出版社, 2001.
[14]王英姿, 紀明山, 祁之秋, 等. 遼寧省果園雜草發生情況調查[J]. 植物保護, 2008, 34(4):98~101.
[15]Levins R. Evolution in changing environments: some theoretical explorations[M]. Princeton:Princeton University Press. 1968.
[16]Schoener T. Resource partitioning in ecological communities[J]. Science, 1974, 185: 27~39.
[17]劉金福,洪偉. 格氏栲群落生態學研究—格氏栲林主要種群生態位的研究[J]. 生態學報,1999,19(3):347~352.
[18]史紅文,江明喜,胡理樂. 瀕危植物毛柄小勾兒茶的生態位研究[J]. 武漢植物學研究,2007,25(2):163~168.
[19]張光明,謝壽昌. 生態位概念演變與展望[J]. 生態學雜志,1997,16(6):46~51.
[20]史作民,程瑞梅,劉世榮. 寶天曼落葉闊葉林種群生態位特征[J]. 應用生態學報,1999,10(3):265~269.
[21]向悟生,李先琨,蘇宗明,等. 元寶山冷杉群落主要樹木種群生態位的初步研究[J]. 武漢植物學研究,2002,20(2):105~112.
[22]余柳青,江榮昌,高子瑜,等. 浙江省稻田雜草群落及其演替[J]. 雜草科學,1993(4):21~23.
[23]魏守輝,強勝,馬波,等. 稻鴨共作及其他控草措施對稻田雜草群落的影響[J]. 應用生態學報,2005,16(6):1067~1071.
[24]章熙谷. 農業生態系統中新物種引進的初步探討[J]. 生態學雜志,1990,9(2):36~39.
資助項目:國家公益性行業(農業)科研專項(200903004)子課題:“茶樹有害生物種類與發生危害特點研究(200903004-43)、江蘇省農作物有害生物種類與發生危害特點研究”。
