馬鈴薯內生細菌的分離鑒定及種群多樣性分析
2016-06-15 03:49:28屈青松彭萬里翟立明林榕姍山東農業(yè)大學生命科學學院山東農業(yè)微生物菌種資源保藏中心山東泰安271018
中國蔬菜 2016年6期
屈青松 彭萬里 翟立明 劉 曉 林榕姍(山東農業(yè)大學生命科學學院,山東農業(yè)微生物菌種資源保藏中心,山東泰安 271018)
?
馬鈴薯內生細菌的分離鑒定及種群多樣性分析
屈青松 彭萬里 翟立明 劉 曉 林榕姍*
(山東農業(yè)大學生命科學學院,山東農業(yè)微生物菌種資源保藏中心,山東泰安 271018)
摘 要:為了分離鑒定馬鈴薯內生細菌,對相同種植地點的3個不同品種馬鈴薯內生細菌的群落結構進行分析;從不同品種馬鈴薯中分離純化菌株,進行基因序列分析和生理生化鑒定;取相同部位馬鈴薯組織進行PCR-DGGE操作,并對DGGE圖譜上的條帶進行差異性分析。最終分離得到9株內生細菌,鑒定1-1、1-2為枯草芽孢桿菌(Bacillus subtilis sp.),1-3為類芽孢桿菌(Paenibacillus sp.),1-4為韓國假單胞菌(Pseudomonas koreensis sp.),1-5、1-6、2-3為解淀粉芽孢桿菌(Bacillus amyloliquefaciens sp.),2-2為蒼白桿菌(Ochrobactrum sp.),3-6為副球菌(Paracoccus sp.)。DGGE圖譜表明不同品種的馬鈴薯內生細菌既有差異性又有相似性。最終結果表明3個品種馬鈴薯內生細菌具有較豐富的種群多樣性,且差異性明顯,品種可能是影響馬鈴薯內生細菌群落結構的重要因素。
關鍵詞:馬鈴薯;DGGE;內生細菌;鑒定
屈青松,男,本科生,專業(yè)方向:生物科學,E-mail:quqingsong@ outlook.com
植物內生菌是指在植物生活史的一定階段或全部階段存在于健康植物內部組織的微生物類群,而被感染的寄主植物不會表現(xiàn)明顯感染癥狀(Herre et al.,2007;陳雪英 等,2008;李瑞 等,2009)。內生菌要從嚴格消毒的組織和汁液中分離,或從植物組織內直接擴增微生物DNA來證明其存在(Stone et al,2000;石晶盈 等,2006)。內生細菌廣泛存在于植物體內,分布于植物的葉、莖、花、果實、種子等器官、組織或細胞間隙之中。部分內生細菌的次生代謝物中有時含有與宿主植物相同或相似的活性成分(Fisher et al.,1993;何勁 等,2006)。
由于內生細菌長期生長于寄主內部,并與之協(xié)同進化,故具有促進植物生長、抵抗病蟲害的能力,部分內生細菌還有增加寄主對環(huán)境脅迫的抗性作用(Munif et al.,2001)。研究表明馬鈴薯的塊莖中具有豐富的內生菌資源(文才藝 等,2004),因此研究馬鈴薯內生細菌的分類及其生物多樣性對后續(xù)的研究有很大的指導意義。
一些不可培養(yǎng)的微生物可通過擴增出其DNA分子的手段證明其存在,變性梯度凝膠電泳(DGGE)就是一種常用的非培養(yǎng)手段研究微生物種群的方法,根據(jù)DNA在不同濃度變性劑中變性行為的不同而導致電泳遷移率發(fā)生變化的原理,將片段大小相同而堿基組成不同的DNA片段分開,從而對微生物群落進行分析(張一 等,2012)。根據(jù)DGGE圖譜中條帶的數(shù)量、亮度和位置可反映出樣品中微生物群落的部分信息,從而可用來對環(huán)境樣品中的微生物多樣性進行定性、半定性分析(劉敏 等,2000)。
