長牡蠣外套膜色遺傳規律初步研究
2015-12-02 03:09:54王慶志叢日浩孔令鋒
海洋科學 2015年11期
關鍵詞:生長
王慶志 , 李 琪 叢日浩 , 孔 寧 孔令鋒 于 紅
(1. 中國海洋大學 海水養殖教育部重點實驗室, 山東 青島 266003; 2. 遼寧省海洋水產科學研究院, 遼寧大連 116023; 3. 中國科學院 海洋研究所, 山東 青島 266071)
在海產經濟貝類中, 許多種類色彩繽紛的貝殼顏色, 長期受到遺傳學家、育種學家的關注。美觀的殼色不僅給消費者帶來視覺享受, 還能顯著提高商品價值。許多貝類的殼色性狀屬于受遺傳與環境雙重影響的數量性狀, 它與生物所處的溫度、鹽度、底質、食物等棲息環境有關[1-2]。一些研究也發現貝類的殼色主要受遺傳因素控制, 并與生長、存活等性狀相關。在選擇育種工作中, 當某一性狀由少數基因和簡單的顯隱性機制決定時, 通過合適的交配設計可使目的性狀分離, 經連續幾代選擇便可獲得純系[3],我國已成功培育出‘中國紅’皺紋盤鮑(Haliotis discus hannai)[4]、‘中科紅’海灣扇貝(Argopecten irradians)[5]、‘蓬萊紅’櫛孔扇貝(Chlamys farreri)[6]和“斑馬蛤”菲律賓蛤仔(Ruditapes philippinesis)[7]等優良新品系。除了較易觀察到的貝殼顏色存在明顯的遺傳規律外, 一些研究還發現貝類的閉殼肌、外套膜等組織的顏色也存在明顯的遺傳規律, 如蝦夷扇貝(Mizuhopecten yessoensis)的橘紅色閉殼肌就屬于單基因控制的質量性狀[8], 育種工作者據此成功選育出橘紅色閉殼肌的“海大金貝”新品種。
長牡蠣(Crassostrea gigas)又稱太平洋牡蠣, 具有生長速度快、肉質鮮美等優點, 是世界上產量最高的海水養殖種類[9]。筆者在長期的育種實踐中發現長牡蠣的外套膜顏色由于色素沉著程度不同, 表現出介于黃、黑之間變化的連續數量性狀[10-11]。在進行長牡蠣市場銷售時, 日本消費者較喜歡黑色外套膜的“半殼貝”牡蠣, 而進行深加工時, 黃色外套膜因色素少而更受歡迎。因此, 選育長牡蠣外套膜的不同純色品系, 可以滿足不同的市場需求, 進而提高長牡蠣的商品價值。本研究以外套膜不同著色程度的長牡蠣為親本構建了不同外套膜色家系, 通過分析其子代的表型性狀, 查清了長牡蠣外套膜色與生長、存活的關系, 并初步分析了長牡蠣外套膜色的遺傳規律, 旨在為長牡蠣的優良品種培育提供科學依據。
1 材料和方法
1.1 家系構建
