生殖支原體MG 427原核表達載體的構建和鑒定
2015-03-12 08:38:26周俊陳列松游曉星曾焱華吳移謀
當代醫學 2015年22期
周俊 陳列松 游曉星 曾焱華 吳移謀
生殖支原體(mycoplasma genitalium,Mg),是迄今發現能進行自我復制的最小的原核微生物,透射免疫電鏡已證實Mg在宮頸上皮細胞內完整存在,在細胞培養模型中,M g能快速地粘附到上皮細胞,定植于細胞內,產生強烈的細胞免疫。M g侵入人類靶細胞后,導致宿主細胞持續性慢性致病,且癥狀隱蔽或無癥狀,可能其具有一系列的自身抵抗氧化應激的機制來確保其持續性感染和致病。先前有報道氧化修復酶蛋氨酸亞砜還原劑A(M srA)在氧化應激中能保護生殖支原體(M g)[1],近年來發現,過氧化物酶基因家族中的有機氫過氧化物抗性蛋白(Oh r),烷基氫過氧化物還原酶(AHPR)及滲透壓誘導蛋白C(Osm C)分布于各種病原體,在感染部位將有機氧化物變為醇類的解毒過程中發揮了重要作用。同樣地,生殖支原體基因組中也發現了相應的Oh r/Osm C同系物[2-3],作為滲透壓調節器系統被廣泛研究的Osm C直到最近發現與Oh r相似性后才揭示其功能,有學者認為部分Osm C蛋白與Oh r一樣,能在體外降解H2O2和有機氫過氧化物[4-5]。生殖支原體MG 427是否有其類似功能,國內外研究甚少,本研究利用重組DNA技術構建了原核表達載體pGEX-6 p-1/MG 427,從而為進一步研究Osm C樣蛋白,闡明Mg的致病機制奠定基礎。
1 材料與方法
1.1 主要試劑 Mg-G 37標準株、pGEX-6 p-1載體及E.coliJM 109均為南華大學病原微生物研究所保存。細菌DNA提取及純化試劑盒、質粒提取試劑盒、基因定點突變試劑盒為美國Axygen產品;Taq DNA聚合酶、限制性內切酶BamHI與NotI、T 4連接酶購自美國NEB公司;
1.2 細菌基因組DNA提取及MG 427擴增 采用細菌基因組DNA提取試劑盒從培養好的Mg標準株G 37中提取基因組DNA。從Gen Bank中獲取mg427全基因序列,依據pGEX 6p-1載體上的多克隆酶切位點設計mg427基因的特異性引物。P 1:CGGAT AAA AAA TAC GAT ATC ACA,P 2: AAGGAAAAAATTA GTG GAC TAA AGT AAC GCT A。按94℃預變性5m in; 94℃變性30 s;55℃退火45 s;72℃延伸60 s;共35個循環的參數進行PCR擴增;末次循環后72℃延伸5m in,終止反應。瓊脂糖凝膠中電泳并在紫外燈下觀察,將產物純化回收保存。
1.3 PCR產物和pGEX-6 p-1空載體的酶切 雙酶切總體積為40.0μL,均含: 10×NEB bu ffer 3 4.0μL;100×BSA 0.4μL;BamHI 1.0μL;NotI 1.0μL;其中PCR雙酶切體系中加mg427純化產物10.0μL;d d H2O 23.6μL; pGEX-6 p-1載體雙酶切體系中加PGEX-6P-1 15μL;ddH2O 18.6μL;然后分別置入37℃水浴箱中,保溫酶切12 h。取酶切后于1.7%的瓊脂糖凝膠上電泳,紫外燈下觀察目的條帶并回收純化目的片段。
1.4 原核表達重組體pGEX-6 p-1/MG 427的構建 將純化好的pGEX-6 p-1雙酶切產物 2.5μL與mg 427PCR純化產物6.0μL,加入T 4 DNA Linkage 0.5μL、T 4 buffer 1.0μL;置于22℃水循環儀中,連接30m in。之后轉化至宿主E.coliJM 109構建pGEX-6 p-1/MG 427,經培養篩選陽性克隆對重組體進行擴增、雙酶切及測序鑒定。
