類幾丁質酶3蛋白1在申克孢子絲菌刺激巨噬細胞的表達和作用研究
2015-03-17 01:12:57黃麗林高文超李玉哲袁立燕何泰龍黃懷球
中國人獸共患病學報 2015年10期
黃麗林,張 靜,高文超,李玉哲,袁立燕,何泰龍,黃懷球
類幾丁質酶3蛋白1在申克孢子絲菌刺激巨噬細胞的表達和作用研究
黃麗林1,張 靜2,高文超2,李玉哲1,袁立燕3,何泰龍1,黃懷球1
目的 探討巨噬細胞類幾丁質酶3蛋白1在申克孢子絲菌刺激后的表達和作用研究。方法 通過測定巨噬細胞與申克孢子絲菌共孵育時殺菌率,使用實時熒光定量 PCR檢測不同時間點巨噬細胞的類幾丁質酶3蛋白1 mRNA表達水平,同時體外實驗檢測類幾丁質酶3蛋白1對申克孢子絲菌的抑菌作用,同時采用空白對照和白念珠菌陽性對照比較兩者的差異。結果 巨噬細胞可吞噬殺滅申克孢子絲菌分生孢子,也可吞噬陽性對照白念珠菌酵母相細胞。巨噬細胞類幾丁質酶3蛋白1 mRNA表達水平在申克孢子絲菌組和空白對照組無明顯差別,作用于陽性對照的白念珠菌組的巨噬細胞類幾丁質酶3蛋白1 mRNA表達水平明顯上調。體外實驗顯示類幾丁質酶3蛋白1 對申克孢子絲菌分生孢子的無明顯抑制作用,對白念珠菌繁殖卻有較強的抑制作用。 結論 申克孢子絲菌未能誘導巨噬細胞表達類幾丁質酶3蛋白1 抑制其增殖從而逃逸清除,而白念珠菌感染可上調巨噬細胞類幾丁質酶3蛋白1 表達從而抑制其增殖減輕感染,這可能是申克孢子絲菌特有的致病作用機制之一。
申克孢子絲菌;白念珠菌;分生孢子;類幾丁質酶3蛋白1
幾丁質(Chitin) 是真菌細胞壁的重要成分, 對真菌生長和致病性具有重要作用[1]。引起孢子絲菌病的雙相型真菌孢子絲菌復合體中各相細胞包含大量幾丁質,在出芽繁殖過程中需要幾丁質酶、葡聚糖酶等降解芽生部位的幾丁質、葡聚糖等細胞壁成分。高等哺乳類動物不存在幾丁質,其消化道、呼吸道、泌尿道等可分泌幾丁質酶或幾丁質結合活性的幾丁質酶樣蛋白防御含幾丁質的病原體感染,起到機體固有免疫的防御作用[2-3]。類幾丁質酶3蛋白1(chitinase 3-like-1,CHI3LI)屬第18家族哺乳動物幾丁質酶家族成員,起催化活性作用的氨基酸發生突變可導致其喪失降解幾丁質的活性,研究表明機體感染多種病原體時均可誘導巨噬細胞分泌CHI3L1參與機體免疫功能 。巨噬細胞CHI3L1在防御申克孢子絲菌和白念珠菌感染的表達和作用未見報道。本研究通過檢測巨噬細胞時CHI3L1在申克孢子絲菌分生孢子、白念珠菌酵母相細胞刺激下的表達和作用,初步探討巨噬細胞CHI3L1在孢子絲菌病發病中的作用及其與白念珠菌感染的差異,將有助進一步探索申克孢子絲菌新的致病作用機制。
1 材料和方法
1.1 材料
1.1.1 菌株來源 申克孢子絲菌CBS359.36株由荷蘭皇家科學和技術院真菌多樣性研究中心提供,白念珠菌ATCC90028株,由中山大學附屬第三醫院醫學真菌研究中心提供。
1.1.2 實驗動物 雌性6~8周BALB/c小鼠由中山大學醫學院動物中心提供,體重20~25 g,實驗通過中山大學動物實驗倫理委員會批準。
1.1.3 主要試劑和儀器 沙氏培養基(SDA)、馬鈴薯葡萄糖瓊脂斜面培養基(PDA)(廣東環凱微生物科技有限公司),無菌磷酸緩沖液(1×PBS)(賽默飛世爾生物化學制品有限公司),低糖DMEM細胞培養基、RPMI-1640培養基、6孔和96孔圓底培養板和細胞培養瓶、胎牛血清、小鼠重組純化幾丁質酶3樣蛋白1(廣州德為生物公司),RNA提取試劑盒(上海碧云天公司),THUNDERBIRD SYBR?qPCR Mix Kit 和 first strand cDNA syhthesis kit (Toyobo公司),實驗中所用的引物由廣州中美泰和公司合成,CO2細胞培養箱(ThermoFisher scientific),超凈工作臺(蘇凈BHC-1300ⅡA/B3,蘇州安泰空氣技術有限公司)。超微量紫外、可見光分光光度計(ThermoFisher scientific), 實時熒光定量PCR系統(BIO-RAD IQ5)。
