滇龍膽GrWRKY2基因的克隆與序列分析*
2015-03-27 05:52:24
玉溪師范學院學報 2015年4期
(玉溪師范學院 資源環境學院,云南 玉溪 653100)
滇龍膽(Gentianarigescens)為云南道地藥材,其主要藥效成分為龍膽苦苷.滇龍膽是龍膽瀉肝片、苦膽草片等200多種中藥的主要成分[1].近年來,龍膽市場需求量劇增,使野生滇龍膽野生資源遭到極大破壞[1].要解決龍膽的藥源問題,必須弄清龍膽苦苷生物合成途徑及其調控機理,這樣才能為通過基因工程生產龍膽苦苷奠定基礎.
研究表明,WRKY轉錄因子能夠調控萜類的生物合成.如,在棉花中,GaWRKY1通過激活(+)-δ-杜松烯合成酶而促進棉子酚的生物合成[2];在擬南芥中,AaWRKY1通過激活紫穗槐-4,11-二烯合成酶的合成進而促進青蒿素的生物合成[3];在番茄毛狀體中,SlWRKY73主要在根中表達,能夠激活萜類合成酶基因SlTPS5的表達[4];在西洋參中,PqWRKY1是三萜人參皂苷生物合成的正調控因子[5].此外,最新研究表明,擬南芥中30%(22/72)的WRKY轉錄因子都對茉莉酸甲酯處理產生響應,而在長春花中該數字為25%(12/48)[6].此前,通過對滇龍膽的根和葉進行轉錄組測序分析,筆者發現GrWRKY2基因在葉中強烈上調,因此該基因可能與龍膽苦苷生物合成途徑的調控有關.
此外,從目前的研究情況看,國內外學者對龍膽的研究主要集中在種子萌發[7,8]、DNA條碼[9,10]、轉錄因子功能[11,12]、育種[13]等方面,而尚未對滇龍膽GrWRKY2基因進行克隆和表達分析.因此,在本研究中,筆者根據滇龍膽轉錄組中的GrWRKY2基因序列,設計一對特異性引物,通過RT-PCR技術成功從滇龍膽幼葉中擴增到該基因,并對其進行序列分析.
1 材料和方法
1.1 材 料
本實驗所用材料為滇龍膽(Gentianarigescens),其植株栽培于玉溪師范學院分子生物學實驗室.RNA提取所用材料為滇龍膽無菌苗幼葉.
1.2 方法
(1)葉片總RNA提取及GrWRKY2基因ORF的克隆.按照多糖植物組織提取試劑RNAiso(Takara,寶生物工程(大連)有限公司)說明書提取滇龍膽幼葉總RNA,按照逆轉錄試劑盒(Takara,寶生物工程(大連)有限公司)說明書合成cDNA.根據中間表達載體pENTR2B多克隆酶切位點和滇龍膽轉錄組中GrWRKY2基因序列,設計一對特異引物GrWRKY2BamHI-F:GGATCCATGTCTGACTCTAATTTTTATCAC(下劃線為BamHI酶切位點),GrWRKY2XhoI-R:CTCGAGTCAAATACTAGGGTTTGGATT(下劃線為XhoI酶切位點,引物由上海捷瑞生物工程有限公司合成).以cDNA為模板進行PCR擴增,反應體系為PrimeSTAR Max Premix(2×,Takara,寶生物工程(大連)有限公司)25 μl,cDNA模板2 μl,正反向引物(10 μmol/L)各1 μl,加ddH2O補足50 μl.PCR反應條件為:98 ℃變性10 s,50 ℃退火15 s,72 ℃延伸5 s,30循環;72 ℃加A尾30 min.PCR產物經1.0%瓊脂糖凝膠電泳分離,然后割膠,使用膠回收試劑盒(Qiagen,德國)對目的片段進行回收,將其連接到pMD19-T載體(Takara,寶生物工程(大連)有限公司).轉化大腸桿菌DH5α(Takara,寶生物工程(大連)有限公司)后進行藍白篩選,挑取白斑搖菌;使用試劑盒提取質粒(百泰克,北京),經酶切檢測正確后進行測序(上海生工,上海),獲得重組質粒pMD19-GrWRKY2.
(2)GrWRKY2基因的生物信息學分析.參考張曉東等[14]的方法對GrWRKY2基因進行生物信息學分析,其中二級結構分析網址為:http://scratch.proteomics.ics.uci.edu/index.html.
