中耳炎影響內(nèi)耳的分子機理探索
2016-05-24 00:35:40王亞娜
中國實驗診斷學 2016年4期
王亞娜,張 良
(天津市公安醫(yī)院 耳鼻喉科,天津300042)
?
中耳炎影響內(nèi)耳的分子機理探索
王亞娜,張良*
(天津市公安醫(yī)院 耳鼻喉科,天津300042)
中耳炎是兒童僅次于上呼吸道感染的第二大常見疾病,病理特點包括中耳(例如咽鼓管、鼓竇、鼓室及乳突氣房)全部或部分結(jié)構(gòu)的炎性病變,發(fā)病原因主要是細菌及病毒感染[1]。急性中耳炎是中耳黏膜的急性化膿性炎癥,通常持續(xù)數(shù)天[2]。慢性中耳炎會將病變炎癥部位延伸至內(nèi)耳[3]。持續(xù)的內(nèi)耳損傷可以導致感覺神經(jīng)性聽力喪失、阻礙語言能力的發(fā)展及社交障礙[4,5]。有效的控制內(nèi)耳炎癥的后遺癥是保護青少年聽力的重要環(huán)節(jié),然而導致內(nèi)耳炎癥的發(fā)病機制仍不清楚。
利用分子生物學手段研究中耳炎是當前該領(lǐng)域研究的發(fā)展趨勢;而通過基因芯片或測序技術(shù)和生物信息學分析,在轉(zhuǎn)錄水平對基因的表達調(diào)控網(wǎng)絡(luò)及機制進行全面而系統(tǒng)的分析,是篩選疾病相關(guān)基因最有效的途徑。本研究采用表達譜基因芯片技術(shù),6個正常小鼠內(nèi)耳組織和8個中耳炎小鼠內(nèi)耳組織進行分析,探討差異表達基因之間的相互作用關(guān)系,并預(yù)測可能調(diào)控這些基因表達的miRNA,在整體轉(zhuǎn)錄水平上分析中耳炎的發(fā)病機理。
1材料與方法
1.1基因表達譜數(shù)據(jù)
本次研究的基因表達譜數(shù)據(jù)GSE49122[6]取自NCBI的GEO數(shù)據(jù)庫(http://www.ncbi.nlm.nih.gov/geo/),疾病組織類型為中耳炎內(nèi)耳組織,包括6個正常小鼠內(nèi)耳組織和8個中耳炎小鼠內(nèi)耳組織。提取出組織中的RNA之后,基于GPL1261 Affymetrix Mouse Genome 430 2.0 Array平臺探測它們在組織中的表達值。
