無瓣海桑與鄉土紅樹植物混交對林地大型底棲動物的影響
2015-02-05 06:14:42唐以杰方展強吳映明李燕妮譚莉君楊學彬陳思敏
生態學報 2015年22期
唐以杰,方展強,何 清,吳映明,陸 棣,李燕妮,譚莉君,楊學彬,陳思敏
1 廣東第二師范學院生物系, 廣州 510303 2 華南師范大學生命科學學院,廣東省高等學校生態與環境科學重點實驗室, 廣州 510631
無瓣海桑與鄉土紅樹植物混交對林地大型底棲動物的影響
唐以杰1,方展強2,*,何 清1,吳映明1,陸 棣1,李燕妮1,譚莉君1,楊學彬1,陳思敏1
1 廣東第二師范學院生物系, 廣州 510303 2 華南師范大學生命科學學院,廣東省高等學校生態與環境科學重點實驗室, 廣州 510631
選擇廣東省雷州市附城鎮和珠海市淇澳島沿海1年生無瓣海桑人工林,分別在林下混種鄉土紅樹植物紅海欖或木欖幼苗,對混交林和無瓣海桑純林林地大型底棲動物群落進行比較,探討無瓣海桑與鄉土紅樹植物混交對林地大型底棲動物的影響。結果顯示,混交林和無瓣海桑純林之間大型底棲動物群落的優勢種存在差異;相似性分析檢驗(One-Way ANOSIM)、等級聚類和非參數多變量標序結果均表明1年無瓣海桑+紅海欖混交林、1年無瓣海桑+木欖混交林和1年無瓣海桑純林之間大型底棲動物群落結構差異顯著。研究還發現在無瓣海桑人工林林下混交紅海欖或木欖這兩種鄉土紅樹植物,可提高林地底棲動物的生物量和物種多樣性。BIOENV分析說明大型底棲動物分布與紅海欖或木欖這兩種混交的鄉土樹種的凋落物量密切相關,這進一步證實了混交鄉土紅樹植物對林地底棲動物多樣性和分布的影響。兩個研究地實驗結果均顯示,在無瓣海桑林下種植木欖的效果要優于紅海欖,表現在木欖的平均苗高、凋落物量、凋落物量占群落凋落物總量百分比和幼苗成活率均高于紅海欖,其對提高林地大型底棲動物生物量和物種多樣性的效果也明顯優于紅海欖。
無瓣海桑;木欖;紅海欖;大型底棲動物;物種多樣性
無瓣海桑(Sonneratiaapetala)從1985年自孟加拉引種我國海南島,90年代從海南引種到廣東、廣西、福建等地。由于該樹種適應性強、生長迅速,近年已成為我國華南沿海灘涂紅樹林造林的重要樹種[1],無瓣海桑的引種擴大了我國紅樹林的種質資源,促進了紅樹林的恢復與發展,而且在控制灘涂互花米草的滋生蔓延方面取得了良好效果。然而,目前無瓣海桑引種后多形成了單種純林,可能是導致鄉土紅樹植物難以在其間定居生長的重要原因,同時純林的生物多樣性低、生態系統穩定性差、易發生病蟲害[2]。調查發現,無瓣海桑人工林林地大型底棲動物生物多樣性明顯低于同林齡鄉土紅樹植物人工林[3- 4],其中一個重要原因是無瓣海桑凋落物的營養水平(如C/N 比)和單寧含量明顯不同于鄉土紅樹植物,本地一些底棲動物對這種食物源“不適應”或“不喜歡”[5]。因此,有學者建議逐步對現有無瓣海桑純林進行改造,修枝后在林下種植適宜的鄉土樹種以形成混交結構[1]。然而,利用鄉土紅樹植物與無瓣海桑混交,對林地底棲動物群落會有怎樣的生態影響?未見報道。實際上,在紅樹林生態系統中,大型底棲動物作為紅樹林生態系統中物質循環、能量流動積極的消費者和轉移者,對紅樹林生態系統的生態功能有重要意義,研究大型底棲動物的多樣性和豐度對人工恢復紅樹林的生境變化均具有潛在的生物、生態指示作用[6]。鑒于此,本研究選擇湛江雷州市附城鎮和珠海市淇澳島兩地同為1年林齡的無瓣海桑純林各一片,分別在林下種植木欖或紅海欖幼苗,比較混交后的無瓣海桑+木欖、無瓣海桑+紅海欖混交林與無瓣海桑純林林地間大型底棲動物群落的差異,探討無瓣海桑與鄉土紅樹植物混交后大型底棲動物群落的生態響應,以及影響大型底棲動物群落結構和物種多樣性的主要環境因子。