2 結果與分析
2.1 滇龍膽GrWRKY2基因序列的克隆
以滇龍膽幼葉cDNA為模板,使用特異性引物擴增出約400 bp的片段.通過TA克隆獲得重組質粒pMD19-GrWRKY2,酶切檢測結果表明雙酶切獲得的兩片段大小之和等于單酶切片段大小,與預期相符.
2.2 GrWRKY2基因的生物信息學分析
利用Genetyx、DNAMAN軟件對GrWRKY2基因cDNA序列進行分析,結果顯示GrWRKY2基因ORF全長362 bp,編碼120個氨基酸.將該序列上傳至GenBank數據庫,獲得登錄號KM215137.
利用DNAMAN 7軟件將GrWRKY2蛋白氨基酸序列與NCBI中相似性較高的部分序列進行比對分析,結果表明GrWRKY2蛋白與已知蛋白序列保守性較高(圖1).利用MEGA6.06將GrWRKY2氨基酸序列與其他相似性較高的WRKY序列進行系統發育分析,結果顯示滇龍膽GrWRKY2蛋白與琴葉擬南芥AlWRKY15和白菜BrWRKY51親緣關系較近(圖2).
使用ExPASy ProtParam tool對GrWRKY2蛋白進行分析,結果表明:GrWRKY2蛋白單體相對分子質量為13.24 kD,pI為8.29;帶正電氨基酸殘基(Arg+Lys)為13,帶負電氨基酸殘基(Asp+Glu)為11,其分子式為C567H907N161O188S8,不穩定指數為38.20,屬于穩定蛋白;脂肪指數為71.58,總平均疏水性(GRAVY)為-0.518,為親水蛋白.分析結果表明還表明,GrWRKY2蛋白含有20種基本氨基酸,其中絲氨酸含量最高,為14.2%,其次是天冬酰胺,為12.5%,色氨酸含量最低,為0.8%.

說明:黑色-相似性等于100%;深灰色-75%≤相似性<100%;淺灰色-50%≤相似性<75%圖1 GrWRKY2蛋白與其他植物WRKY蛋白保守區的比對結果
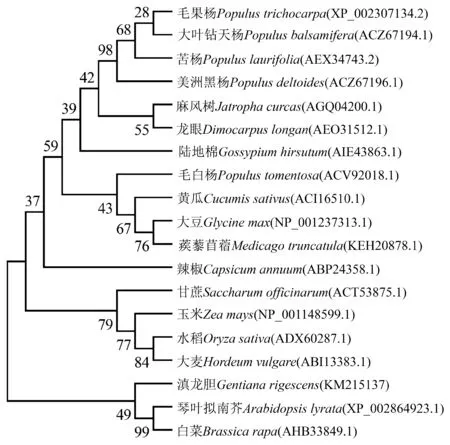
圖2 GrWRKY2蛋白與植物中其他WRKY2蛋白的系統發育分析

圖3 GrWRKY2蛋白的三維結構預測
利用Sspro對GrWRKY2蛋白進行二級結構分析,結果表明:該蛋白二級結構中α-螺旋(H)占28.33%,無規則卷曲(C)占59.17%,延伸帶(E)占12.50%.利用Swiss-Model Workspace,使用自動模式預測GrWRKY2蛋白的三級結構,結果如圖3.該模型以擬南芥AtWRKY4[1wj2.1]為模板,在第89~120氨基酸處建模,序列相似度為53.13%.使用InterPro在線工具對GrWRKY2蛋白的保守結構域進行分析,結果表明GrWRKY2蛋白包含DNA-binding WRKY(IPR003657)保守結構域.
利用SignalP 4.1服務器分析GrWRKY2蛋白,未發現信號肽,表明該蛋白為非分泌型蛋白.利用TMHMM工具預測GrWRKY2蛋白的跨膜螺旋區,結果表明GrWRKY2蛋白不含跨膜螺旋區域,為非膜蛋白.使用WoLF PSORT軟件進行亞細胞定位分析,結果為GrWRKY2蛋白的定位系數分別為:細胞核5.0,葉綠體3.0,細胞質3.0,分泌途徑2.0.
