棉花對鹽脅迫的響應機制及緩解措施的研究進展
2015-04-29 00:44:03劉祎崔淑芳張海娜錢玉源王廣恩金衛平李俊蘭
農學學報 2015年11期
劉祎 崔淑芳 張海娜 錢玉源 王廣恩 金衛平 李俊蘭

摘要:鹽分是阻礙作物生長發育的重要逆境之一。歸納了鹽脅迫對棉花生長發育、產量和品質的影響;分析了棉花適應鹽逆境、減輕傷害的自我調節和響應機制,包括滲透調節、膜脂調節、離子分布和分子機制;總結了耐鹽品種選育、栽培管理技術和分子育種等緩解棉花鹽脅迫的主要措施;最后提出棉花耐鹽相關研究的方向:分子機理與生理機制相結合;常規栽培措施與理化手段相結合;遙感監測系統的開發與應用。
關鍵詞:棉花;鹽脅迫;響應機制;緩解措施
中圖分類號:S562 文獻標志碼:A 論文編號:cjas15040008
0引言
鹽漬地在世界各地分布很廣,主要分布在澳大利亞、俄羅斯、中國、印度尼西亞、巴基斯坦等國家。土壤鹽漬化是世界農業可持續發展的重要限制因素,并隨著人類發展不斷地加重擴大,已成為重要的環境問題之一。隨著人口不斷增長,耕地逐漸減少,糧食安全問題倍受重視,糧棉爭地矛盾日益顯現,鹽堿地植棉在全國棉花生產體系中所占份額越來越大,地位越來越重要。根據第二次全國土地調查資料顯示,中國鹽漬地面積約5.25億畝,其中約5000萬畝分布于棉花優勢產區。因此,深入研究其耐鹽堿性,提高現有鹽堿地棉田產量,逐步推進開發鹽堿地植棉,對解決棉糧爭地、改善和利用鹽堿地等問題具有重要意義。棉花是耐鹽的先鋒作物,但耐鹽能力有限,其種間耐鹽性也有較大差異。鹽脅迫對棉花生長發育乃至產量和品質的影響程度,不僅與棉花的生長發育特性、環境變化等因素有關,也取決于棉花對鹽脅迫的適應性。目前,棉花耐鹽的相關研究多集中在鹽脅迫下的生理損傷、耐鹽遺傳機理和分子機理等方面。本研究就棉花對鹽脅迫的響應機制及緩解的主要措施進行闡述,以期為今后相關的研究提供參考依據。
1鹽脅迫對棉花生長發育和產量品質的影響
1.1鹽脅迫對棉花生長發育的影響
通常,鹽脅迫阻礙棉花根部和地上部的生長,導致側根生長受到抑制,葉面皺縮且面積減少,莖部纖弱,地上和地下部的鮮物質量下降,其中,地上部尤為敏感。高鹽脅迫會對光合、呼吸作用產生嚴重影響,阻礙氮、磷肥吸收利用。隨著鹽濃度增加,花鈴數減少,落鈴率增加,從而導致成鈴減少。不同鹽濃度對棉花生長發育的不同時期、不同器官都有影響。當0.2%<鹽濃度<0.4%時,能促進種子萌發;當0.5%<鹽濃度<0.7%時,生理機能、物質代謝受到影響,幼苗生長受限;當鹽濃度>0.7%時,則顯著抑制棉花的生長發育,導致產量下降、品質降低。鹽脅迫使Na+大量積累,無機離子缺乏限制了水分,導致棉花營養失衡。在0.4%鹽土壤環境中,耐鹽品種出苗的耐鹽脅迫能力強于感鹽品種,并隨播種時間的推遲,抑制作用越明顯。生長過程中,鹽脅迫影響葉綠素合成,光合作用受抑制,葉片中葉綠素含量和光合速率下降,Na+、脫落酸(ABA)含量上升,K+、玉米素核苷(ZR)含量降低,促進葉片的衰老,耐鹽性好的品系葉綠素相對值降低幅度較小。較低NaCl濃度對棉花愈傷組織生長有促進作用,而高NaCl濃度對棉花愈傷組織有明顯的抑制作用。