1.2芯片數(shù)據(jù)處理及數(shù)據(jù)分析
下載得到原始CEL格式數(shù)據(jù)后,導入R編程語言,基于affy[7]函數(shù)包進行背景校準、去除組間差異標準化,得到可以進行后續(xù)分析的基因表達值矩陣格式數(shù)據(jù)。隨后,利用limma[8]函數(shù)包,對基因在中耳炎和正常小鼠內(nèi)耳組織中的表達值進行student’st檢驗,并利用Benjamini&Hochberg(BH)校正方法對得到的P值進行校正,把滿足表達值變化倍數(shù)大于2或者小于0.5并且校正P值小于0.05的基因作為中耳炎小鼠內(nèi)耳組織相對于正常小鼠內(nèi)耳組織的差異表達基因(DEGs)。
1.3功能與通路富集分析
在得到DEGs之后,利用DAVID(http://david.abcc.ncifcrf.gov/)[9]在線工具對它們進行功能和通路富集分析。以P值小于0.05為閾值,篩選DEGs富集的基因本體條目(GO terms)和通路(KEGG pathways)。
1.4miRNA-Gene調(diào)控網(wǎng)絡(luò)構(gòu)建
miRNA是一種長度大約在22-25nt的非編碼RNA,它可以在翻譯過程中通過抑制基因翻譯或者降解蛋白來抑制基因的表達。TargetScan(http://www.targetscan.org/)[10]是一個收錄了包括人類和小鼠在內(nèi)的多個物種的miRNA與基因之間調(diào)控關(guān)系的數(shù)據(jù)庫,本次研究通過TargetScan數(shù)據(jù)庫,得到了可能調(diào)控DEGs的miRNA,并且利用Cytoscape[11]軟件構(gòu)建了miRNA-Gene調(diào)控網(wǎng)絡(luò)。
1.5基因相互作用網(wǎng)絡(luò)構(gòu)建