3 討 論
WRKY蛋白家族的作用比較廣泛,主要參與生物脅迫、非生物脅迫、種子發育、種子休眠與萌發、衰老等過程[15].最近的研究結果還表明,WRKY蛋白還參與萜類生物合成的調控[2,3,5].WRKY蛋白的最典型特征是包含“WRKY”結構域,其中W=色氨酸、R=精氨酸;K=賴氨酸;Y=酪氨酸.根據該保守結構域,筆者在滇龍膽數據庫中進行搜索,發現了大量的WRKY蛋白,其中GrWRKY2基因在葉中表達量遠遠高于根,可能參與龍膽苦苷的生物合成.為此,本文對GrWRKY2基因進行克隆和生物信息學分析.但在基因克隆過程中,由于受到DNA聚合酶種類、PCR循環數等影響,往往會導致所克隆的片段發生突變.因此,本研究采用目前保真性最高的酶PrimeSTAR Max Premix,并將PCR循環數設置為30,來最大限度地防止所克隆片段的突變,從而使所克隆的基因序列具有很高的可信度.
使用生物信息學對所克隆的基因片段進行分析,BLAST比對結果和系統發育分析結果均表明所克隆片段為WRKY基因.亞細胞定位分析結果表明,GrWRKY2蛋白可能定位于細胞核,這與西洋參PqWRKY1的細胞核定位結果相一致[5].保守結構域分析結果表明,GrWRKY2蛋白包含DNA-binding WRKY(IPR003657)保守結構域,因此,筆者推測GrWRKY2蛋白可能通過該保守結構域來調控細胞核內基因的表達.
[1]金航,張霽,張金渝,等.滇龍膽[M].昆明:云南科技出版社,2013:1-5.
[2]Xu Y,Wang J,Wang S,etal.Characterization of GaWRKY1,a cotton transcription factor that regulates the sesquiterpene synthase gene (+)-δ-cadinene synthase-A[J].Plant Physiol,2004,135(1):507-515.
[3]Ma D,Pu G,Lei C,et al.Isolation and characterization of AaWRKY1, anArtemisiaannuatranscription factor that regulates the amorpha-4,11-diene synthase gene,a key gene of artemisinin biosynthesis[J].Plant Cell Physiol,2009,50(12):2146-2161.
[4]Spyropoulou E A,Haring M A,Schuurink R C.RNA sequencing onSolanumlycopersicumtrichomes identifies transcription factors that activate terpene synthase promoters[J].BMC Genomics,2014,15(1):402.
[5]Sun Y,Niu Y,Xu J,etal.Discovery of WRKY transcription factors through transcriptome analysis and characterization of a novel methyl jasmonate-induciblePqWRKY1 gene fromPanaxquinquefolius[J].Plant Cell, Tiss Org,2013,114(2):269-277.
[6]Schluttenhofer C,Pattanaik S,Patra B,etal.Analyses ofCatharanthusroseusandArabidopsisthalianaWRKY transcription factors reveal involvement in jasmonate signaling[J].BMC Genomics,2014,15(1):502.
[7]Zhang J,Zhang J,Wang Y,etal.Effects of tree species on seed germination and seedlings growth of Chinese medicinal herbGentianarigescens[J].Allelopathy J,2012,29(2):325-332.
[8]Zhang J,Wang Y,Yang T,etal.Use of gibberellic acid to overcome the allelopathic effect of a range of species on the germination of seeds ofGentianarigescens,a medicinal herb[J].Seed Sci Techno,2012,40(3):443-447.
[9]Wong K,But P P,Shaw P.Evaluation of seven DNA barcodes for differentiating closely related medicinal Gentiana species and their adulterants[J].Chinese Medicine,2013,8(1):16.
[10]Nakatsuka T,Saito M,Sato-Ushiku Y,etal.Development of DNA markers that discriminate between white- and blue-flowers in Japanese gentian plants[J].Euphytica,2012,184(3):335-344.
[11]Nakatsuka T,Yamada E,Saito M,etal.Heterologous expression of gentian MYB1R transcription factors suppresses anthocyanin pigmentation in tobacco flowers[J].Plant Cell Rep,2013,32(12):1925-1937.
[12]Nakatsuka T,Saito M,Yamada E,etal. Isolation and characterization of GtMYBP3 and GtMYBP4, orthologues of R2R3-MYB transcription factors that regulate early flavonoid biosynthesis,in gentian flowers[J].J Exp Bot,2012,63(18):6505-6517.
[13]Gu Y.The gentian returned after travelling by spacecraft Shenzhou-10 which suggested that space breeding project has been started in Yunnan Province[EB/OL].2013[cited 2013 2013-7-12];Available from:http://www.chinanews.com/gn/2013/07-12/5037538.shtml.
[14]張曉東,李彩霞,王連春,等.滇龍膽GrSLS1基因的克隆和原核表達[J].西北植物學報,2014,34(7):1311-1317.
[15]Rushton P J,Somssich I E,Ringler P,etal.WRKY transcription factors[J].Trends Plant Sci,2010,15(5):247-258.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