1.2鹽脅迫對棉花產量和品質的影響
棉花干物質生產受土壤鹽分影響顯著,高鹽度條件下棉花生育進程滯后,生殖生長與營養生長不協調,造成脫落率增加,經濟產量下降。土壤鹽度顯著影響棉花對N、P和K養分的吸收、積累、分配,在子棉、鈴殼中的吸收量與鹽度呈負相關。棉花成苗率,株高,果枝數,單鈴重,單株最大葉面積,霜前花率,整齊度指數和斷裂比強度均隨著灌溉水鹽度的增加而降低,并且通過馬克隆值對纖維品質產生負面影響。棉花耐鹽性與鈴重、纖維長度呈極顯著正相關;與種仁蛋白質含量、馬克隆值呈顯著負相關,鹽脅迫后,蛋白質含量均降低。適當范圍濃度的鹽可作為營養成分,提高種子產量,但是Na+的大量攝入會導致葉片早熟或早衰,產種量驟減,不同基因型的棉纖維性狀受到的影響也是多種多樣。
2棉花對鹽脅迫的響應機制
作物的耐鹽特性不僅與植株體內鹽分的吸收、運輸、分配、排泄、膜功能和離子區域化作用相關,耐鹽物質的合成和積累也是提高耐鹽性的重要途徑。棉花是非鹽生植物,但是能夠像鹽生植物一樣,通過不同的耐鹽機制來抗拒鹽脅迫,主要包括滲透調節、膜脂調節、離子分布和分子機制。
2.1滲透調節
受到鹽脅迫時,外界滲透勢低,植物體內細胞出現水分虧缺現象,即滲透脅迫,為保證水分的正常供應,細胞會積累某些可溶性物質來避免傷害。鹽脅迫造成的滲透脅迫容易引起細胞脫水,棉株通過吸收大量無機鹽,并將鹽離子積累到液泡,以提高液泡溶液濃度,從而降低細胞水勢,以適應鹽脅迫造成的低水勢,進而使植株在較高滲透條件下吸水,保持膨壓,提高耐脫水性。但是,過多鹽離子的積累會產生毒害作用。
2.2膜脂調節
鹽脅迫下,植物細胞內Na+過量積累,導致活性氧產生,打破了清除系統的動態平衡,啟動膜脂過氧化及脫脂作用,致使膜蛋白和膜脂損失,膜結構破壞,此時超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、過氧化物酶(POD)、谷胱甘肽還原酶(GR)和抗壞血酸(AsA)等活性在一定程度上發生變化,以維持膜結構和功能的穩定。人們通過對鹽脅迫下植物體內抗氧化防御系統的大量研究,發現SOD、POD、AsA等清除活性氧的酶系和抗氧化物質協同作用共同抵抗鹽脅迫誘導產生的氧化傷害,單一的抗氧化酶不足以防御這種氧化脅迫。受到鹽脅迫,耐鹽性棉花品種葉片的膜脂過氧化清除系統活性(包括POD、CAT、GR)大幅度上升,不耐鹽型品種的指標不變或下降。楊艷兵等用0.3%NaCl脅迫處理中熟棉‘N181,幼苗葉片中保護酶活性及脯氨酸含量明顯增加,丙二醛含量降低。Askim等提出,脅迫使CAT、POD活性降低,導致過氧化氫積聚和氧化應激引發脂質過氧化反應。耐鹽型品種,可以保持SOD和AsA活性,同時脯氨酸含量會大大提高。