片口自然保護區不同海拔團竹克隆生長研究
2015-08-20 11:25:29劉美張濤譚永燕劉蘭嬌馬李紅
江蘇農業科學 2015年7期
劉美 張濤 譚永燕 劉蘭嬌 馬李紅
摘要:通過對四川省北川縣片口自然保護區不同海拔梯度下團竹的分株形態特征進行調查,研究其克隆生長特征及克隆繁殖的大小依賴關系。結果表明:母株高度、當年生立竹生物量及高度、間隔子直徑、比間隔子長、間隔子分枝強度隨海拔的改變均有顯著性變化;母株、間隔子的生物量受海拔影響不明顯。在各海拔環境中,母株大小均顯著影響間隔子大小,并通過間隔子的作用顯著影響當年生立竹的大小。團竹的克隆生長不僅受海拔梯度的影響,也受自身母株大小的影響,因此可通過改善母株質量而影響克隆分株。
關鍵詞:片口自然保護區;團竹;不同海拔;克隆生長
中圖分類號:S795.201 文獻標志碼: A 文章編號:1002-1302(2015)07-0197-03
團竹(Fargesia obliqua)為禾本科箭竹屬植物,是我國瀕危物種大熊貓(Ailuropoda melanoleuca)的主食竹之一。該品種近似缺苞箭竹(F.denudata),稈叢生,葉片寬大,基部常不對稱,次脈常為4對,易于區分。團竹分布于北川、平武、松潘、茂縣4縣交界的高山地區,海拔2 500~3 300 m,并為該區垂直分布最高的竹種,其下依次為缺苞箭竹(F.denudata)、青川箭竹(F.rufa)、油竹子(F.angustissima)[1]。克隆生長是指自然條件下,生物通過營養方式自發形成遺傳上相同,并具有潛在獨立性的新個體或分株的能力或過程[2]。克隆繁殖使得克隆植物具備了較強的水平擴展能力,占據一個大的水平空間,并通過克隆分株的可塑性適應異質性環境[3-9]。植物個體大小與繁殖的關系是植物生活史理論研究的基本內容[6-7,10],有助于了解植物生態適應機制。團竹是多年生一次性開花結實的木質草本克隆植物,自1986年易同培于北川發現以來,對團竹的后續研究報道較少,目前尚無其生態習性、克隆生長構型與形態可塑性、繁殖及適應對策、資源調查等方面的報道。本研究以2004年片口自然保護區團竹開花死亡后新長成的團竹為對象,調查不同海拔環境中團竹的分株形態特征,探討海拔變化對團竹分株形態特征的影響,以及團竹母株個體大小對克隆分株、間隔子生長的影響,以揭示其克隆繁殖生態適應機制,為保護和培育團竹提供理論依據。