在生物體的大部分生物過程中,基因都通過與其他基因的相互作用來發(fā)揮作用,而不是單獨發(fā)揮作用。STRING(http://string-db.org/)[12]數(shù)據(jù)庫通過整理已有的諸如高通量數(shù)據(jù)、實驗室低通量數(shù)據(jù)等信息來源,形成了許多物種的基因之間的相互作用,并以一個綜合得分來評估一對相互作用的可靠性。本次研究通過STRING數(shù)據(jù)庫,以綜合得分大于0.4為閾值,篩選出基因之間的相互作用對,并通過Cytoscape軟件構(gòu)建出基因相互作用網(wǎng)絡(luò)(PPI)。
2結(jié)果
2.1芯片數(shù)據(jù)分析結(jié)果
預(yù)處理之后,芯片上的基因在各個樣本中的表達值見圖1A,由A可以得到標準化后芯片上所有基因的整體表達值在各樣本之間基本上是一致的,這說明預(yù)處理消除了組間差異,使后續(xù)的差異分析更加可靠。與正常小鼠內(nèi)耳組織相比,在中耳炎小鼠內(nèi)耳組織中得到了195個DEGs(圖1B),并且由圖1C可知DEGs能夠很好的將正常和中耳炎小鼠分開,這也證明了得到的DEGs的可靠性。其中,前10個下調(diào)和上調(diào)倍數(shù)最多的基因見表1。
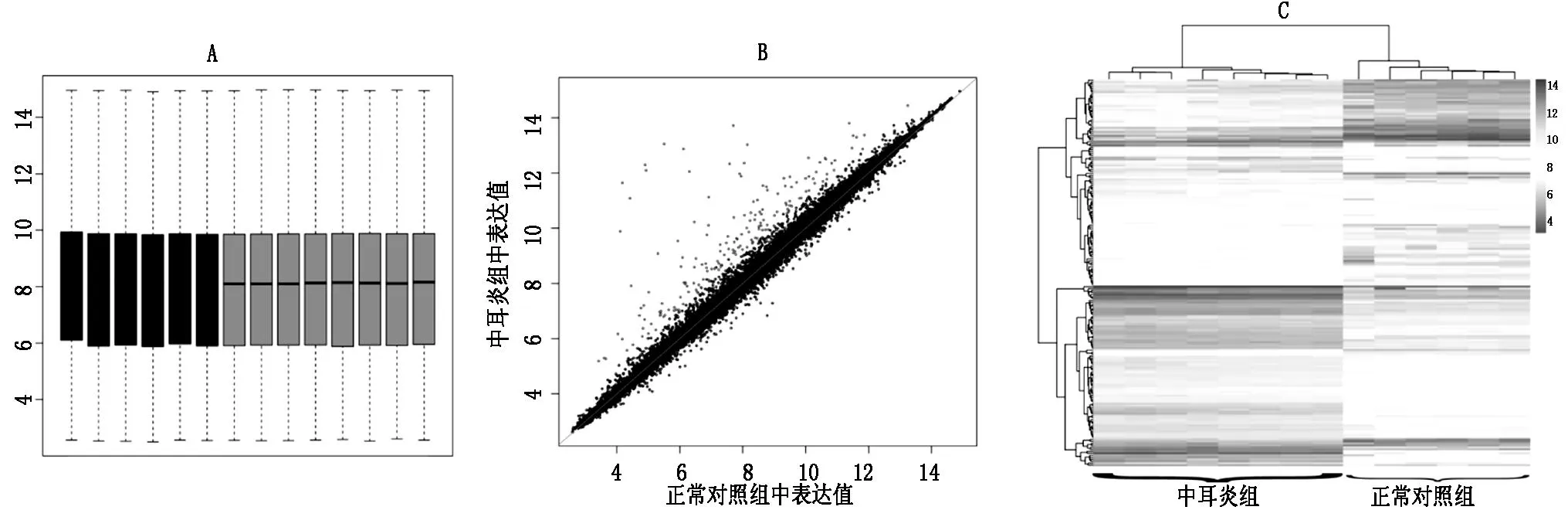
圖1芯片數(shù)據(jù)的預(yù)處理及差異表達基因篩選A:標準化后芯片上所有基因在每個樣本中的表達值,藍色樣本是正常小鼠樣本,綠色樣本是中耳炎小鼠樣本,縱軸是表達值。B:差異表達基因分布,黑點是非差異表達基因,紅點是在中耳炎小鼠中表達上調(diào)的基因,共有176個,藍點是在中耳炎小鼠中表達下調(diào)的基因,共有19個。C:差異表達基因及樣本的雙向聚類。
表1在中耳炎樣本中下調(diào)和上調(diào)倍數(shù)最大的前10個基因
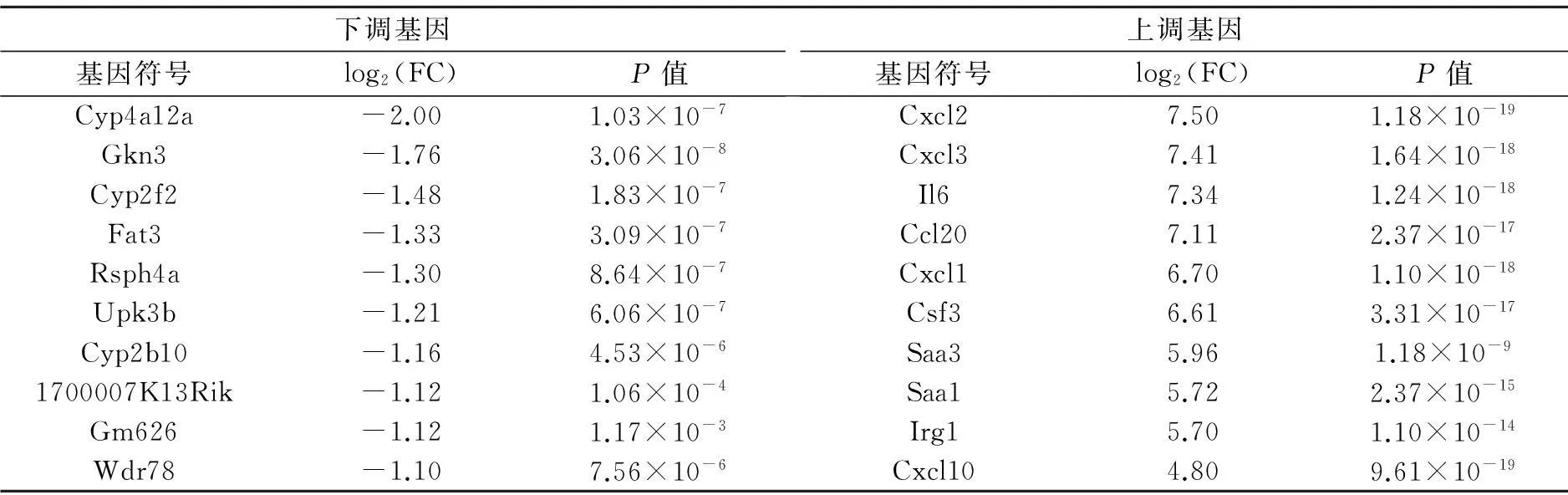