棉花可以通過體內合成某些小分子有機物(例如:脯氨酸、葡萄糖和其它氨基酸等)、新的應激蛋白(或某種蛋白量的改變)來提高細胞的滲透壓,以降低水勢,以適應環境的變化。還可以通過降解體內蛋白質、淀粉,來提高滲透壓,改變代謝過程,同時,有大量脯氨酸、可溶性糖的合成維持滲透壓。不同耐鹽級別品種的細胞質膜透性差異顯著,耐鹽性越強其葉片細胞質膜透性越小。耐鹽性強的品種,苗期葉片內游離脯氨酸、可溶性糖、類胡蘿卜素的含量,棉苗的過氧化物酶活性、葉綠素a、b、a+b含量及凈光合速率均顯著高于耐鹽性弱的品種,丙二醛含量顯著低于耐鹽性弱的品種。
2.3離子分布
耐鹽生理中,碳水化合物是主要的滲透調節劑,參與平衡液泡水勢。有機酸同樣是一類重要的有機物,可以作為一種滲透調節劑,還可以用來維持植物體內的離子平衡,通過中和過量Na+、K+、Ca2+、Mg2+等陽離子,一定程度上降低了陽離子毒害。Flowers等研究表明,由于鹽生和非鹽生植物的細胞質酶對Na+的敏感程度不同,因此Na+、CI的離子區域化分布是鹽漬環境的適應結果。離子的區域化分布,液泡中Na+的濃度大于胞質中的濃度,鹽脅迫時,Na+、Cl-等離子積累在液泡中。在鹽脅迫下棉花植株體內的ca2+、Na+、Cr含量隨土壤鹽度的增加而增加,根莖木質部及葉片的Na+含量隨外界鹽濃度的增大而迅速上升,尤以葉片中含量最高。耐鹽型品種,體內的陽離子以Na+為主,并且有約90%積累在地上部。NaCl脅迫顯著降低棉苗葉片和根系中Ca2+、K+、Mg2+、P3+和Mn4+的含量。耐鹽性強的品種苗期葉片內K+含量顯著高于耐鹽性弱的品種,Na+含量顯著低于耐鹽性弱的品種。
2.4分子機制
鹽脅迫下,質膜和液泡膜的ATP酶/H+泵產生逆向轉運質子驅動,通過Na+/H+逆向轉運蛋白將細胞質中的Na+泵入液泡,緩解鹽脅迫造成的危害。棉花的耐鹽性強弱因基因型而異,基因型間的耐鹽性差異與丙二醛、脯氨酸、可溶性糖、類胡蘿卜素含量和保護酶活性以及鹽離子在葉片中積累量的差異有關㈣。棉花是較耐鹽植物,V-PPase(焦磷酸酶)在鹽脅迫下起到離子區隔化的作用,而植物的逆境脅迫與DNA甲基化有重要關系。在鹽脅迫下,DNA甲基化變異在根中發生的更為頻繁,幾乎以甲基化降低為主。李春賀利用miRNA基因芯片雜交技術,miRNA家族作用的靶基因分析,miRNAs參與植物逆境條件下的信號傳導,很可能對棉花的耐鹽機制有重要作用。何林池等發現親緣關系很近而耐鹽水平差異顯著的‘中棉所35和‘中棉所12存在點突變和插入/缺失,說明這些SNP/InDel可能對耐鹽性狀有重要的調控功能。通過KEGG分析,耐鹽品系顯著富集的信號通路是抗原加工過程,而鹽敏感品系顯著富集的是生物堿合成途徑㈣。另外,LEA蛋白基因、水通道蛋白基因、K+通道蛋白及其基因、鹽激蛋白及其基因等,也在棉花耐鹽響應機制中發揮重要作用。
3緩解鹽脅迫的主要措施
3.1耐鹽品種的鑒定篩選及培育