1 試驗地區與方法
1.1 試驗地區自然概況
北川片口自然保護區地處四川盆地西緣、青藏高原東南緣岷山中段、龍門山西側,位于北川縣片口、小壩、開坪3鄉境內,地理坐標為104°10′~104°26′E,32°02′~32°12′N,占地面積8 253 hm2。自然保護區內為北亞熱帶濕潤季風氣候,年均氣溫7.2~11.2 ℃,年均降雨量800 mm。區內土壤呈明顯的垂直帶譜分布,森林(含灌木林地)覆蓋率達997%,植被垂直地帶性分布明顯。該保護區是以保護大熊貓、金絲猴(Rhinopithecus roxellana)、扭角羚(Budorcas taxicolor)等珍稀野生動物及其自然生態系統為主的自然保護區,是岷山山系自然保護區的典型代表之一[11]。
1.2 調查方法
于2012年10月中旬,在保護區內皮棚子處海拔2 700~2 900 m的團竹種群中開展野外調查。其喬木層主要由岷江冷杉(Abies faxoniana)構成,伴生樹種主要有紅樺(Betula albo-sinensis)、糙皮樺(Betula utilis)等;灌木層為團竹、花楸(Sorbus pohuashanensis)、紫花杜鵑(Rhododendron amesiae)等;草本層植物稀少。區內團竹種群無明顯蟲害及大熊貓采食痕跡。
在海拔2 700~2 900 m范圍內,每隔100 m分別隨機設置3個10 m×10 m的樣地。在各樣地內隨機選擇20株(包括一級母株[12]、間隔子、當年生竹筍長成的立竹)完整團竹分株對,分別記錄母株和當年生立竹的高度和基徑、間隔子的長度和直徑、地下莖分蘗數(分枝強度)。用自封袋裝好帶回實驗室,洗盡泥沙后將母株、間隔子、當年生立竹進行分裝,于65 ℃恒溫烘干至恒質量,冷卻后使用萬分之一電子天平稱質量并記錄。
1.3 數據分析
采用SPSS 18.0軟件分析所有試驗數據;采用One-way ANOVA軟件分析不同海拔環境下團竹分株形態特征的差異性,并作LSD多重比較;采用Regression軟件分析克隆分株的生物量模型、母株和克隆生長參數的回歸關系,并檢驗顯著度。其中,比間隔子長為間隔子長與間隔子生物量的比值[7];間隔子分枝強度為地下莖分蘗數。