1.2 實驗方法
1.2.1 申克孢子絲菌分生孢子、白念珠菌酵母相細胞菌懸液制備 將申克孢子絲菌CBS359.36株(菌絲相)轉種至SDA斜面培養基,于培養箱中25 ℃培養活化7 d,然后轉種至馬鈴薯葡萄糖瓊脂(PDA)斜面培養基上,培養1周后有產孢。用槍頭將2 mL生理鹽水注入長滿菌落的斜面,輕輕吹打,然后吸出至EP管。室溫下靜置20 min左右,待菌絲沉淀后,輕輕將上清吸出至另一EP管。12 000 r/min 5 min,將上清去除,然后加入1 mL生理鹽水。用血球計數板調整菌懸液濃度至1×105cfu/mL。將已配置菌懸液稀釋后接種于PDA平板,驗證菌懸液活力及濃度。白念珠菌ATCC 90028轉種至SDA斜面培養基,室溫下培養72 h后,顯微鏡下觀察真菌細胞的形態是否以酵母相孢子為主。
1.2.2 小鼠巨噬細胞的制備 取20~25 g健康雌性6~8周,BALB/c小鼠36只,頸椎脫臼處死,消毒小鼠腹部皮膚,持鑷提起腹中部皮膚并剪開,暴露腹膜,避開血管分別腹腔注射無菌1×PBS 8 mL,反復輕揉腹壁5 min后,無菌注射器分別取腹腔液5~6 mL,5 000 r/min離心5 min,棄上清,加入DMEM培養液調整細胞濃度至1.0×106/mL,細胞懸液置入培養瓶,37 ℃,5% CO2細胞培養箱2 h后,吸棄上清,用預熱培養液洗2次,留下貼壁細胞,吉姆薩染色光學顯微鏡下顯示95%以上為巨噬細胞,加入DMEM培養加10%胎牛血清液孵育細胞。
1.2.3 申克孢子絲菌清除率的測定 將申克孢子絲菌分生孢子、白念珠菌酵母相細胞按每瓶1 mL(1.0×106/mL)接種于上述含1.0×106/mL,巨噬細胞的培養瓶中,每個時間點培養3份, 以等量的真菌孢子懸液和酵母相細胞作為空白對照,放入5%的CO2細胞培養箱,37 ℃孵育24 h、48 h、72 h、96 h后取出,在光學顯微鏡下觀察巨噬細胞外的申克孢子絲菌分生孢子、白念珠菌酵母相細胞出芽生殖和菌株形態改變并與空白對照組比較。然后轉至25 ℃下分別培養5 d、3 d,溫度的改變是為了更容易菌落計數。巨噬細胞活力通過臺盼藍染料的實驗證實。申克孢子絲菌分生孢子的殺傷率=[1-(CFU實驗組/CFU空白對照組)]×100%[4]。
1.2.4 Real-time PCR 檢測巨噬細胞CHI3L1 mRNA表達水平 實驗組為將申克孢子絲菌分生孢子、白念珠菌酵母相細胞按每瓶1 mL(1.0×106/mL)接種于上述含1.0×106/mL巨噬細胞的培養瓶中,空白對照組為1 mL生理鹽水加入含1.0×106/ mL巨噬細胞的培養瓶中,每個時間點培養3份,收集各個時間點共培養的巨噬細胞用RNA提取試劑盒法按照說明書提取細胞總RNA,核酸凝膠電泳提示28 s、18 s、5 s 3條清晰的條帶,采用紫外分光光度計評估其完整性并估算其產量。按FSQ-101反轉錄試劑盒說明書分別以0.5~1 μg 的總 RNA 為模板逆轉錄合成 cDNA。按照說明書采用FastStart DNA Master SYBR Green (Roche Diagnostics, Indianapolis, IN)嵌合熒光法,對目標基因進行相對定量分析,real-time PCR中用到的引物 CHI3L1 forward 5′- GTACAAGCTGGTCTGCTACTTC-3′ 和CHI3L1 reverse 5′- ATGTGCTAAGCATGTTGTCGC-3′,內參βactin forward 5′-CAGTAACAGTCCGCCTAGAA-3′ 和 βactin reverse 5′-GATTACTGCTCTGGCTCCTA-3′。