3.1.1鑒定篩選耐鹽種質資源的評價和篩選也是耐鹽育種的基礎和前提條件。目前,已知的耐鹽性檢測方法有鹽池鑒定法、發芽率和出苗率估測法、生物學性狀比較法、離子比值法、整體植株鑒定法、花粉萌發率測定法、多標記組合鑒定法等。
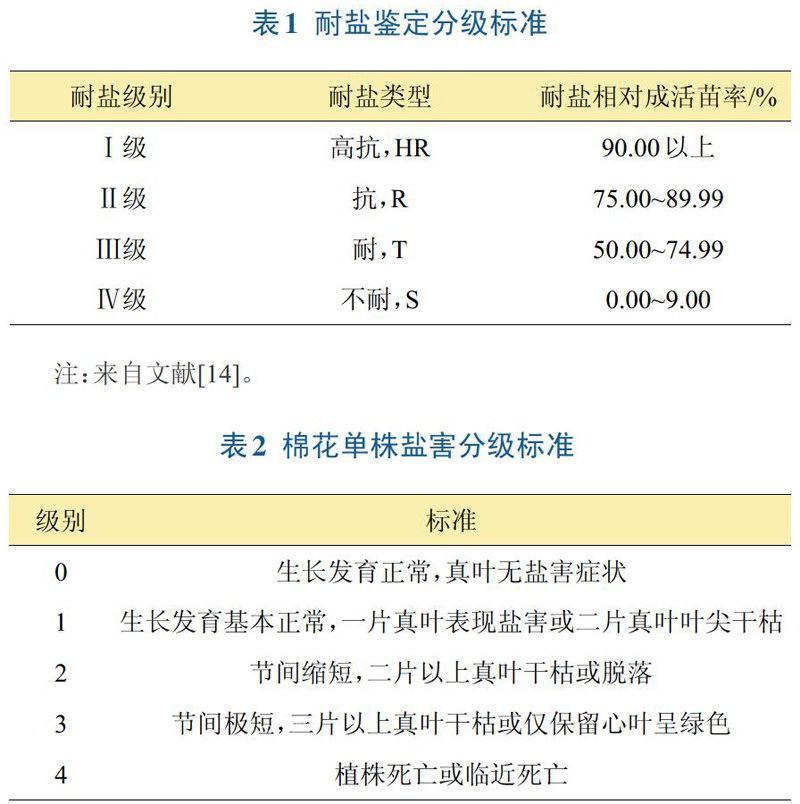
鹽池鑒定法是比較傳統的耐鹽性鑒定方法。葉武威將土壤基質含鹽量0.4%作為棉花耐鹽性鑒定的濃度,統計施鹽10天后的相對成活苗率(salinity-resistance index,SRI%),將耐鹽性分為4級(表1)。劉雅輝等采用鹽池原土鑒定法,在土壤全鹽含量0.4%和0.1%(對照)脅迫下,通過調查幼苗長勢、鹽害癥狀、二葉期的株高和葉片數,對棉苗分類并計算鹽害指數,確立了以鹽害指數為分級指標的棉花苗期耐鹽性評價標準。沈法富等以大田單株棉花鹽害癥狀為耐鹽性的分級標準(表2)。幼苗期是鑒定棉花耐鹽性的重要階段,種子相對發芽率、相對棉苗干物質質量、相對株高、K+/Na+和Ca2+/Na+等相對指標可用來輔助鑒定棉花萌發期耐鹽性。利用水培的方法對其整體植株進行耐鹽性鑒定,棉花葉片總面積和葉片鮮重減少百分率可以反映棉株整體的耐鹽性。利用液滴培養技術對花粉耐鹽性進行鑒定,棉花花粉粒萌發的百分率也是反映其耐鹽性的可靠指標。隨著生物技術不斷進步,分子標記的出現給耐鹽性研究帶來新思路、新方法。張麗娜等采用SSR技術,初步制定出多標記組合鑒定法,用于棉花耐鹽分子標記輔助鑒定。
3.1.2耐鹽品種選育全國以中國農業科學院棉花研究所為主的抗逆性鑒定中心,培育出一批常規抗旱、耐鹽新品種(系),并成功地在中國旱區、鹽堿地進行推廣種植。沈法富等利用Hayman的方法進行試驗,表明耐鹽和敏鹽品種的一般配合力效應差異達極顯著,耐鹽×敏鹽為棉花耐鹽育種的最佳組合。棉花的耐鹽性存在著加性和顯性效應,以加性效應為主,耐鹽性呈不完全顯性,受一對主效基因控制。結合F2代的狹義遺傳力效應,可以在F2選擇改良棉花的耐鹽性。辛承松等以雙價抗蟲棉品種‘SGK321和普通陸地棉品種‘石遠321(SY321)為材料,通過試驗表明選用耐鹽性強的親本并注重耐鹽性選擇是培育耐鹽抗蟲棉品種的有效途徑。
3.2栽培管理技術
通過播前種子處理、地膜覆蓋、灌溉耕作和施加外源物質等方法,對提高棉花耐鹽性均有一定的改良效果。
3.2.1種子處理播前對種子進行選擇和處理,在一定程度上可以提高棉花耐鹽性。研究表明,播前將種子在不同鹽濃度溶液中處理,可提高棉花抗鹽性。在鹽分脅迫下,利用適當濃度范圍的縮節胺、腐胺、2-氯-6-(三氯甲基)吡啶、CaCl2浸種可以提高棉種發芽率、發芽勢,降低平均發芽時間。部分種類及濃度的氨基酸能緩解鹽脅迫對棉籽萌發及棉花幼苗生長的抑制作用,而對脯氨酸的生成有不同程度的抑制作用。