本研究所用材料為長牡蠣威海本地養殖群體。參考王慶志等[10]的外套膜著色 1-4級分類標準, 從中選取外套膜色為4級純黑、1級純黃和2-3級的個體為親本。參考王慶志等[12]的方法構建全同胞家系,每個雄性牡蠣與1個雌性牡蠣交配, 成功構建3個外套膜純黑家系(BB1、BB2、BB3), 2個外套膜純黃家系(YY1、YY2), 3個純黑雌×純黃雄雜交家系(BY1、BY2、BY3), 2個純黃雌×純黑雄雜交家系(YB1、YB2)和 2個 2-3級著色對照家系(CC1、CC2), 家系親本信息見表1。
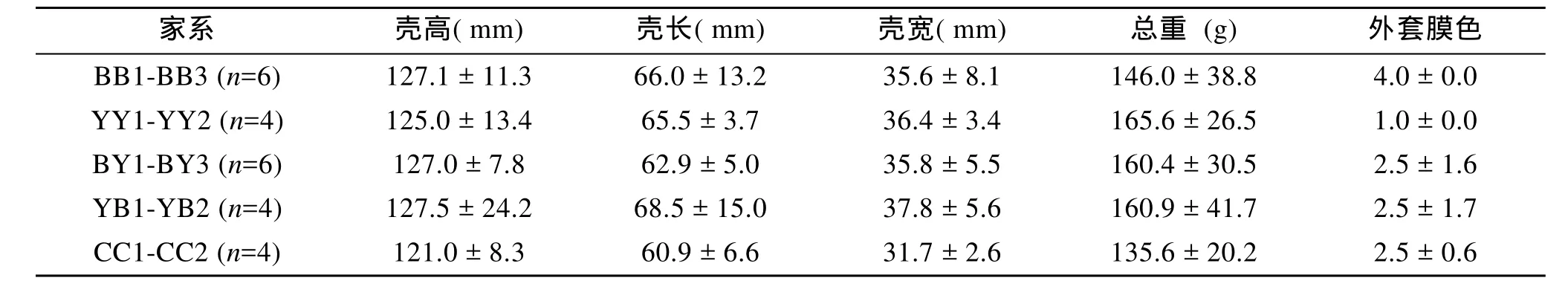
表1 長牡蠣不同外套膜色家系的親本性狀Tab.1 Parents’ traits with different mantle pigmentation families in Crassostrea gigas (±SD)
1.2 幼體培育與養成
將構建的家系在 100 L聚乙烯桶中進行孵化培育, 測量計算不同家系的卵徑、受精率、孵化率等參數。浮游幼蟲期密度控制在2~5個/mL, 投喂適量的單胞藻餌料, 各家系投餌、換水、充氣等管理一致,避免相互混雜。當幼蟲出現眼點時, 投放扇貝殼制成的固著基采苗。參考王慶志等[13]的方法進行稚貝與成體養成, 當稚貝生長至0.5~1cm時, 轉入扇貝養殖籠養成。為減小環境等外因造成的誤差, 轉入扇貝籠時盡量使各層稚貝的密度相同。至150日齡后, 將其分離成單體牡蠣, 每層放30個。
1.3 性狀測量
對測量家系的籠層進行標記, 每次均測量同層牡蠣, 記錄存活個體數。分別測量了牡蠣60日齡、150日齡、350日齡和450日齡的殼高, 350日齡和450日齡的成活率, 按照外套膜著色1-4級分類標準,統計 450日齡的外套膜著色情況, 每一家系測量統計50個。
1.4 統計分析
所有統計分析用SPSS 16.0軟件完成, 用單因素方差分析(ANOVA)比較不同外套膜色組間和組內家系間的差異, 顯著性水平為 P<0.05。為增加數據的正態性, 進行方差分析時, 受精率、孵化率和存活率被轉化為正弦函數值, 殼高被轉化為常用對數。
2 結果
2.1 卵徑、受精率與孵化率
各實家系的卵徑為 55.8~57.4 μm, 受精率為96.9%~98.6%, 孵化率為 75.9%~82.2%, 不同家系間無顯著差異(P>0.05, 表2)。
2.2 生長與存活
60日齡對照組的殼高顯著大于其他交配組(P<0.05), BB1-BB3和 CC1-CC2殼高的差異不顯著(P>0.05); 150日齡CC1-CC2的殼高顯著大于BB1-BB3和YY1-YY2(P<0.05), BB1-BB3和YY1-YY2的殼高差異不顯著(P>0.05); 350日齡和450日齡各家系間差異均不顯著(P>0.05)。存活方面, 350和450日齡各家系在不同生長階段均無顯著差異(P>0.05, 表3)。