1.2.5 CHI3L1蛋白抑制孢子絲菌分生孢子芽生能力測定 配制RPMI-1640培養基稀釋CHI3L1蛋白并模擬體內37 ℃環境,驗證CHI3L1蛋白在體外對申克孢子絲菌分生孢子、白念珠菌酵母相細胞作用及其差異。96孔板第1至第6列孔濃度為0.5~16 μg/mL。將蛋白稀釋至2倍終濃度,按第1至第6列孔濃度由高至低的順序加樣,第7列及第8列孔分別作為陰性對照。取100 μL上述申克孢子絲菌分生孢子加入1至3列,而白念珠菌酵母相菌懸液加入4至6列孔混勻,并于37 ℃恒溫培養箱孵育120 h,取出后在倒置顯微鏡觀察真菌形態改變,然后轉種到固體PDA斜面培養基25 ℃下分別培養5 d、3 d。
1.2.6 統計學分析 采用SPSS 19.0統計軟件分析。量資料數據以均數±標準差表示。符合方差齊性采用組內采用t檢驗,組間采用方差分析,檢驗前檢查方差齊性,方差不齊者用秩和檢驗。P<0.05為差異有統計學意義。
2 結 果
2.1 巨噬細胞體外非調理性殺菌率 巨噬細胞在不同時間點對申克孢子絲菌、白念珠菌的殺菌率如圖1所示。巨噬細胞在24 h對申克孢子絲菌清除率為13.5%,48 h為16.7%,72 h為18.9%,96 h為22.1%,而對白念珠菌的清除率24 h為16.5%,48 h為21.7%,72 h為28.9%,96 h為31.1%。巨噬細胞對申克孢子絲菌分生孢子、白念珠菌酵母相細胞的非調理性殺菌率隨時間延長有所增多。結果顯示巨噬細胞對申克孢子絲菌及白念珠菌均可以吞噬及殺傷,在此過程中存在真菌細胞壁破壞和幾丁質的暴露,其中巨噬細胞對白念珠菌的殺菌能力高于申克孢子絲菌,各時間點組間比較菌存在統計學差異(P<0.05)。見圖1。
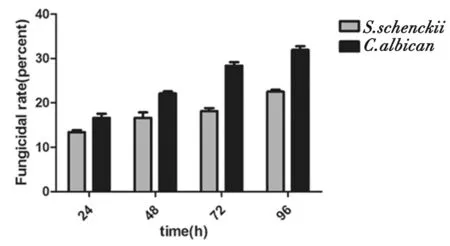
圖1 巨噬細胞的非調理性殺菌率
2.2 巨噬細胞外真菌形態觀察 采用光學顯微鏡下觀察巨噬細胞與孢子絲菌分生孢子、白念珠菌共培養過程中巨噬細胞外培養基中未吞噬的孢子絲菌分生孢子、白念珠菌的出芽及菌絲生長情況, A圖中黑色箭頭所示為巨噬細胞外申克孢子絲菌出芽,B圖中黑色箭頭為白念珠菌出芽及菌絲伸長,C圖和D圖為空白對照的申克孢子絲菌和白念珠菌。見圖2。

圖2 巨噬細胞外真菌的出芽和菌絲形成(×400)
Fig.2 Germination and hyphae of fungi outside macrophages(×400)
2.3 真菌對巨噬細胞的CHI3L1誘導表達 采用real-time PCR方法定量檢測CHI3L1 mRNA轉錄水平,結果顯示孢子絲菌共培養組和空白對照組相比,各時間點CHI3L1轉錄水平無明顯差異(P>0.05),而白念珠菌組CHI3L1的轉錄水平明顯上調,各時間點與申克孢子絲菌及空白對照組相比均有統計學差異(P<0.05)。見圖3。說明申克孢子絲菌不能有效誘導巨噬細胞表達CHI3L1,而白念珠菌可上調巨噬細胞CHI3L1的表達。
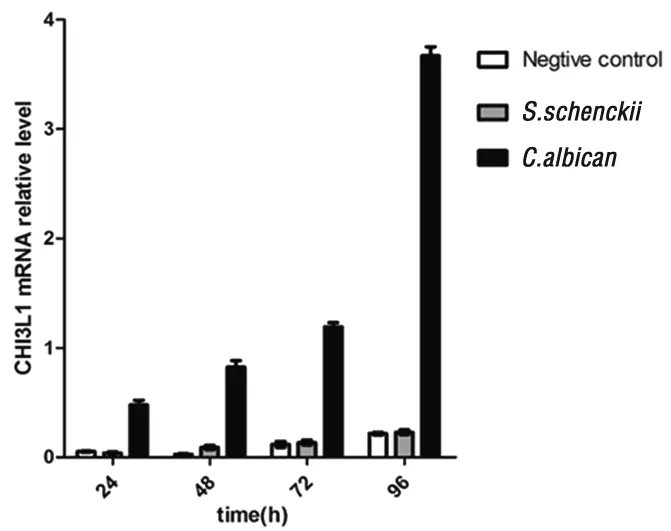
圖3 真菌對巨噬細胞的CHI3L1誘導表達差異
Fig.3 CHI3L1 differential expression in macrophages upon fungal stimulation