3.2.2耕作方式覆膜可有效的改善植株生長,提高耐鹽性,增加皮棉產量。董合忠等提出在黃河三角洲及其生態相似的棉花產區,播前30天覆地膜可以提高土壤溫度,延遲鹽分積累,有效增加產量。將雙膜和精量播種技術緊密聯系,可以顯著改善出苗情況,以及低鹽和低溫等不利因素。與平作種植方式相比,上蓋地膜下埋秸稈、高壟覆膜,促進長勢,有利于提高棉花干物質質量和氮、磷、鉀的積累量。土壤鹽分隨灌溉水鹽度的增加而增高,適宜鹽度和灌溉量的微咸水滴灌對棉花生長、產量以及水氮利用率影響不大,高鹽度的咸水灌溉會導致棉花產量顯著降低。Wang等在新疆南部進行田間試驗表明,生長期進行膜下滴灌用淡水和淡鹽水,收獲后用淡水漫灌可以避免土壤鹽漬化。周麗等以典型荒漠區原生鹽堿荒地及開墾農田為研究對象,長期的耕作可以改變土壤可溶性鹽分在土層中的分布,經過多年開墾利用后,鹽荒地土壤可溶性鹽分和pH均顯著降低。
3.2.3外源施加施加氮肥、磷肥,可以改善棉株的耐鹽性。施加酸堿平衡護理劑、鹽堿土改良劑,可以調節酸堿平衡,改善土壤團粒結構,提高土壤肥力,對鹽堿地減輕鹽堿危害有明顯效果,增產效果明顯。采用黃腐酸作為化學改良劑改良膜下滴灌鹽堿地后,土壤pH、易溶鹽總量、SO42-、HCO3-、Mg2+、Cl-、Ca2+、K+、Na+均呈明顯降低趨勢,棉花長勢、產量等均優于試驗區外對照。使用硫磺粉進行改良鹽堿地的研究也是基于硫桿菌的作用,可以降低土壤的pH,改善養分狀況,促進了棉花植株的生長。在鹽堿地定植膠囊KlebsiellaoxytocaRs-5、施用假單細胞菌Pseudomonas putidaRs-198http://www.sciencedirect.com/science/arficle/pii/S1164556309001046-corl#corl、拉烏爾菌RaoultellaplanticolaRs_2提高棉花發芽勢和苗期耐鹽性。低濃度的冠菌素和縮節安處理減少植株內活性氧的含量同時增加植物光合能力以減輕棉花在鹽脅迫下的傷害。適當的SNP可以促進棉花生長,提升鹽脅迫下抗氧化防御系統的能力。增施磷肥也可以提高棉花的抗鹽性。
3.3分子輔助育種
隨著基因與克隆技術的發展,棉花生長發育機理的進一步明確,轉基因種質資源創新體系的建立和完善,將會發掘更多的耐鹽基因,并有更多基因轉移到棉花中,不斷的培育綜合性狀優良的轉基因棉花新品種(系)。
3.3.1棉花中耐鹽基因的發掘劉章偉等運用cDNA-AFLP技術,獲得了僅在旱地棉鹽脅迫下特異表達的轉錄片段(TDF),這些鹽誘導表達的基因主要涉及離子轉運、活性氧清除、細胞信號傳導、細胞分裂、轉錄調節、膜保護、滲透調節等功能蛋白。利用RACE結合RT-PCR技術獲得陸地棉s一腺苷甲硫氨酸合成酶基因(GhSAMS),該基因的表達受鹽脅迫誘導,在鹽敏感材料中誘導被推遲。從棉花中克隆出蘋果酸脫氫酶(Malatedehydrogenase,MDH)基因(GhMDH),該基因編碼的蛋白酶具有脫氫酶活性。膜聯蛋白基因GhAnnl、GhWRKY39的過表達,可提高棉花抗鹽性。GhBI-1基因能夠響應生物與非生物脅迫信號的誘導。陸地棉中親環蛋白基因GhCyp1是耐鹽脅迫的基因。