2.3 外套膜色遺傳規律
親本外套膜為純黑的3個家系: BB1中100%為4級; BB2中3級占10.4%, 4級占89.6%; BB3中2級占4.3%, 3級占15.2%, 4級占80.4%。親本外套膜為純黃的2個家系: YY1 中95.2%為1級, 4.8%為2級; YY2中1級占27.3%, 2級占45.5%, 3級占24.2%,4級占 3.0%。3個外套膜純黑雌×純黃雄雜交家系:BY1中1級占36.7%, 2級占20.0%, 3級占16.7%, 4級占26.7%; BY2中1級占26.7%, 2級占23.3%, 3級占23.3%, 4級占26.7%; BY3中1級占13.3%, 2級占36.7%, 3級占26.7%, 4級占23.3%。2個純黃雌×純黑雄雜交家系: YB1中1級占10.0%, 2級占20.0%, 3級占36.7%, 4級占33.3%; YB2中2級占34.8%, 3級占30.3%, 4級占19.1%。2個對照家系: CC1中2級占20.0%, 3級占50.0%, 4級占30.0%; CC2中2級占40.0%, 3級占 26.7%, 4級占33.3%(圖 1)。

表2 長牡蠣不同外套膜色家系的卵徑、受精率和孵化率Tab.2 Egg-diameter, fertilized rate and hatching rate of C. gigas with different mantle pigmentation families (±SD)
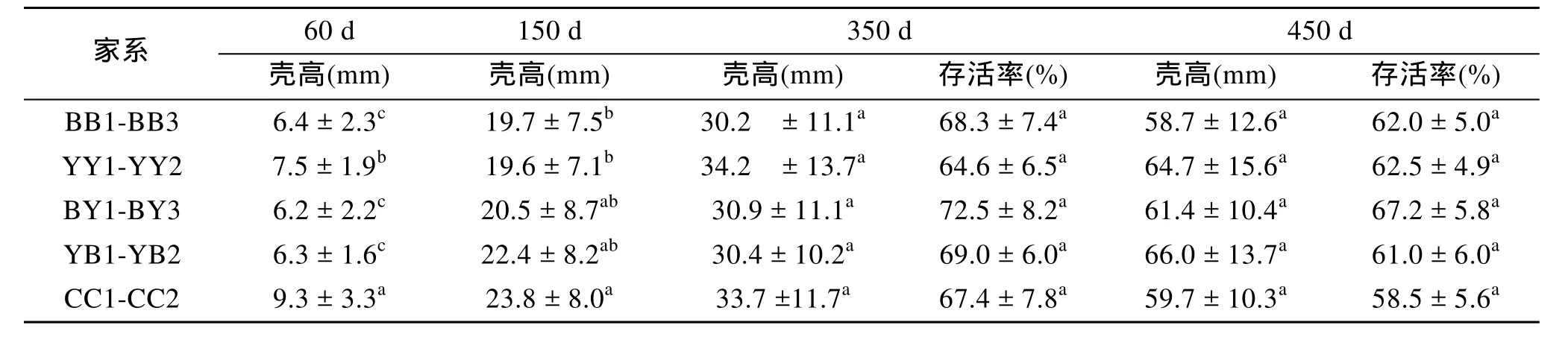
表3 長牡蠣不同外套膜色家系的生長與存活Tab.3 Growth and survival performance of C. gigas in different mantle pigmentation families (±SD)
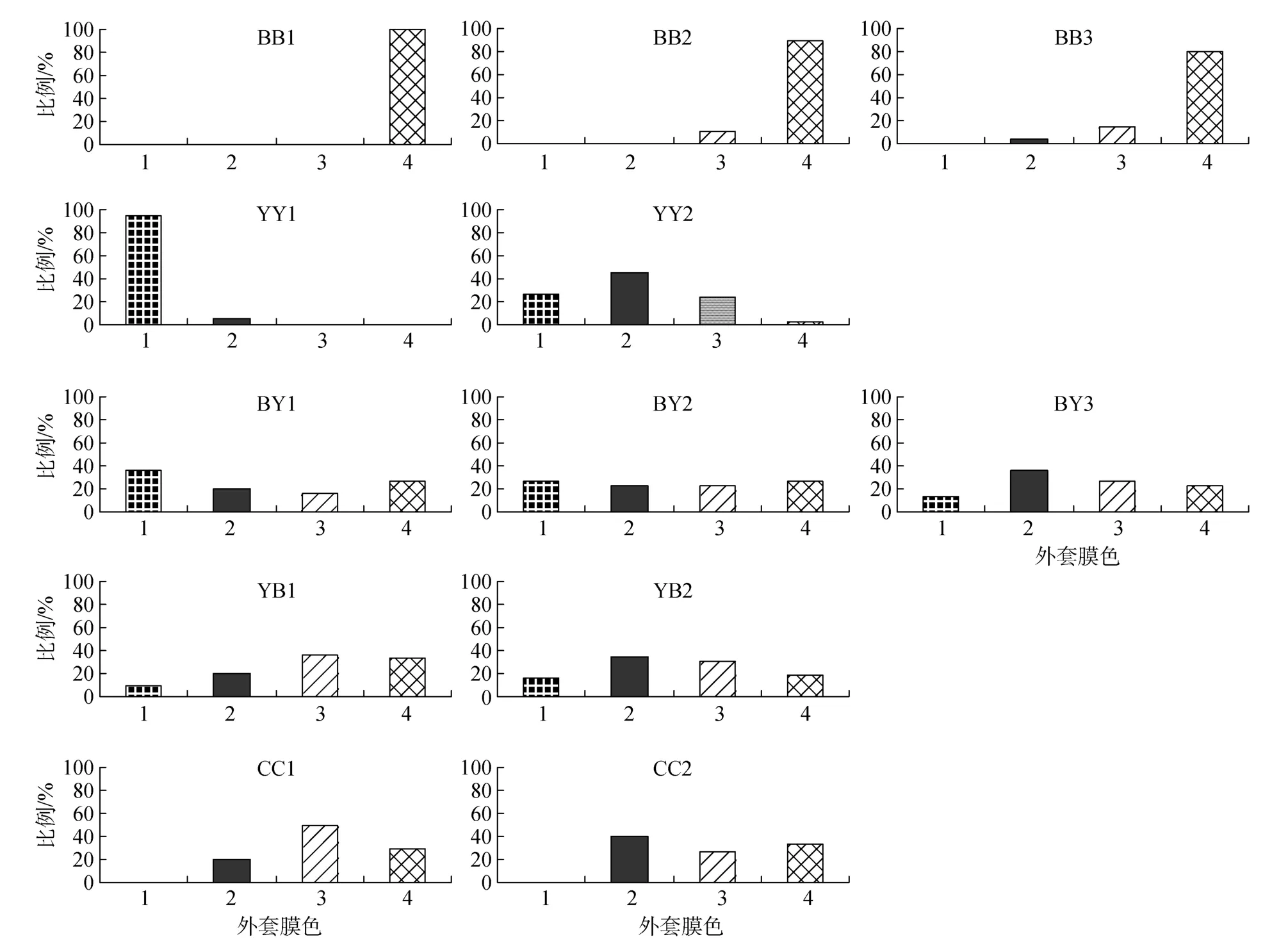
圖 1 外套膜不同著色程度個體在各家系中所占的比例Fig.1 Percentage of individuals with different degree mantle pigmentation in each family
3 討論