2.4 小鼠源性CHI3L1對真菌繁殖體外抑制實驗 光學顯微鏡下觀察顯示在 CHI3L1蛋白大于2 μg/ mL 濃度作用72 h作用后,在孢子絲菌和白念珠菌形態輕微異常,菌落計數結果顯示孢子絲菌的菌落形成數與空白對照組無明顯差異(P>0.05),說明CHI3L1蛋白對申克孢子絲菌分生孢子出芽生殖無明顯影響。而白念珠菌組菌落數量與對照組相比明顯減少(P<0.05),提示CHI3L1可抑制白念珠菌的繁殖。見圖4。
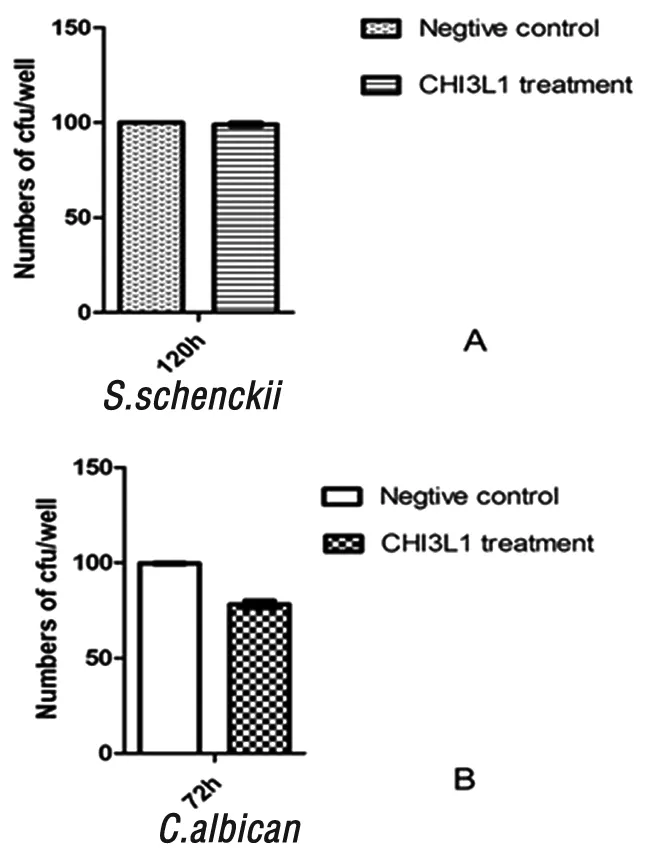
圖4 小鼠源性CHI3L1對真菌繁殖的抑制試驗
Fig.4 Fungal colony inhibition assay of mouse CHI3L1invitro
3 討 論
幾丁質是由其底物 N- 乙酰- D- 氨基葡萄糖通過 β- 1,4糖苷健聚合形成的直鏈大分子, 廣泛存在自然界中[5],作為申克孢子絲菌、白念珠菌等真菌細胞壁的重要成分,其與細胞的發生、維持和分隔形成有密切關系,對真菌生長、生理功能和致病性有重要意義[1]。高等哺乳類動物不含幾丁質,其消化道、呼吸道、泌尿道等可分泌幾丁質酶或有幾丁質結合活性的幾丁質酶樣蛋白防御含幾丁質的病原體感染[2-3]。其中CHI3LI屬第18家族哺乳動物幾丁質酶家族成員,起催化活性作用的氨基酸發生突變可導致其喪失降解幾丁質的活性,研究表明機體感染多種病原體時均可誘導巨噬細胞分泌CHI3L1參與調節機體免疫功能[6-7],含幾丁質的真菌等病原體感染時更為明顯。巨噬細胞是固有免疫的中重要的免疫細胞,可識別、吞噬、殺滅和消化提呈病原體,同時分泌一些蛋白酶、細胞因子參與機體免疫反應。申克孢子絲菌分生孢子、白念珠菌酵母相細胞等刺激巨噬細胞時,主要通過模式識別受體如TLR2、TLR4、DECTIN-1等識別、吞噬,然后利用胞內的多種殺菌機制殺滅胞內病原菌,如呼吸爆發產生ROS、蛋白酶、溶酶體等,發揮其固有免疫和抗原提呈功能[8-12]。探討巨噬細胞CHI3L1蛋白在孢子絲菌病發病中的作用及其與白念珠菌感染的差異,將有助進一步探索申克孢子絲菌新的致病作用機制。本研究通過檢測申克孢子絲菌分生孢子、白念珠菌酵母相細胞作用于巨噬細胞時CHI3L1的表達差異和觀察其對真菌的清除和抑制狀況,初步探討CHI3L1在孢子絲菌病發病中的作用及其與白念珠菌感染可能差異。
雖然申克孢子絲菌分生孢子可被巨噬細胞膜上免疫識別受體識別,進而被吞噬和殺滅。真菌細胞壁含有幾丁質,在其出芽生殖過程中需要幾丁質酶、葡聚糖酶等降解芽生部位等幾丁質和葡聚糖。但研究顯示培養上清中巨噬細胞外的孢子絲菌分生孢子出芽生殖和形態無明顯改變,體外實驗證實CHI3L1對孢子絲菌、白念珠菌形態改變無明顯作用,發現白念珠菌菌落計數明顯減少,但申克孢子絲菌數量卻無明顯異常。Real-time PCR 結果顯示申克孢子絲菌未能有效誘導CHI3L1的表達,白念珠菌卻能明顯上調巨噬細胞的CHI3L1表達,這可能與申克孢子絲菌細胞壁幾丁質含量較少和具有白念珠菌不同的結構特征有關。CHI3L1蛋白可通過結合幾丁質與真菌自身的幾丁質酶競爭結合位點,影響真菌細胞形態和阻礙真菌的出芽生殖,從而防御真菌感染增殖,而非直接催化降解幾丁質的功能。在肺炎鏈球菌、肝片吸蟲等病原體感染過程中,巨噬細胞主要通過誘導性分泌CHI3L1[6-7],促進分泌TGFβ1等Th2型細胞因子,從而促進纖維母細胞增殖,纖維化限制病原體擴散,同時促進M2型巨噬細胞轉化、抑制巨噬細胞凋亡等[6-7]。研究顯示CHI3L1可在炎癥性、過敏性疾病、腫瘤等非感染性疾病中誘導表達,CHI3L1已作為支氣管哮喘的診斷和疾病嚴重程度評估的分子標記和探索作為哮喘的免疫治療手段[13-14]。含有幾丁質的申克孢子絲菌未能有效誘導巨噬細胞CHI3L1的表達,孢子絲菌病常見病理表現為肉芽腫性炎癥改變,表現為炎癥細胞浸潤、纖維組織增生、血管新生等,其病理改變與Th2型細胞因子的作用有關,容易導致真菌感染遷延難愈[15]。如此認為,申克孢子絲菌未能誘導巨噬細胞表達類幾丁質酶3蛋白1 抑制其增殖從而逃逸清除,而白念珠感染可上調巨噬細胞類幾丁質酶3蛋白1 表達從而抑制其增殖減輕感染,這可能是申克孢子絲菌特有的致病作用機制之一。申克孢子絲菌分生孢子如何逃避巨噬細胞所誘導CHI3L1表達的殺傷作用機制需要通過進一步研究進行探討。