前人關于馬鈴薯內生細菌的篩選多是建立在富集、篩選的傳統(tǒng)方法上,這樣會忽略掉馬鈴薯內生細菌中的大部分不可培養(yǎng)的菌株(Stenberg,1999),而利用分子生物學的方法可以克服傳統(tǒng)方法的缺點,更快速、更準確地檢測內生細菌的種群數(shù)目。
本試驗以泰山1號、中薯5號、荷蘭15號3個馬鈴薯品種為研究對象,從其塊莖中篩選內生細菌,并鑒定其所屬種群;同時對這3個馬鈴薯品種的相同位置塊莖進行了PCR-DGGE分析,分析塊莖中內生細菌的生物多樣性。
1 材料與方法
1.1 試驗材料
供試馬鈴薯品種分別為:泰山1號(編號1,泰安市農業(yè)科學研究院作物研究所選育)、中薯5號(編號2,中國農業(yè)科學院蔬菜花卉研究所選育)、荷蘭15號(編號3,荷蘭費烏瑞它系列品種),均由泰安市農業(yè)科學研究院經濟作物研究所種植,于2015年1月采集,長勢良好。所用引物參考林榕姍(2012)設計(表1),由南京金斯瑞生物科技有限公司合成。

表1 擴增引物
1.2 試驗方法
試驗于2015年1~9月在山東農業(yè)大學生命科學院,山東農業(yè)微生物菌種資源保藏中心進行。
1.2.1 馬鈴薯內生細菌的分離與純化 參照Ying等(2011)和Kumar等(2013)的方法,分別對3個品種的馬鈴薯進行表面消毒,用干凈的打孔器對馬鈴薯塊莖進行打孔,取距離馬鈴薯表皮0.5~1.0 cm處塊莖進行研磨,將研磨液用無菌水稀釋至1×10-2、1×10-3、1×10-43個梯度,稀釋后的懸液分別取100 μL涂布于869培養(yǎng)基(蛋白胨10 g、酵母膏10 g、NaCl 5 g、葡萄糖1 g、CaCl20.261 g、瓊脂20 g、水1 L、pH 7.0。)、LB培養(yǎng)基,每個梯度懸液涂布3個平行,將培養(yǎng)皿置于28 ℃培養(yǎng)箱培養(yǎng)3~5 d。挑取單菌落于培養(yǎng)基上進行純化,觀察純化后的菌落形態(tài);并參照《常見細菌系統(tǒng)鑒定手冊》方法進行革蘭氏染色,選取不同種的菌落再次純化,并制作斜面及甘油管保藏。
1.2.2 馬鈴薯內生細菌的鑒定 ① 16S rRNA 序列分析:采用細菌基因組DNA提取試劑盒(OMEGA)提取分離、純化后的菌株DNA。以所提取DNA為模板,使用P1/P2引物(表1)進行PCR擴增。PCR反應體系(50 μL):10×buffer 5 μL,MgCl23 μL,dNTP 4 μL,引物各1 μL,Taq DNA聚合酶(TAKARA)0.2 μL,DNA模板2 μL,用ddH2O補足50 μL。PCR反應條件:95 ℃預變性5 min;94 ℃變性1 min,58 ℃退火30 s,72 ℃延伸1 min30 s,35個循環(huán);72 ℃延伸10 min。PCR產物用1%的瓊脂糖凝膠電泳檢測后,送上海生工生物工程有限公司進行測序。根據(jù)測序結果,將擴增得到的序列在GenBank中進行BLAST分析,利用MEGA 6.06軟件進行ClustalW分析,并以Neighbor-joining方法構建系統(tǒng)發(fā)育樹,用Bootstrat (1 000次重復)進行檢驗。② 菌株生理生化特征、菌落特征和細胞形態(tài)測定:參照《伯杰細菌鑒定手冊》第8版分類系統(tǒng)及《常見細菌系統(tǒng)鑒定手冊》方法對所純化菌株的生理生化特征、形態(tài)特征等指標進行測定。