在本研究中, 長牡蠣不同外套膜色家系的卵徑、受精率和孵化率均無顯著差異; 不同外套膜色家系的生長雖然在60日和150日齡時略有差異, 但生長至350日齡和450日齡成體階段時, 家系間的生長差異已無顯著差異; 存活方面, 各家系在不同生長階段的存活率無顯著差異。這與 Brake等[14]的研究結果相似, 即牡蠣的殼色、外套膜色與體重與存活沒有顯著的相關性。因此, 進行長牡蠣的外套膜色的人工選育時, 不會對牡蠣的生長和存活帶來負面影響。
研究表明, 長牡蠣的殼色、外套膜色等的變異是由很多微效基因控制的數量性狀, Imai和 Sakai[15]在比較日本沿海不同群體長牡蠣的性狀差異時, 發現不同群體間長牡蠣的外套膜色存在顯著的差異, 且深色外套膜在育種世代中具有可遺傳的特性; 王慶志等[10]也發現中國、日本和韓國群體的長牡蠣在外套膜的著色程度和比例上存在明顯差異; Brake等[14]在構建的大量長牡蠣全同胞家系中發現, 通過一代或兩代的人工選育, 一些家系子代的殼色和外套膜色較好的遺傳了親本的特征, 具有較高的遺傳力;Evans等[16]通過全同胞家系估算長牡蠣殼色的廣義遺傳力和狹義遺傳力分別為 0.91 ± 0.38 和 0.59 ±0.19, 在2個家系中發現“黑殼”和“白殼”性狀的分離符合孟德爾3∶1遺傳分離定律, 并由此推斷殼色如果由一對主效基因控制, 則白色對黑色具有顯性效應。一些研究已證明這些性狀可能由數目很少的主效基因控制, Ge等[17]利用BSA與AFLP相結合的策略, 鑒定出一個與殼色主效基因或 QTL關聯的SCAR標記, 可用于鑒定含有殼色主基因的個體; Ge等[18]還研究了殼色的遺傳模式, 發現長牡蠣貝殼背景色由一個位點的兩個等位基因控制, 金黃色基因相對于白色為顯性, 白殼背景色對于黑殼前景色存在上位顯性效應。
本研究中, 在外套膜純黑的BB1-BB3家系子代中基本全部為 4級著色的個體, 且未發現有外套膜純黃的1級著色個體; 而在外套膜純黃的YY1-YY2家系子代中發現YY1子代基本全部為1級著色, 但在 YY2家系的子代中, 外套膜色出現了明顯的分離現象, 如果將1-2級歸為黃色, 3-4級歸為黑色, 黃色與黑色子代的比例為73∶27, 基本符合3∶1孟德爾遺傳分離定律。因此, 可以推斷長牡蠣外套膜色與長牡蠣殼色的遺傳分離機制基本相同, 即外套膜色可能由一個主效基因控制, 且黃色對黑色具有顯性上位效應。這一結果將為長牡蠣外套膜色的人工選育提供理論依據。同時, 外套膜色在純色家系子代及雜交家系子代中出現的外套膜色連續分布現象也表明,溫度、鹽度、食物等環境因素對外套膜顏色的形成可能有一定程度的影響。
[1] 管云雁, 何毛賢. 海產經濟貝類殼色多態性的研究進展[J]. 海洋通報, 2009, 28(1): 108-114.