下調(diào)基因基因符號log2(FC)P值上調(diào)基因基因符號log2(FC)P值Cyp4a12a-2.001.03×10-7Cxcl27.501.18×10-19Gkn3-1.763.06×10-8Cxcl37.411.64×10-18Cyp2f2-1.481.83×10-7Il67.341.24×10-18Fat3-1.333.09×10-7Ccl207.112.37×10-17Rsph4a-1.308.64×10-7Cxcl16.701.10×10-18Upk3b-1.216.06×10-7Csf36.613.31×10-17Cyp2b10-1.164.53×10-6Saa35.961.18×10-91700007K13Rik-1.121.06×10-4Saa15.722.37×10-15Gm626-1.121.17×10-3Irg15.701.10×10-14Wdr78-1.107.56×10-6Cxcl104.809.61×10-19
注:log2(FC)中FC是指變化倍數(shù)。
2.2富集的功能和通路
把DEGs上傳到DAVID數(shù)據(jù)庫中,發(fā)現(xiàn)這些基因主要富集于與炎癥或者免疫等生物過程相關(guān)的GO terms及KEGG pathways。其中,按照P值最顯著的前20個GO terms見圖2,得到的13條富集的通路見表2。
2.3在TargetScan數(shù)據(jù)庫中,得到了可能調(diào)控DEGs的miRNA共115個,它們與DEGs形成了319個miRNA-Gene調(diào)控關(guān)系對。其中,miRNA-Gene調(diào)控網(wǎng)絡(luò)見圖3A。在miRNA-Gene網(wǎng)絡(luò)中,基因所受到的miRNA的直接調(diào)控個數(shù)稱為該基因的節(jié)點度。
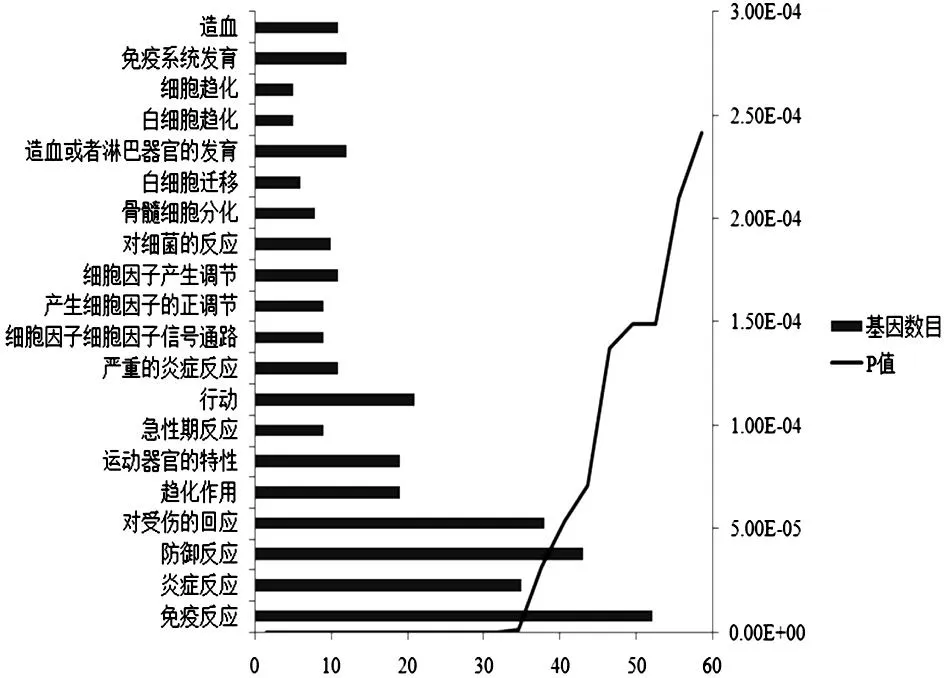
圖2 差異表達基因富集的GO terms
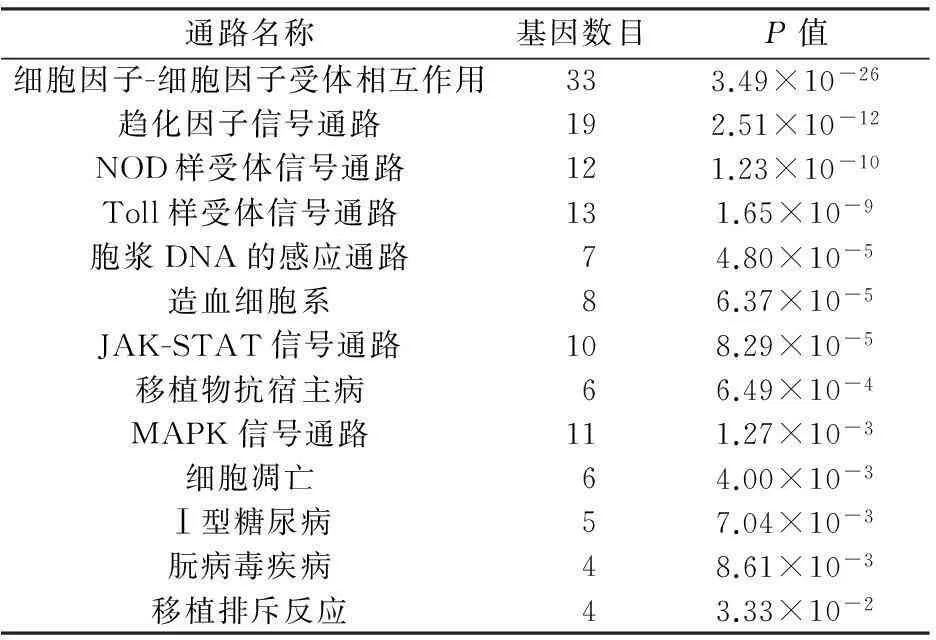