[1]Walker LA, Lenardon MD, Preechasuth K, et al. Cell wall stress induces alternative fungal cytokinesis and septation strategies[J]. J Cell Sci, 2013, 126(Pt 12):2668-2677. DOI:10.1242/jcs.118885
[2]Homer RJ, Zhu Z, Cohn L, et al. Differential expression of chitinases identify subsets of murine airway epithelial cells in allergic inflammation[J]. Am J Physiol Lung Cellul Mol Physiol, 2006, 291(3):L502-511. DOI:10.1152/ajplung.00364.2005
[3]Mizoguchi E. Chitinase 3-like-1 exacerbates intestinal inflammation by enhancing bacterial adhesion and invasion in colonic epithelial cells[J]. Gastroenterology, 2006, 130(2):398-411. DOI:10.1053/j.gastro.2005.12.007
[4]Dong BL, Li DS, Liu WH, et al. Differential expression of reactive oxygen species in human neutrophils stimulated by the yeast phase of Sporothrix Schenckii and Candida albicans[J]. Chin J Dermatol, 2013, 46(4):253-257. (in Chinese) DOI:10.3760/cma.j.issn.0412-4030.2013.04.009 董碧麟, 李東升, 柳衛凰, 等.申克孢子絲菌酵母相與白念珠菌誘導中性粒細胞活性氧簇的差異性表達[J].中華皮膚科雜志,2013,46(4):253-7. DOI:10.3760/cma.j.issn.0412-4030.2013.04.009
[5]Wagener J, Malireddi RK, Lenardon MD, et al. Fungal chitin dampens inflammation through IL-10 induction mediated by NOD2 and TLR9 activation[J]. PLoS Pathog, 2014, 10(4):e1004050. DOI:10.1371/journal.ppat.1004050