1.2.3 馬鈴薯塊莖總DNA的提取 取3個品種馬鈴薯相同位置塊莖,方法同1.2.1,利用E.Z.N.A.Soil DNA Kit和E.Z.A.N. Cycle-Pure Kit(OMEGA)提取并純化馬鈴薯塊莖總DNA。
1.2.4 PCR-DGGE分析 ① PCR擴增及定量:以提取所得馬鈴薯塊莖總DNA 為模板,使用 GC-357F/517R 引物(表1)進行PCR擴增。PCR反應體系(50 μL):10×buffer 5 μL,MgCl23 μL,dNTP 4 μL,DNA模板1 μL,引物各2 μL,Taq DNA聚合酶0.2 μL,用ddH2O補足50 μL。PCR反應條件:95 ℃預變性5 min;94 ℃變性1 min,57 ℃退火1 min,72 ℃延伸30 s,24個循環(huán);72℃延伸10 min。PCR產物使用E.Z.A.N.Cycle-Pure Kit進行純化,純化后使用1%的瓊脂糖凝膠電泳檢測,-20 ℃保存?zhèn)溆谩"?DGGE電泳:嚴格按照Bio-Rad的DGGE說明手冊步驟操作。
2 結果與分析
2.1 馬鈴薯內生細菌的篩選
從3個不同品種馬鈴薯塊莖中共分離得到18株內生菌,其中6株菌篩選自泰山1號,分別編號1-1、1-2、1-3、1-4、1-5、1-6;6株菌篩選自中薯5號,分別編號2-1、2-2、2-3、2-4、2-5、2-6;6株菌篩選自荷蘭15號,分別編號3-1、3-2、3-3、3-4、3-5、3-6。經形態(tài)學的觀察排除重復菌株和放線菌,共得到9株內生細菌分別為1-1、1-2、1-3、1-4、1-5、1-6、2-2、2-3、3-6,通過革蘭氏染色結果可知(圖1),1-1、1-2、1-5、1-6、2-3為革蘭氏陽性菌,1-3、1-4、2-2、3-6為革蘭氏陰性菌。
2.2 馬鈴薯內生細菌菌株的鑒定
2.2.1 16S rRNA序列分析結果 對9株馬鈴薯內生細菌所提取的DNA進行1%瓊脂糖凝膠電泳檢測,各條帶均較清晰,且無特異性條帶,說明這9株菌株的基因組DNA濃度較高,純度較好。以所提取的9株菌株基因組DNA為模板進行PCR擴增,并將擴增后的產物進行測序,分別得到這9株菌株的16S rRNA片段,DNA長度均在1 300~1 500 bp之間,將9株菌株的序列上傳至GenBank,獲得GenBank登錄號分別如下,1-1:KT781674、1-2:KT831431、1-3:KT831432、1-4:KU041145、1-5:KT831433、1-6:KT831434、2-2:KT831435、2-3:KT831436、3-6:KT831437。
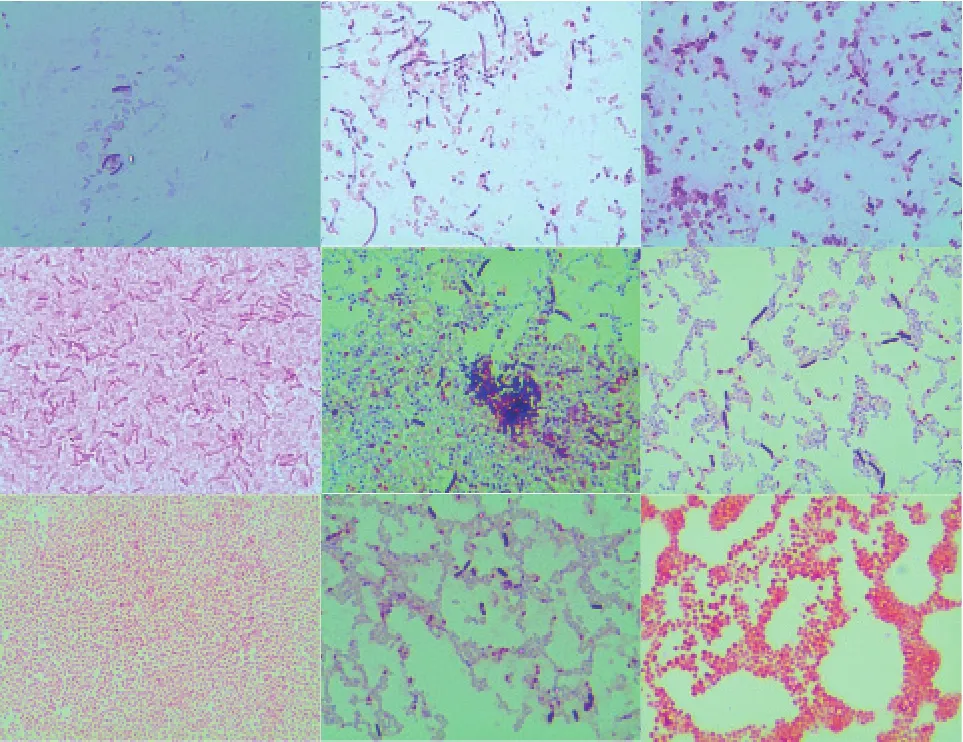
圖1 9株馬鈴薯內生細菌的革蘭氏染色顯微圖(光學顯微鏡,100倍×10倍)彩色圖片參見《中國蔬菜》網站:www.cnveg.org。