1 材料與方法
1.1 研究區域概況
研究地位于廣東省湛江市雷州市附城鎮和珠海市淇澳島兩地的人工紅樹林區,其中雷州市人工紅樹林區東經為110°09′—110°10′,北緯20°52′—20°56′,屬北熱帶和南亞熱帶季風氣候區,年均氣溫22.8—23.4 ℃,年均水溫25—27 ℃,年均太陽總輻射4 500—5 600 MJ/m2,年均降水量1 534.6 mm,潮汐屬不正規半日潮,土壤多為濱海沙土和濱海鹽土[7]。另外一個研究地珠海市淇澳島紅樹林自然保護區東經為113°36′40′′—113°39′15′′,北緯為22°23′40′′—22°27′38′′,屬于低緯度亞熱帶季風區,日照充足,年平均氣溫22.4 ℃,最冷月1月的平均氣溫15.3 ℃,最熱月7月的平均氣溫28.3 ℃;年平均降雨量1 700—2 200 mm,東風為常向風,空氣年平均相對濕度為79%。潮汐屬不正規半日潮,平均高潮位0.17 m,平均低潮位-0.14 m,水質比較清潔;土壤屬于濱海鹽漬草甸沼澤土[8]。
1.2 采樣方法及樣品處理
分別在雷州市附城鎮沿海紅樹林區和珠海市淇澳島紅樹林自然保護區,選擇1年林齡的無瓣海桑人工純林;其中附城鎮純林面積約為3500 m2,自陸緣向海的林帶寬度約為50 m,無瓣海桑林的種植密度為2 m × 2 m;珠海市淇澳島純林面積約為4900 m2,自陸緣向海的林帶寬度約為70 m,無瓣海桑林的種植密度也為2 m × 2 m。實驗時,將純林分為大致相等的3部分,中間相隔5 m,左邊部分不進行混交,以作對照;中間部分在林下種植紅海欖幼苗,右邊部分在林下種植木欖幼苗,種植前,對于郁閉度大于0.6的無瓣海桑林分,進行砍枝處理;對于郁閉度不足0.6林分,對林下的互花米草或雜草、雜灌清除干凈;接著選擇無病蟲害的健壯紅海欖或木欖袋苗進行種植,種植規格為行距×株距=0.6 m×0.6 m。最后對種植后的紅海欖或木欖苗木進行撫育管理,即清除種植幼苗周圍互花米草以及雜草、雜灌和垃圾。混交3個月后,分別于2013年5、8、11月和2014年2月進行相關調查。大型底棲動物的調查是自陸緣向海,將每種人工林按林帶寬度分為等距離的4處,每處布設1個站點。其中,雷州市附城鎮1年無瓣海桑純林、1年無瓣海桑+紅海欖混交林和1年無瓣海桑+木欖混交林的4個站點分別標記為A1—A4、B1—B4和C1—C4;珠海市淇澳島1年無瓣海桑純林、1年無瓣海桑+紅海欖混交林和1年無瓣海桑+木欖混交林的4個站點分別標記為D1—D4、 E1—E4和F1—F4。每個站點均與陸緣平行方向上取4個50 cm × 50 cm的定量樣框,先撿出框內底表動物,再挖至30 cm深的土壤,用0.5 mm孔徑篩子分選出底內動物,所獲樣品用5%的甲醛溶液現場固定,于實驗室內分析鑒定。在進行大型底棲動物調查的同時,每個站點布設一個10 m × 5 m的樣方,對樣方內所有植物進行全面調查,測定各樣方內無瓣海桑的平均樹高和郁閉度,紅海欖或木欖苗高和成活率;每個站點在動物取樣樣方的右側,與動物取樣樣方相隔5 m,用孔徑1.5 mm玻璃纖維網布設置3個圍網,每個圍網所圈的地面面積為1 m2,每5天收集圍網內的凋落物,仔細辨別凋落物,按無瓣海桑、紅海欖或木欖分開,每月月末將收集的凋落物集中測定各種紅樹植物的凋落物量。同時,在每個測量無瓣海桑平均樹高等指標的10 m × 5 m的樣方中,按對角線隨機選5個樣點,挖取0—30 cm層的底泥,均勻混合,取1 kg左右的混合土壤作為測試樣品。土壤粒徑組成、鹽分和有機質含量分別用比重計法、重量法和重鉻酸鉀法測定,土壤pH值用電位法,每個樣品3次重復,各樣林生境理化因子水平取全年的平均值。