通路名稱基因數(shù)目P值細胞因子-細胞因子受體相互作用333.49×10-26趨化因子信號通路192.51×10-12NOD樣受體信號通路121.23×10-10Toll樣受體信號通路131.65×10-9胞漿DNA的感應(yīng)通路74.80×10-5造血細胞系86.37×10-5JAK-STAT信號通路108.29×10-5移植物抗宿主病66.49×10-4MAPK信號通路111.27×10-3細胞凋亡64.00×10-3Ⅰ型糖尿病57.04×10-3朊病毒疾病48.61×10-3移植排斥反應(yīng)43.33×10-2
2.4PPI網(wǎng)絡(luò)
在STRING數(shù)據(jù)庫中,以綜合得分大于0.4為閾值,共得到了828個基因之間的相互作用對。其中,PPI網(wǎng)絡(luò)見圖3B。與miRNA-Gene網(wǎng)絡(luò)類似,在PPI網(wǎng)絡(luò)中與某基因有直接相互作用的基因的個數(shù)稱為該基因的節(jié)點度。并且我們發(fā)現(xiàn),有10個基因在PPI網(wǎng)絡(luò)和miRNA-Gene調(diào)控網(wǎng)絡(luò)中的節(jié)點度都大于等于5(表3),并且這些基因在中耳炎小鼠內(nèi)耳組織表達值相對于正常小鼠內(nèi)耳組織表達值都發(fā)生了較大的變化(圖4),所以這些基因可能在中耳炎對內(nèi)耳產(chǎn)生影響中有較重要的作用。
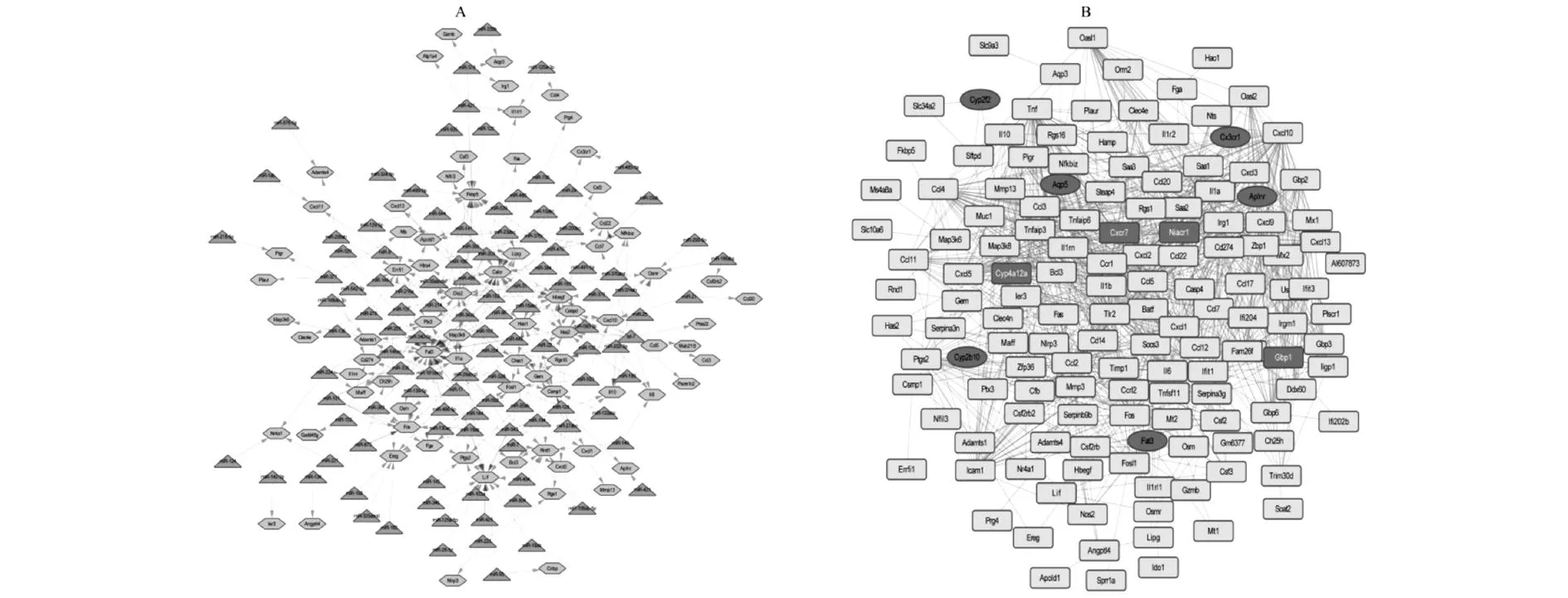
圖3差異表達基因的miRNA-Gene調(diào)控網(wǎng)絡(luò)和PPI網(wǎng)絡(luò)。A:miRNA-Gene調(diào)控網(wǎng)絡(luò),淺藍色六邊形是差異表達基因,綠色三角形是miRNA。B:PPI網(wǎng)絡(luò),深藍色長方形是在中耳炎小鼠中表達下調(diào)的基因,紅色長方形是在中耳炎小鼠中表達上調(diào)的基因
表3在miRNA-Gene和PPI網(wǎng)絡(luò)中節(jié)點度都大于等于5的基因