對這9株菌株的16S rRNA 進行Blast分析,均在GenBank中找到相似性在95%以上模式菌株,下載這些相似性較高的序列,用MEGA 6.06軟件進行多重比對,并構建了系統(tǒng)發(fā)育樹(圖2)。依據(jù)系統(tǒng)發(fā)育樹可知,1-1、1-2、1-5、1-6、2-3之間的相似性較高,且與Bacillus subtilis sp.和Bacillus amyloliquefaciens sp.親緣關系較近;1-3與Paenibacillus sp.親緣關系較近;1-4 與Pseudomonas koreensis sp.親緣關系較近;2-2 與Ochrobactrum ciceri sp.親緣關系較近;3-6與Paracoccus chinensis sp.親緣關系較近。
2.2.2 生理生化特征鑒定結果 根據(jù)生理生化鑒定與形態(tài)觀察(表2),結合16S rRNA 基因鑒定結果,鑒定1-1、1-2為枯草芽孢桿菌(Bacillus subtilis sp.),1-3為類芽孢桿菌(Paenibacillus sp.),1-4為韓國假單胞菌(Pseudomonas koreensis sp.),1-5、1-6、2-3為解淀粉芽孢桿菌(Bacillus amyloliquefaciens sp.),2-2為蒼白桿菌(Ochrobactrum sp.),3-6為副球菌(Paracoccus sp.)
2.3 馬鈴薯內生細菌的DGGE指紋圖譜分析結果
依據(jù)DGGE圖譜分析可知(圖3),馬鈴薯塊莖的內生細菌種類較多,且不同馬鈴薯品種之間存在差異性。采用Quantity One 分析軟件對DGGE圖譜進行聚類分析,并生成系統(tǒng)樹(UPGMA)發(fā)現(xiàn),相同種植地區(qū)不同品種的馬鈴薯內生細菌在DGGE圖譜中電泳的條帶數(shù)目、亮度、遷移率均存在明顯的差異。由系統(tǒng)樹可知(圖4),相同地區(qū)不同品種的馬鈴薯內生細菌的群落結構的差異性較大。

圖2 基于16S rDNA序列的馬鈴薯內生細菌系統(tǒng)發(fā)育樹括號中的代碼表示GenBank登錄號。

表2 9株內生細菌的生理生化試驗結果和細胞特征
其中P1、P2的條帶數(shù)目相對較多,說明泰山1號、中薯5號的內生細菌的菌落結構最為復雜,P3條帶數(shù)目較少并且亮度較弱,說明荷蘭15號的內生細菌的菌落結構相對簡單。除P1、P2、P3中的共有條帶外,p1-6、p1-10為P1特有條帶;p2-1為P2特有條帶。這些條帶的分布特征也可以說明相同種植地區(qū)不同品種的馬鈴薯內生細菌有較大的群落差異。

圖3 3個馬鈴薯品種內生細菌的DGGE分析P1、P2、P3分別代表泰山1號、中薯5號、荷蘭15號馬鈴薯內生細菌的DGGE條帶。
3 結論與討論
3.1 馬鈴薯內生細菌細菌株
利用生理生化鑒定可以將微生物的不同類群分開。將16S rDNA基因序列同源性分析與生理生化試驗相結合是一種能夠比較快速精確分離細菌的方法。本試驗中系統(tǒng)發(fā)育學分析有較好的相關性。
16S rRNA 基因序列分析表明,菌株1-1、1-2為枯草芽孢桿菌(Bacillus subtilis sp.),1-5、1-6、2-3為解淀粉芽孢桿菌(Bacillus amyloliquefaciens sp.),這5株內生細菌均屬于芽孢桿菌屬。因為芽孢桿菌在分離中的內生細菌有較大比重,所以初步將芽孢桿菌確定為3種馬鈴薯的優(yōu)勢菌株。此外1-3為類芽孢桿菌(Paenibacillus sp.)、1-4為韓國假單胞菌(Pseudomonas koreensis sp.)、3-6為副球菌(Paracoccus sp.)這3類菌株均為植物內生菌的常見菌株。2-2為蒼白桿菌(Ochrobactrum sp.),雖然關于蒼白桿菌的植物內生菌相關報道較少,但也有人在植物根部分離到蒼白桿菌(劉杰 等,2011;劉芳 等,2014)。