[6]Dela Cruz CS, Liu W, He CH, et al. Chitinase 3-like-1 promotesStreptococcuspneumoniaekilling and augments host tolerance to lung antibacterial responses[J]. Cell Host Microbe, 2012, 12(1):34-46. DOI:10.1016/j.chom.2012.05.017
[7]Flynn RJ, Irwin JA, Olivier M, et al. Alternative activation of ruminant macrophages byFasciolahepatica[J]. Vet Immunol Immunopathol, 2007, 120(1-2):31-40. DOI:10.1016/j.vetimm.2007.07.003
[8]Martinez-Alvarez JA, Perez-Garcia LA, Flores-Carreon A, et al. The immune response againstCandidaspp. andSporothrixschenckii[J]. Revista Iberoamericana de Micologia, 2014, 31(1):62-66. DOI:10.1016/j.riam.2013.09.015
[9]de CNT, Ferreira LS, Arthur RA, et al. Influence of TLR-2 in the immune response in the infection induced by fungusSporothrixschenckii[J]. Immunolog Investigat, 2014, 43(4):370-390. DOI:10.3109/08820139.2013.879174
[10]Sassa MF, Saturi AE, Souza LF, et al. Response of macrophage Toll-like receptor 4 to aSporothrixschenckiilipid extract during experimental sporotrichosis[J]. Immunology, 2009, 128(2):301-309. DOI:10.1111/j.1365-2567.2009.03118.x
[11]Kajiwara H, Saito M, Ohga S, et al. Impaired host defense againstSporothrixschenckiiin mice with chronic granulomatous disease[J]. Infect Immun, 2004, 72(9):5073-5079. DOI:10.1128/IAI.72.9.5073-5079.2004
[12]Gow NA, Netea MG, Munro CA, et al. Immune recognition ofCandidaalbicansbeta-glucan by dectin-1[J]. J Infect Dis, 2007, 196(10):1565-1571. DOI:10.1086/523110
[13]Duru S, Yuce G, Ulasli SS, et al. The relationship between serum YKL-40 levels and severity of asthma[J]. Iranian J Allergy Asthma Immunol, 2013, 12(3):247-253.