2 結果與分析
2.1 不同海拔團竹分株的個體特征
團竹分株個體形態特征中,母株高度、當年生立竹生物量和高度、間隔子直徑均隨海拔的升高而不斷增大,且差異顯著。比間隔子長隨海拔的升高呈先減小后增大的趨勢,且差異極顯著。間隔子分枝強度隨海拔的升高呈先增大后減小的趨勢,且差異極顯著。母株生物量和基徑、間隔子長度和生物量均無明顯差異(表1)。
2.2 不同海拔團竹分株的生物量模型
團竹母株生物量和基徑、當年生立竹生物量和基徑、間隔子生物量和直徑的回歸關系見表2。隨著海拔由低到高,母株生物量隨基徑的增大分別呈指數增長、三次曲線增長、線性增長趨勢,總體呈二次曲線增長趨勢,各增長均達極顯著水平;當年生立竹生物量隨基徑的增大分別呈對數增長、二次曲線增長、線性增長趨勢,總體呈指數增長趨勢,各增長均達極顯著水平;間隔子生物量和直徑分別呈指數增長、線性增長、指數增長趨勢,總體間隔子生物量隨直徑的增大呈指數增長趨勢,各增長均達極顯著水平。
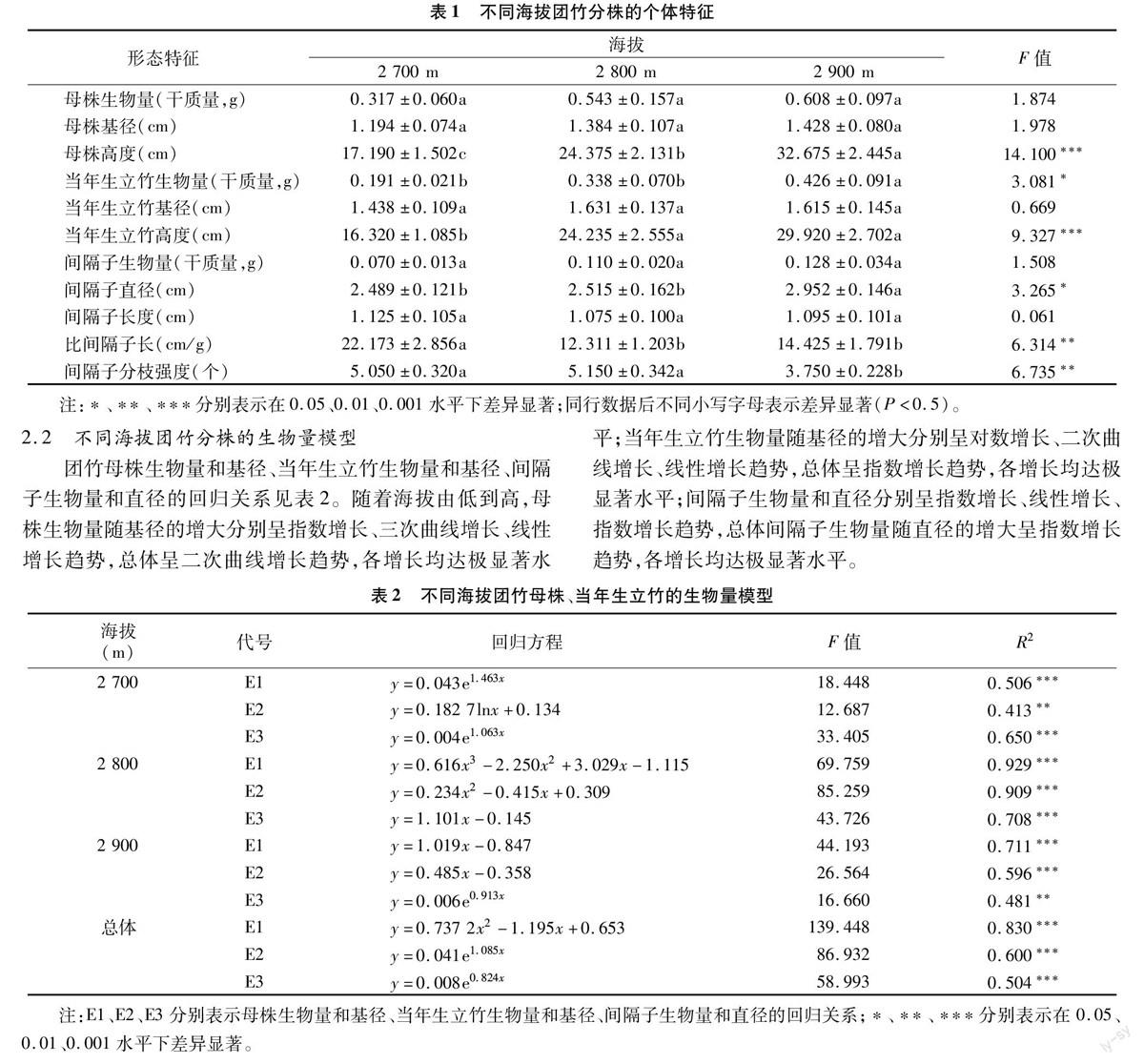
2.3 不同海拔母株與間隔子的回歸關系
在海拔2 700、2 900 m處,間隔子直徑隨母株基徑的增大均呈顯著的二項式增長;在海拔2 800 m處,間隔子直徑隨母株基徑的增大呈極顯著的線性增長。間隔子直徑隨母株生物量的增大呈極顯著的線性增長(圖1)。
2.4 不同海拔母株與當年生立竹的回歸關系
在海拔2 700 m處,當年生立竹基徑與母株基徑、母株生物量均呈顯著的線性增長;在海拔2 800 m處,當年生立竹基徑隨母株基徑的增長呈極顯著的線性增長,隨母株生物量的增長呈極顯著的對數增長;在海拔2 900 m處,當年生立竹基徑與母株基徑呈極顯著的指數增長,與母株生物量呈極顯著的線性增長(圖2)。
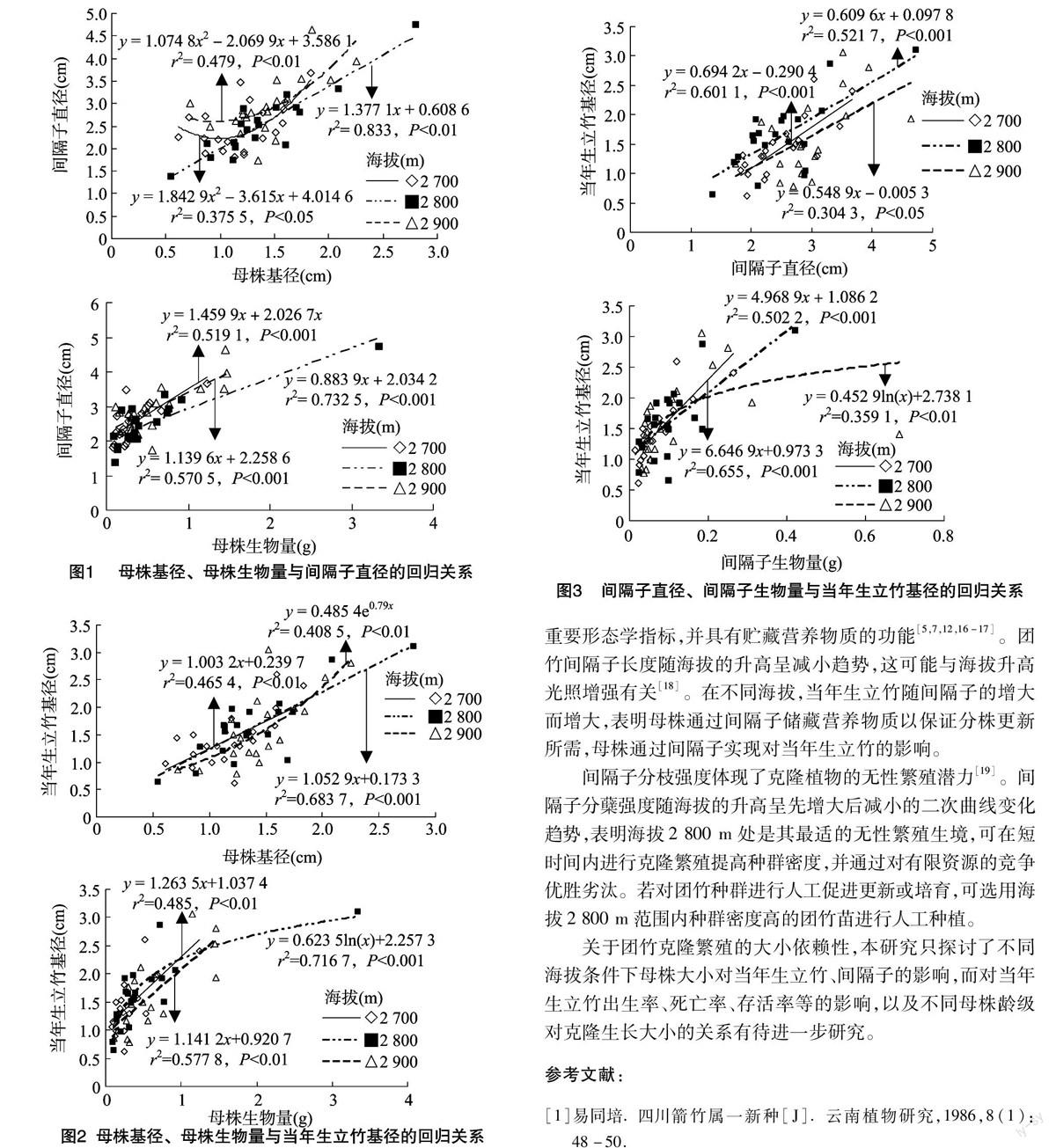
2.5 間隔子對當年生立竹的影響
在海拔2 700、2 800 m處,當年生立竹基徑隨間隔子直徑、間隔子生物量的增大均呈極顯著的線性增長;在海拔2 900 m處,當年生立竹基徑與間隔子直徑呈極顯著的線性增長,與間隔子生物量呈極顯著的對數增長(圖3)。