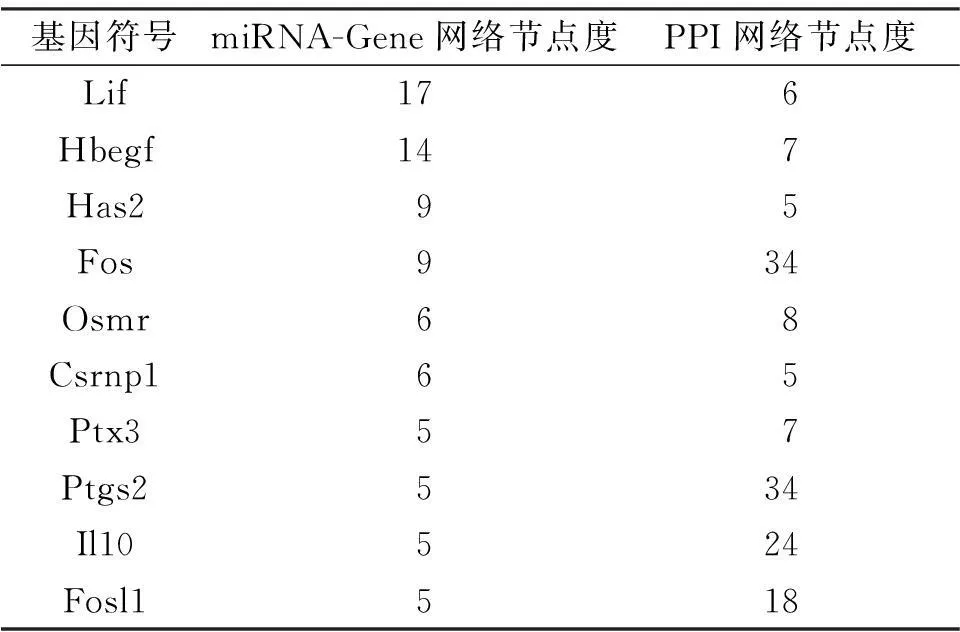
基因符號miRNA-Gene網(wǎng)絡(luò)節(jié)點度PPI網(wǎng)絡(luò)節(jié)點度Lif176Hbegf147Has295Fos934Osmr68Csrnp165Ptx357Ptgs2534Il10524Fosl1518
3討論
中耳炎是臨床上最常見的兒童感染類疾病之一,盡管多數(shù)病例可以得到滿意的治療效果,但接近20%的兒童患者會復發(fā),或轉(zhuǎn)變成慢性中耳炎,并最終導致多種并發(fā)癥的發(fā)生,包括傳導性耳聾和感音神經(jīng)性耳聾,嚴重影響兒童語言能力的發(fā)展。所以更深刻的了解中耳炎發(fā)病過程中的炎癥病理機制,有助于阻止中耳炎及其并發(fā)癥的發(fā)生。
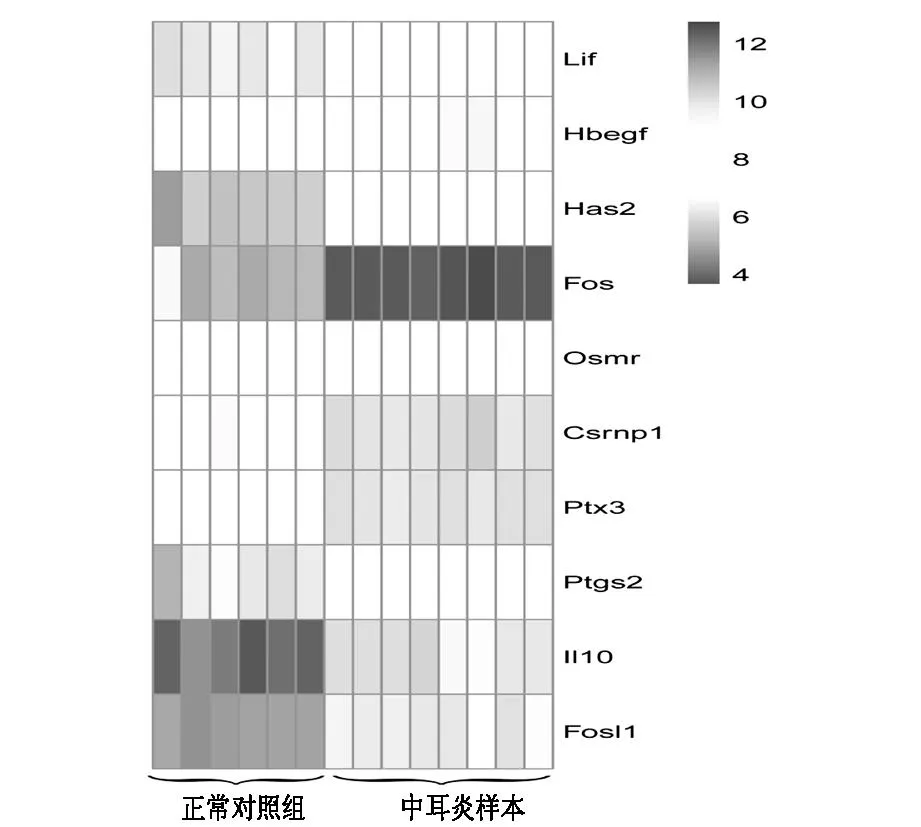
圖4 在PPI和miRNA-Gene網(wǎng)絡(luò)中節(jié)點度都大于等于5的
在本研究中,我們通過分析基因表達譜芯片,對6個正常小鼠內(nèi)耳組織和8個中耳炎小鼠內(nèi)耳組織中基因的表達進行高通量分析,獲得195個差異表達基因,其中表達上調(diào)的有176個,表達下調(diào)的有19個。
GO分析是指對所有差異表達基因進行相應(yīng)的GO功能注釋,隨后對所有GO進行顯著性分析,進而達到對基因的表達及其功能進行系統(tǒng)而全面分析的目的。本研究對195個差異表達基因進行GO分析,結(jié)果發(fā)現(xiàn)差異表達基因主要富集于白細胞趨化、細胞因子產(chǎn)生調(diào)節(jié)、炎癥反應(yīng)和免疫反應(yīng)等GO條目,而且這些生理變化可能和中耳炎的發(fā)病機理密切相關(guān)。KEGG(京都基因與基因組百科全書)是基因組破譯方面的數(shù)據(jù)庫,KEGG 的PATHWAY 數(shù)據(jù)庫整合當前在信號通路和分子互動網(wǎng)絡(luò)信息。