芽孢桿菌是一種有較高農業(yè)生產價值的細菌,具有繁殖速度快、生命力強、體積大等特征(Stenberg,1999)。特別是在抑菌方面有較大的開發(fā)價值,能夠較好地抑制有害菌、病原菌等有害微生物的的生長繁殖,在生防菌肥的工業(yè)化生產中已經被投入使用。芽孢桿菌還能產生豐富的代謝生成物,從而被農作物利用促進作物生長(張霞 等,2007)。本試驗所篩選到的內生菌種類在應用上有較高的價值,后續(xù)的研究仍待開展。
3.2 DGGE
關于內生菌的分離鑒定方法一般都是依賴于傳統(tǒng)的平板分離,這種方法簡單易行,較為普遍,但其分離得到的菌株都為可培養(yǎng)的細菌,而無法分離不可培養(yǎng)的內生菌。近年來關于利用非培養(yǎng)方法研究微生物種群變得普遍起來。而DGGE又是一種常用的非培養(yǎng)手段研究微生物種群的方法,本試驗根據(jù)電泳所得DGGE圖譜的條帶數(shù)量和亮度對馬鈴薯內生細菌進行研究。但是DGGE方法還是有一定的局限性,包括DNA模板長度過短、PCR擴增的偏向性、無法獲得菌株等制約因素的存在。
本試驗對相同地點種植的不同品種馬鈴薯內生細菌進行DGGE分析,結果表明相同地區(qū)種植的不同品種的馬鈴薯內生細菌的群落結構差異性較大(圖4)。由于這3種不同品種的馬鈴薯生長于相同的地點,所以也受到相似的外部環(huán)境因素的影響,在排除外部因素的影響后,推測造成這種內生菌的群落結構差異的主要原因是馬鈴薯品種這一內因的差異。不同品種馬鈴薯的生長代謝不同造就了馬鈴薯內生細菌所生長的外環(huán)境的差異,這是造成內生菌群落差異的根本原因。
而這些不同品種馬鈴薯的內生細菌中還有一些共有菌株,這說明不同品種馬鈴薯內生細菌除了差異性之外還有共有性,一些生態(tài)幅較廣的內生細菌對不同環(huán)境有較強的適應性,從而造成了這種結果。
此外,在DGGE圖譜中條帶p1-11、p2-10、p3-8條為P1、P2、P3的共有條帶,相比于其他條帶也更亮,而本試驗所篩選出的內生細菌中芽孢桿菌的數(shù)量占絕大部分,故推測該共有條帶所對應的內生細菌為本試驗所篩選出的芽孢桿菌,是本試驗所用樣品內生細菌的優(yōu)勢菌株,但也不排除該條帶對應不可培養(yǎng)內生菌的可能性。
參 考文獻
陳雪英,都曉偉,李斌.2008.內生菌與藥用活性成分相關性研究進展.國外醫(yī)藥:植物藥分冊,23(2):47 52.
何勁,劉蘊哲,康冀川.2006.植物內生菌及其在農業(yè)和醫(yī)學上的用途.貴州農業(yè)科學,(3):113-115.
李瑞,郭利偉,郭偉云,李延蘭.2009.抗腫瘤藥用植物及其內生菌活性代謝產物的研究.安徽農業(yè)科學,37(16):7508-7509.
林榕姍.2012.細菌型豆豉發(fā)酵機理及功能性研究〔博士論文〕.泰安:山東農業(yè)大學.
劉芳,袁宗勝,張國防,陳威.2014.毛竹內生細菌種群多樣性分析.福建農業(yè)學報,29(12):1236-1239.