3 結論與討論
海拔梯度包含了溫度、水分、光照等環境因子的劇烈變化,因此成為研究植物環境適應性、對全球氣候變化響應的理想區域[13]。本研究中各海拔條件下,團竹母株的生物量、基徑、高度,當年生立竹的生物量、高度,間隔子的生物量、直徑均隨海拔的升高而不斷增大。潘紅麗等在對臥龍自然保護區華西箭竹的研究中發現,生物量隨海拔的升高呈先升高后降低的趨勢[13];劉興良等在研究中發現,生物量隨海拔的升高呈下降趨勢[14]。以上結論均與本研究結果不一致,這可能是由于各品種的最適生長海拔不同,是物種適應異質性環境的結果。
物種的繁殖需要消耗大量營養物質,植物個體大小是衡量植物對資源積累的重要指標之一[6-7,12]。以基徑和生物量衡量團竹母株大小,發現在不同海拔條件下,當年生立竹、間隔子大小均隨母株的增大而增大,呈顯著正相關,可見團竹的克隆生長在個體水平具有大小依賴關系,這與解蕊等、王瓊等、劉左軍等的研究結果[7,12,15]一致。
間隔子是研究克隆植物在資源異質性環境中覓食行為的
重要形態學指標,并具有貯藏營養物質的功能[5,7,12,16-17]。團竹間隔子長度隨海拔的升高呈減小趨勢,這可能與海拔升高光照增強有關[18]。在不同海拔,當年生立竹隨間隔子的增大而增大,表明母株通過間隔子儲藏營養物質以保證分株更新所需,母株通過間隔子實現對當年生立竹的影響。
間隔子分枝強度體現了克隆植物的無性繁殖潛力[19]。間隔子分蘗強度隨海拔的升高呈先增大后減小的二次曲線變化趨勢,表明海拔2 800 m處是其最適的無性繁殖生境,可在短時間內進行克隆繁殖提高種群密度,并通過對有限資源的競爭優勝劣汰。若對團竹種群進行人工促進更新或培育,可選用海拔2 800 m范圍內種群密度高的團竹苗進行人工種植。
關于團竹克隆繁殖的大小依賴性,本研究只探討了不同海拔條件下母株大小對當年生立竹、間隔子的影響,而對當年生立竹出生率、死亡率、存活率等的影響,以及不同母株齡級對克隆生長大小的關系有待進一步研究。
參考文獻:
[1]易同培. 四川箭竹屬一新種[J]. 云南植物研究,1986,8(1):48-50.