本研究對195個差異表達基因進行KEGG PATHWAY分析,結(jié)果發(fā)現(xiàn)差異表達基因主要富集于細胞因子-細胞因子受體相互作用、趨化因子信號通路和MAPK信號通路等信號通路。這些信號通路與免疫系統(tǒng)的激活密切相關(guān),提示這些基因和信號通路可能在中耳炎的發(fā)病過程中發(fā)揮重要的作用。
Hbegf (Heparin-Binding EGF-Like Growth Factor)是一個蛋白編碼基因,參與PI-3K信號通路、生長因子活性和上皮生長因子受體的綁定,其表達水平與圍產(chǎn)期壞死性小腸結(jié)腸炎和白喉密切相關(guān)。Suzukawa等[13]報道,在小鼠中耳炎模型中,HB-EGF促進中耳黏膜上皮的增生,并提示HB-EGF可能是改善黏膜生長的治療靶點,進而阻止這種兒童疾病后遺癥的發(fā)生。在我們的研究中,發(fā)現(xiàn)Hbegf在中耳炎小鼠內(nèi)耳組織中的表達同樣顯著升高。我們對該基因進一步進行miRNA-Gene網(wǎng)絡(luò)節(jié)點度和PPI網(wǎng)絡(luò)節(jié)點度分析,發(fā)現(xiàn)該基因的表達受到miR-203、miR-182和miR-194等14個miRNA的調(diào)控,而且該基因可能與Mmp3、Ptgs2和I16等7個差異表達基因相互作用,提示該基因在中耳炎的發(fā)生發(fā)展過程中起至關(guān)重要的作用。
總而言之,我們的結(jié)果結(jié)合其他小組的研究可能為中耳炎的發(fā)病基因及其后遺癥的理解提供了寶貴的理論支持。然而,有些結(jié)果還是有爭議的;所以,我們需要更大的樣本量和分子生物學實驗來進一步解釋相關(guān)miRNA和基因是如何參與中耳炎致病基因的表達。
參考文獻:
[1]Pichichero ME.Recurrent and persistent otitis media[J].Pediatr Infect Dis J,2000,19:911.