目前已在棉花中發現的一些耐鹽相關基因,有受高鹽誘導的基因(GhGnT、GhCPK5、GhMPK2、GhCpyl、GhVP、GhDREB、GhPTACGhBI-1等),調控耐鹽相關基因表達基因(GhEREB2、GhDBP1、GhDREB1、GhHB1、GhERF1、GhZFP1等),受ABA誘導的基因(GhMT3a、GhDREB1L、GhNAC1、GhDi19-1、GhCBF1、GhSRK2D等),調節離子運輸,維持滲透壓,促進水分吸收的基因(Lea、GhNHX1、GhTIP1、GhCNGC2、GaP5CS、GaTPS等),參與信號傳導途徑,清除活性氧的基因(GhPK1、Cu/ZnSOD等)等。
3.3.2轉基因技術的應用利用轉基因技術,將耐鹽堿相關基因轉入到棉花中,是增強棉花耐鹽堿性的重要手段。在棉花品種‘YZ1中過表達水稻的SNAC1基因后,250mmol/LNaCl條件下水培,其根系發育生長旺盛,蒸騰速率降低,脯氨酸含量上升,丙二醛含量下降,其耐鹽堿性得到了提高。棉花中轉入來自濱藜屬的nhaA基因,可以改善Na+和K+積累。試驗表明,將山菠菜的AhC-MO基因、膽堿脫氫酶基因beta和突變的乙酰乳酸合成酶als基因等外源基因(表3)導入到棉花中,可以提高轉基因植株的耐鹽和耐低溫能力。
4展望
長期以來,如何提高棉花的抗鹽性,增加在鹽逆境下的產量一直是人們關注的焦點。目前,對棉花的抗鹽機制研究有限,更加深入系統的了解棉花抗鹽機制,探明棉花對鹽堿脅迫的反應機制和抗鹽機理,是指導生物技術和其它措施改造棉花提高抗鹽堿能力的前提。當今,學科間交叉網絡的發展,給棉花的耐鹽工作提出了新的思考和契機。
(1)分子機理與生理機制相結合。棉花耐鹽性是一個復雜的數量性狀,由多基因控制。隨著轉基因技術的不斷深入和發展,部分轉基因棉花獲得了較好的經濟和社會效益,但是耐鹽轉基因棉花研究方面,仍然存在著亟待解決的問題,例如:耐鹽相關基因轉化率過低,基因表達對耐鹽能力改善的局限性,如何感知環境脅迫信號,并且如何進行調控等。可以通過對生理機制、分子機理和育種的結合應用,篩選、培育耐鹽品種。李維江等研究表明,耐鹽性因基因型而異,棕色棉和綠色棉耐鹽性最強,因此,可以結合生產,選育實用性強的品種。
(2)常規栽培措施與理化手段相結合。通過抗鹽鍛煉、使用生長調節劑和改造鹽堿土等傳統栽培措施是提高棉花的抗鹽能力的有效手段。研究表明,磁處理可以通過影響鹽脅迫后植物細胞內離子轉移和分布變化,進而引起與之相關的代謝酶類、生理活性發生改變,嘗試提高鹽漬環境下棉花種子的萌發率。
(3)遙感監測系統的開發與應用。林蔚等運用軟件工程的思維,耦合了基于地面遙感手段構建的棉花功能葉水、鹽狀況監測模型和土壤介電常數模型,開發了具有監測濱海鹽土棉田棉花和土壤水、鹽狀況功能的遙感監測系統。李靜等通過對棉田土壤鹽分的光譜反演研究,選擇鹽漬化光譜敏感的波段建立了鹽分指數SI1,BI,SI2,NDSI,SI3監測棉田土壤鹽漬化的模型,為土壤鹽漬化遙感動態監測提供可能。