1.3 大型底棲動物數據分析
1.3.1 物種數和物種優勢度指數(Y)[9]
Y=Pi×fi
(1)
式中,Pi為研究區種i的個體數占總個體數的比例,fi為種i在研究區內各站位出現的頻率,當Y>0.02時,該種為優勢種。物種數和物種優勢度指數計算的數據取各人工林4個站點的全年平均值。
1.3.2 棲息密度和生物量差異顯著性分析
以季節和生境為因素,用Excel 2010對不同林分濕地中大型底棲動物群落的平均棲息密度和平均生物量進行無重復雙因素方差分析。
1.3.3 多樣性測度
采用Margalef物種豐富度指數(d)、Shannon-Wiener多樣性指數(H′) 以及Pielou均勻度指數(J′)。
Margalef物種豐富度指數
d=(S-1)/log2N
(2)
Shannon-Wiener指數
(3)
Pielou均勻度指數
(4)
式中,S為總種類數,N為總個體數,Pi為第i種的個體數與樣品中的總個數的比值。
1.3.4 動物群落結構的分析
大型底棲動物群落結構的多變量分析采用非參數多變量群落結構分析方法,對各站點的豐度數據先進行全年平均處理,為了減少機會種對群落結構的干擾,刪除在總體中相對豐度 < 1%的種類,但保留其中任一站點相對豐度>3%的種類。根據各站點中累計占89%的大型底棲動物種類豐度(經平方根轉化)為基礎構建Bray-Curtis相似性矩陣,再進一步進行類平均法站點等級CLUSTER聚類和非參數多變量排序(MDS),同樣,利用上述的Bray-Curtis相似性矩陣,用One-Way ANOSIM分析來檢驗不同生境之間群落結構的差異;用SIMPER分析鑒定對各生境動物群落劃分起主要作用的種;利用BIOENV分析各生境中大型動物群落與對應環境因子的相關系數,分析和選擇能夠解釋動物群落結構的最佳環境因子組合。用RELATE檢驗揭示大型底棲動物群落和環境(非)相似性矩陣之間相關性,然后利用PRIMER軟件包中的圖形分析和多變量分析等方法對群落結構進行研究[10]。
2 結果與分析
2.1 各站點環境理化因子水平
從表1可以看出,兩個研究地均顯示在無瓣海桑純林下種植木欖幼苗,其平均苗高、凋落物量、凋落物量占群落凋落物總量百分比和成活率都高于紅海欖,單因素方差分析顯示下木層的這4個因子水平差異極顯著(P< 0.01),這說明在無瓣海桑純林下種植木欖的效果要優于紅海欖。雷州市3種人工林上木層無瓣海桑的樹高和凋落物量差異不顯著(P> 0.05),郁閉度差異顯著(P< 0.05)。珠海市3種人工林上木層無瓣海桑的樹高、凋落物量和郁閉度差異均不顯著(P> 0.05)。從表1還可以看出不管是雷州還是珠海,同一研究地3種類型人工林的土壤理化因子水平差異都不顯著(P> 0.05)。

表1 兩個研究地3種人工林的理化因子水平 (平均值±標準差)Table 1 Level of physical-chemical factors of the three kinds of plantations at the two research sites (Average ± S.D.)