[2]Juhn SK,Jung TT,Lin J,et al.Effects of inflammatory mediators on middle ear pathology and on inner ear function[J].Ann N Y Acad Sci,1997,830:130.
[3]Smirnova MG,Kiselev SL,Gnuchev NV,et al.Role of the pro-inflammatory cytokines tumor necrosis factor-alpha,interleukin-1 beta,interleukin-6 and interleukin-8 in the pathogenesis of the otitis media with effusion[J].Eur Cytokine Netw,2002,13:161.
[4]Schachern P,Tsuprun V,Cureoglu S,et al.The round window membrane in otitis media:effect of pneumococcal proteins[J].Arch Otolaryngol Head Neck Surg,2008,134:658.
[5]Moon SK,Woo JI,Lee HY,et al.Toll-like receptor 2-dependent NF-kappaB activation is involved in nontypeable Haemophilus influenzae-induced monocyte chemotactic protein 1 up-regulation in the spiral ligament fibrocytes of the inner ear[J].Infect Immun,2007,75:3361.
[6]MacArthur CJ,Hausman F,Kempton JB,et al.Otitis media impacts hundreds of mouse middle and inner ear genes[J].PLoS One,2013,8:e75213.
[7]Gautier L,Cope L,Bolstad BM,et al.Affy-analysis of Affymetrix GeneChip data at the probe level[J].Bioinformatics,2004,20:307.
[8]Diboun I,Wernisch L,Orengo CA,et al.Microarray analysis after RNA amplification can detect pronounced differences in gene expression using limma[J].BMC Genomics,2006,7:252.
9.Dennis G,Sherman BT,Hosack DA,et al.DAVID:Database for Annotation,Visualization,and Integrated Discovery[J].Genome Biol,2003,4:P3.
[10]Agarwal V,Bell GW,Nam JW,et al.Predicting effective microRNA target sites in mammalian mRNAs[J].Elife,2015,doi:10.7554/eLife.05005.PubMed - in process.
[11]Shannon P,Markiel A,Ozier O,et al.Cytoscape:a software environment for integrated models of biomolecular interaction networks[J].Genome Res,2003,13:2498.
[12]Szklarczyk D,Franceschini A,Wyder S,et al.STRING v10:protein-protein interaction networks,integrated over the tree of lif[J]e.Nucleic Acids Res,2015,43:D447.
[13]Suzukawa K,Tomlin J,Pak K,et al.A mouse model of otitis media identifies HB-EGF as a mediator of inflammation-induced mucosal proliferation[J].plos one,2014,9:e102739.
(收稿日期:2015-09-26)
*通訊作者
文章編號:1007-4287(2016)04-0540-04
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
財經(jīng)(2017年2期)2017-03-10 14:35:35
財經(jīng)(2016年15期)2016-06-03 07:38:02
財經(jīng)(2016年3期)2016-03-07 07:44:46
財經(jīng)(2016年6期)2016-02-24 07:41:51
鄭州大學學報(醫(yī)學版)(2015年2期)2015-02-27 14:50:46