[2] Liu X, Wu F, Zhao H, et al. A novel shell color variant of the Pacific abalone Haliotis discus hannai Ino subject togenetic control and dietary influence[J].Journal of Shellfish Resesrch, 2009, 28(2): 419-424.
[3] Falconer D S, MacKay T F C. 儲明星, 譯. 數量遺傳學導論(第 4版)[M]. 北京: 中國農業科技出版社,2000.
[4] 劉曉, 張國范, 趙洪恩. 皺紋盤鮑“中國紅”品系的選育[J]. 動物學雜志, 2003, 38(4): 27.
[5] 許飛, 鄭懷平, 張海濱, 等. 海灣扇貝“中科紅”品種與普通養殖群體不同溫度下早期性狀的比較[J].水產學報, 2008, 32(6): 876-883.
[6] 王偉, 唐玉卿. 雜交櫛孔扇貝試養技術[J]. 水產科學,2003, 22(2): 37-43.
[7] 劉 振, 閆喜武, 張躍環, 等. 菲律賓蛤仔不同品系斑馬蛤的選擇反應及現實遺傳力[J]. 海洋與湖沼,2014, 45(2): 259-264.
[8] 任曉亮, 侯睿, 王珊, 等. 控制蝦夷扇貝閉殼肌積累類胡蘿卜素相關基因的篩查[J]. 中國海洋大學學報,2012, 42(9): 41-47.
[9] FAO (Food and Agriculture Organization). World aquaculture production of fish, crustaceans, molluscs,etc., by principal species [M]. Rome: Food and Agriculture Organization of the United Nations, 2012.
[10] 王慶志, 李琪, 劉士凱, 等. 長牡蠣不同地理群體選育系數量性狀的比較 [J]. 中國海洋大學學報, 2011,41(7/8): 36-41.
[11] 孔寧, 李琪, 叢日浩, 等. 長牡蠣 F3 代快速生長選育群體生長特性的研究[J]. 海洋科學, 2015, 39(3): 7-11.
[12] 王慶志, 李琪, 劉士凱, 等. 長牡蠣幼體生長性狀的遺傳力及其相關性分析 [J]. 中國水產科學, 2009,16(5): 736-743.
[13] 王慶志, 李琪, 劉士凱, 等. 長牡蠣成體生長性狀的遺傳參數估計[J]. 中國水產科學, 2012, 19(4):700-706.
[14] Brake J, Evans F, Langdon C. Evidence for genetic control of pigmentation of shell and mantle edge in selected families of Pacific oysters, Crassostrea gigas[J]. Aquaculture, 2004, 229: 89-98.
[15] Imai T, Sakai S. Study of breeding of Japanese oyster,Crassostrea gigas[J]. Tohoku Journal of Agricultural Research, 1961, 12(1): 125-171.
[16] Evans S, Camara M, Langdon C. Heritability of shell pigmentation in the Pacific oyster, Crassostrea gigas[J].Aquaculture, 2009, 286(3-4): 211-216.
[17] Ge J, Li Q, Yu H, et al. Identification and mapping of a SCAR marker linked to a locus involved in shell pigmentation of the Pacific oyster (Crassostrea gigas)[J]. Aquaculture, 2014, 434(2014): 249-253.
[18] Ge J, Li Q, Yu H, et al. Mendelian inheritance of golden shell color in the Pacific oyster (Crassostrea gigas) [J].Aquaculture, 2015, 441(2015): 21-24.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