[14]Muzzarelli RA. Chitins and chitosans as immunoadjuvants and non-allergenic drug carriers[J]. Marine Drugs, 2010, 8(2):292-312. DOI:10.3390/md8020292
[15]Carlos IZ, Sassa MF, da Graca Sgarbi DB, et al. Current research on the immune response to experimental sporotrichosis[J]. Mycopathologia, 2009, 168(1):1-10. DOI:10.1007/s11046-009-9190-z
Differential expression and function of chitinase 3-like-1 in macrophage stimulated bySporothrixschenckii
HUANG Li-lin1,ZHANG Jin2,GAO Wen-chao2,LI Yu-zhe1,YUAN Li-yan3,HE Tai-long1,HUANG Huai-qiu1
(1.DepartmentofDermatologyandVenereology,theThirdAffiliatedHospital,SunYat-senUniversity,Guangzhou510630,China;2.DepartmentofDermatologyandVenereology,SunYat-senMemorialHospital,SunYat-senUniversity,Guangzhou510120,China;3.DepartmentofDermatology,GuangdongProvincialCenterforSkinDiseasesandSTIsControlandPrevention,Guangzhou510500,China)
We evaluated the differential expression and function of chitinase 3-like-1 in macrophage stimulated bySporothrixschenckiiandCandidaalbicansfungicidal ability of macrophage after stimulation withSporothrixschenckiiandCandidaalbicansseparately was detected. The expression of CHI3L1 gene in macrophage stimulated bySporothrixSchenckiiandCandidaalbicanswas evaluated with real-time PCR. The function of CHI3L1 protein in macrophages against the reproduction ofSporothrixschenckiiandCandidaalbicanswas detectedinvitro. Results showed that macrophages could engulf and killSporothrixSchenckiiandCandidaalbicansinvitro. The expression of CHI3L1 gene in macrophage stimulated byCandidaalbicanswas increased obviously. At the same time, CHI3L1 protein can damper the reproduction ofCandidaalbicans. However, the expression of CHI3L1 gene was not elevated when macrophage was stimulated bySporothrixschenckiiand CHI3L1 protein played little role in reproduction ofSporothrixschenckii. The expression of CHI3L1 gene in macrophage was elevated after stimulation withCandidaalbicans, but was not elevated withSporothrixSchenckii. In correspondence with differential expression, CHI3L1 in macrophages could impair the reproduction ofCandidaalbicansbut had a weak function onSporothrixschenckiiwhich might contribute to the pathogenesis of sporotricosis. Keywords:Sporothrixschenckii;Candidaalbicans; conidium; chitinase 3-like-1
Huang Huai-qiu, Email:hhuaiqiu@medmail.com.cn
國家自然科學基金面上項目(No.81371746),廣東省省級科技計劃項目(No.2013B010404013)
黃懷球,Email:hhuaiqiu@medmail.com.cn
1.中山大學附屬第三醫院皮膚病科,廣州 510630; 2.中山大學孫逸仙紀念醫院皮膚病科,廣州 510120; 3.廣東省皮膚性病防治中心,廣州 510500
10.3969/j.issn.1002-2694.2015.10.009
R379.2
A
1002-2694(2015)10-0938-05
2015-03-03;
2015-08-03
Supported by the National Nature Foundation of China (No. 81371746) and the Guangdong Provincial Science and Technology Foundation Committee (No. 2013B010404013)