劉杰,汪恩濤,陳文新.2011.豆科植物根瘤內生細菌的發(fā)現(xiàn)及其研究進展.微生物學報,51(8):1010-1006.
劉敏,朱開玲,李洪波,張濤,尚天.2000.應用PCR-DGGE技術分析黃海冷水團的細菌群落組成.生態(tài)學報,20(6):1021-1027.
石晶盈,陳維信,劉愛媛.2006.植物內生菌及其防治植物病害的研究進展.生態(tài)學報,26(7):2395-2401.
文才藝,吳元華,田秀玲.2004.植物內生菌研究進展及其存在的問題.生態(tài)學雜志,23(2):86-91.
張霞,唐文華,張力群.2007.枯草芽孢桿菌B931防治植物病害和促進植物生長的作用.作物學報,33(2):236-241.
張一,王鳳忠,杜秉海,靳志剛,李研,韓明渠.2012.基于變性梯度凝膠電泳的青縣蘋果再植障礙園與豐產園土壤細菌多樣性研究.山東農業(yè)科學,44(1):71-74.
Fisher P J,Petrini O,Sutton B C.1993.A comparative study of fungal endophytes in leaves,xylem and bark of Eucalyptus in Australia and England.Sydowia,45:338-345.
Herre E A,Mejia L C,Kyllo D A,Rojas E,Maynard Z,Butler A,Van Bael S A.2007.Ecological implication of anti-pathogen effects of tropical fungal endophytes and mycorrhizae.Ecology,88 (3):550-558.
Kumar N R,Raman R P,Jadhao S B,Brahmchari R K,Kumar K,Dash G.2013.Effect of dietary supplementation of Bacillus licheniformis on gut microbiota,growth and immune response in giant freshwater prawn,Macrobrachium rosenbergii(de Man,1879).Aquaculture International,(2):387-403.
Munif A,Hallmann J,Sikora R A.2001.Induced systemic resistance of selected endophytic bacteria against Meloidogyne incognita on tomato.Meded Rijksuniv GentFak Landboenvkd Toegep Biol Wet,66(2b):663-669.
Stenberg B.1999.Monitoring soil quality of arable land:microbiological indicators.Soil Plant Sci,49(1):1-24.
Stone J K,Bacon C W,White J F Jr.2000.An overview of endophytic microbes:endophytism defined.New York:Microbial endophytes:29-33.
Ying M,Rajkumar M,Luo Y M,F(xiàn)reitas H.2011.Inoculation of endophytic bacteria on host and non-host plants-effects on plants growth and Ni uptake.Journal of Hazardous Materials,195:30-237.
Isolation and Identification of Endophytic Bacteria and Analysis of Population Diversity in Different Potato Varieties
QU Qing-song,PENG Wan-li,ZHAI Li-ming,LIU Xiao,LIN Rong-shan*
(College of Life Science,Shandong Agricultural University,Shandong Agricultural Resource Collection of Microorganisms,Tai'an 271018,Shandong,China)
Abstract:In order to isolate and identify endophytic bacteria in potato and analyze the community structure of 3 different potato varieties,this experiment isolated and purified strains from different potato varieties,and carried out gene sequence analysis and physiological and biochemical identification.The PCR-DGGE operation was performed on the same site in potato,and analyzed the bands on DGGE.Nine strains of endophytic bacteria were isolated,identified 1-1,1-2 as Bacillus subtilis sp.,1-3 as Paenibacillus sp.,1-4 as Pseudomonas koreensis sp.,1-5、1-6、2-3 as Bacillus amyloliquefaciens sp.,2-2 as Ochrobactrum sp.,3-6 as Paracoccus sp.The DGGE bands indicated that there were both differences and similarities in different potato varieties.The results showed that 3 potato varieties had rich population diversity,and their differences were also obvious.The species may be an important factor influencing the structure of endophytic bacteria community.
Key word:DGGE;Potato;Endophytic bacteria;Identification
*通訊作者(
Corresponding author):林榕姍,女,副教授,碩士生導師,專業(yè)方向:資源與環(huán)境微生物,E-mail:lrs2680@163.com
收稿日期:2016-01-18;接受日期:2016-04-20
基金項目:國家級大學生創(chuàng)新創(chuàng)業(yè)訓練計劃項目(201410434084)