[2]董 鳴,于飛海. 克隆植物生態學術語和概念[J]. 植物生態學報,2007,31(4):689-694.
[3]Cook R E. Growth and development in clonal plant populations[M]. New haven:yale university press,1985:259-296.
[4]董 鳴. 異質性生境中的植物克隆生長:風險分攤[J]. 植物生態學報,1996,20(6):543-548.
[5]宋利霞,陶建平,冉春燕,等. 臥龍亞高山暗針葉林不同林冠環境下華西箭竹的克隆生長[J]. 植物生態學報,2007,31(4):637-644.
[6]解 蕊. 亞高山不同針葉林冠下大熊貓主食竹的克隆生長[D]. 北京:北京林業大學,2009:3-34.
[7]解 蕊,李俊清. 亞高山針葉林冠下缺苞箭竹的克隆生長[J]. 東北林業大學學報,2009,37(8):22-25.
[8]陶建平,宋利霞. 亞高山暗針葉林不同林冠環境下華西箭竹的克隆可塑性[J]. 生態學報,2006,26(12):4019-4026.
[9]Doust L L. Population dynamics and local specialization in a clonal perennial(Ranunculus repens).Ⅲ.Responses to light and nutrient supply[J]. Journal of Ecology,1987,75(2):555-568.
[10]Klinkhamer P L,Meelis E,Jong T J,et al. On the analysis of size-dependent reproductive output in plants[J]. Functional Ecology,1992,6(3):308-316.
[11]岳 鵬,周 勇. 四川片口自然保護區總體規劃[R]. 成都:四川省林業廳,四川省林業勘察設計研究院,2005:1-75.
[12]王 瓊,蘇智先,雷濘菲,等. 慈竹母株大小對克隆生長的影響[J]. 植物生態學報,2005,29(1):116-121.
[13]潘紅麗,田 雨,劉興良,等. 臥龍自然保護區華西箭竹(Fargesia nitida)生態學特征隨海拔梯度的變化[J]. 生態環境學報,2010,19(12):2832-2839.
[14]劉興良,劉世榮,宿以明,等. 巴郎山川滇高山櫟灌叢地上生物量及其對海拔梯度的響應[J]. 林業科學,2006,42(2):1-7.
[15]劉左軍,杜國禎,陳家寬. 不同生境下黃帚橐吾(Ligularia virgaurea)個體大小依賴的繁殖分配[J]. 植物生態學報,2002,26(1):44-50.
[16]Kleiju D,Van G J M. The exploitation of heterogeneity by a clonal plant in habitats with contrasting productivity levels[J]. Journal of Ecology,1999,87:873-884.
[17]Dong M,De K H. Plasticity in morphology and biomass allocation in cynodon dactylon,a grass species forming stolons and rhizomes[J]. Oikos,1994,70:99-106.
[18]羅學剛,董 鳴. 蛇莓克隆構型對光照強度的可塑性反應[J]. 植物生態學報,2001,25(4):494-497.
[19]吳福忠,王開運,楊萬勤,等. 大熊貓主食竹群落系統生態學過程研究進展[J]. 世界科技研究與發展,2005,27(3):79-84.