1.5 PCR介導的定點突變 在mg 427基因序列中第289~291位核苷酸所組成的密碼子為TGA,為確保其在E.coli中表達,將其同義突變為TGG,突變引物如下:TB 1:5′—CGT GAA CTT AAC ATT CAC TGG GAA ATT CAC TCT CCT AAT G—3′,TB 2:5′—C ATT AGG AGA GTG AAT TTCGTG AAT GTT AAG TTC ACG—3 ,突變擴增條件如下:95℃ 30 s;95℃ 50 s,55℃ 40 s,68℃ 5m in,共12個循環,68℃補全延伸10m in,4℃保存。在反應體系中加1μLDpnI限制性內切酶37℃下恒溫水浴箱孵育1 h消化目板質粒DNA。將突變好的產物轉化感受態E.coliXL-Blue。篩選陽性克隆對重組體進行雙酶切及測序鑒定。
2 結果
2.1mg 427基因的擴增 用特異性引物擴增MG-37標準株目的基因mg427,獲得約426 bp大小條帶,與目的片段大小一致。見圖1。
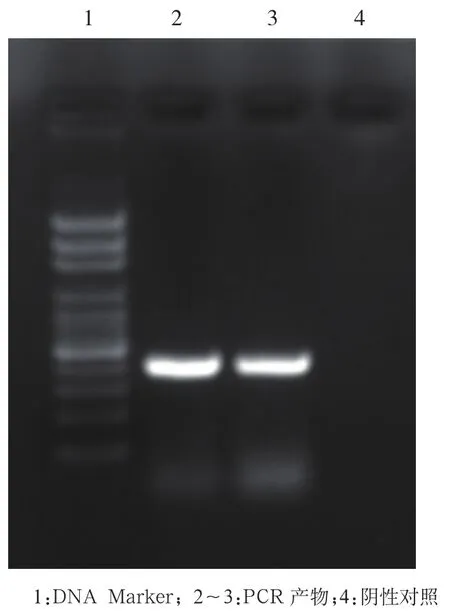
圖1 瓊脂糖電泳分析mg427 PCR擴增結果
2.2 pGEX-6 p-1/MG 427重組質粒的鑒定 PCR擴增重組好的pGEX-6 p-1/MG 427質粒也可獲得約426 bp的片段;經BamHI和NotI雙酶切重組質粒,結果經瓊脂糖凝膠電泳,在紫外線下可見:大片段接近5000 bp,小片段與目的基因長度426 bp相近,初步證明mg427目的基因成功地嵌入pGEX-6 p-1 載體中。見圖2。
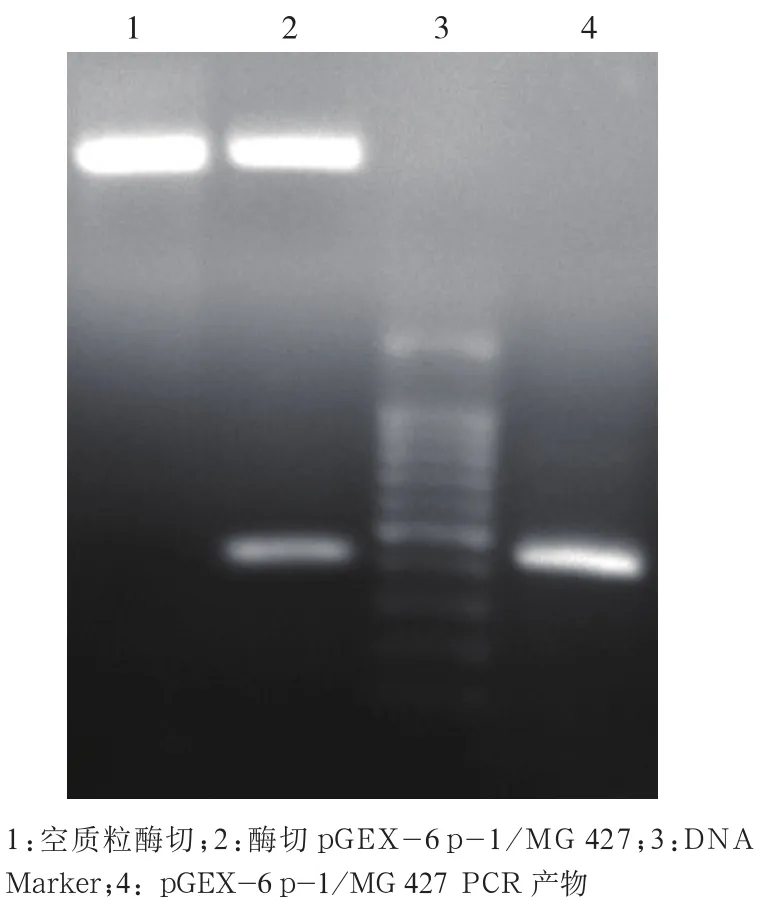
圖2 重組質粒pGEX-6 p-1/MG 427的酶切與PCR鑒定
2.3 重組質粒目的基因測序 將重組質粒送上海生工進行DNA序列測定,測序結果與GenBank所登錄的Mg G 37的mg 427序列進行blast,結果顯示二者完全一致,且密碼子讀碼框架正確。
2.4 PCR介導的定點突變后目的基因測序 將突變后的重組質粒進行測序,測序結果與GenBank所登錄的Mg G 37的mg 427序列進行blast,在mg 427基因序列中第289~291位核苷酸所組成的密碼子為TGA突變為TGG,整個核苷酸序列未發生缺失、錯配、移碼。
3 討論
支原體的致病機制主要包括掠奪宿主細胞營養成分、黏附或融合以及免疫病理引起的損傷。當支原體感染宿主細胞后,能在原發部位產生ROS,包括超氧化物、H2O2以及有機氫過氧化物等(OHPS)[6-7],這些物質作用于DNA堿基或磷酸核糖部分,可引起病原體DNA不可逆的損傷[8-9],這要求支原體必須具備某些保護性機制以防止宿主對自身損傷,以維持其入侵宿主細胞,并能在在宿主內形成慢性感染的過程中具有非常重要的意義。通過我們前期的生物信息學研究,發現在Mg種系群中也有Osm C的基因同源編碼序列mg 427存在,然而其功能目前尚未明了。
本研究以Mg G 37基因組DNA為模板,PCR擴增出mg 427基因序列,同時選用pGEX-6 P-1為表達載體,構建pGEX-6 p-1/mg 427重組質粒。由于Mg的一個重要的分子生物學特征是在其轉錄翻譯過程中以TGA作為色氨酸的密碼子,而在E.coli中TGA卻是轉錄終止的信號,這影響了Mg基因在E.coli中的表達。我們利用單點突變技術將TGA特異性突變為TGG。突變篩選后再行測序鑒定,確使該基因全序列在大腸埃希菌中得以高效表達。通過對重組質粒進行酶切鑒定和進一步的核苷酸序列分析,證實了插入序列與目的基因的一致性及讀碼框正確。在成功構建原核表達載體pGEX-6 p-1/mg 427后,為后續進一步研究生殖支原體的致癥機制提供了可能。
[1] Dhandayuthapani S,Blaylock MW,Bebear CM,et al.Peptide methionine sulfoxide reductase (MsrA) is a virulence determinant in Mycoplasma genitalium[J].JBacteriol,2001,183:5645-5650.
[2] Zhang W,Baseman JB.Functional characterization of osmotically inducible protein C (MG_427) from Mycoplasma genitalium[J].J Bacteri ol,2014,196(5):1012-1019.
[3] Caswell CC,Baumgartner JE,Martin DW,et al.Characterization of the organic hydroperoxide resistance system of Brucella abortus 2308[J].J Ba cteriol,2012,194(18):5065-5072.
[4] Saikolappan S,Das K,Sasindran SJ,et al.OsmC proteins of Mycobacterium tuberculosis and Mycobacterium smegmatis protect against organic hydroperoxide stress[J].Tuberculosis (Edinb),2011,91(1):S119-S127.
[5] Ta P,Buchmeier N,Newton GL,et al.Organic hydroperoxide resistance protein and ergothioneine compensate for loss of mycothiol in Mycobacterium smegmatis mutants[J].J Bacteri ol,2011,193(8):1981-1990.
[6] Machado CX 1,Pinto PM,Zaha A,et al.A peroxiredoxin from Mycoplasma hyopneumoniae with a possible role in H2O2 detoxification[J].Microbiolog y,2009,155(Pt 10):3411-3419.
[7] Sornchuer P,Namchaiw P,Kerdwong J,et al.Copper chloride induces antioxidant gene expression but reduces ability to mediate H 2O2 toxicity in Xanthomonas campestris[J].Microbiology,2014,160(Pt 2):458-466.
[8] Chen Z,Lu M,Zhuang G,et al.Enhanced bacterial biosensor for fast and sensitive detection of oxidatively DNA damaging agents[J].Anal Chem,2011,83(9):3248-3251.
[9] Fontenelle C,Blanco C,Arrieta M,et al.Resistance to organic hydroperoxides requiresohr and ohrR genes in Sinorhizobium meliloti[J].BMC Microbiol,2011(11):100-107.