2.2 兩個研究地3種人工林大型底棲動物種類組成和優勢種
調查發現,雷州市3種紅樹人工林的底棲動物以甲殼類居多,其次為腹足類;而珠海市3種紅樹人工林以腹足類居多,其次為甲殼類。無論雷州市還是珠海市,1年無瓣海桑+木欖混交林的大型底棲動物物種數明顯高于1年無瓣海桑純林和1年無瓣海桑+紅海欖混交林。雷州市1年無瓣海桑+紅海欖混交林物種數明顯高于1年無瓣海桑純林,而珠海市1年無瓣海桑+紅海欖混交林物種數略低于1年無瓣海桑純林(表2)。
從表3可見,同一地點不同類型人工林大型底棲動物的優勢種存在差異,不同地點同一類型人工林大型底棲動物的優勢種也存在差異。無論雷州市還是珠海市,兩個地點均是1年無瓣海桑+木欖混交林優勢種種數高于1年無瓣海桑純林和1年無瓣海桑+紅海欖混交林。
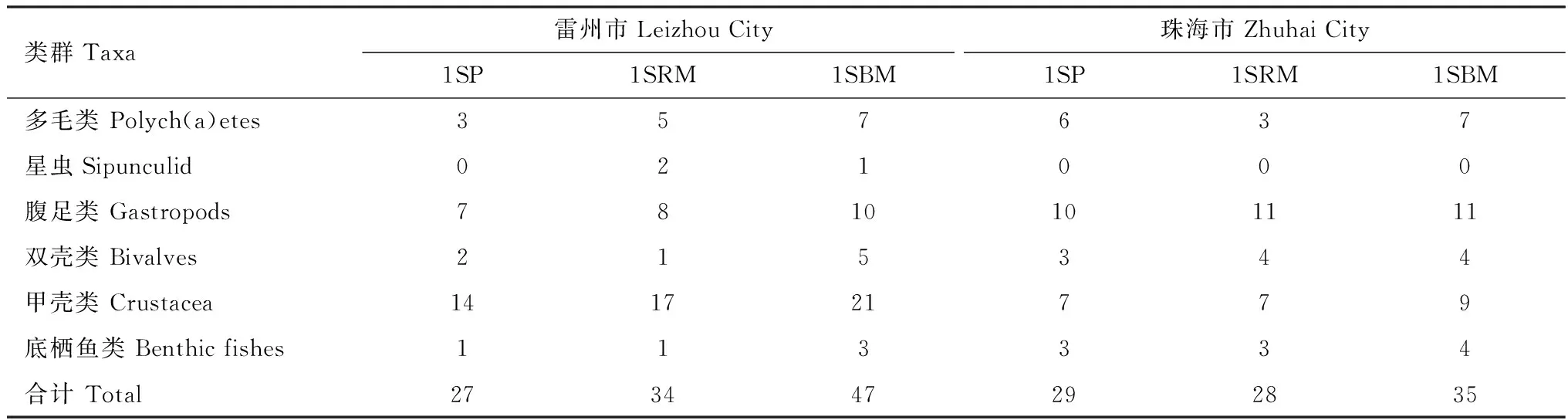
表2 兩個研究地3種人工林大型底棲動物物種分布Table 2 Species distribution of macrofauna of the three kinds of plantations at the two research sites

表3 兩個研究地3種人工林大型底棲動物優勢種Table 3 Dominant species of macrofauna of the three kinds of plantations at the two research sites
2.3 兩個研究地3種人工林大型底棲動物密度和生物量
兩個研究地不同類型人工林大型底棲動物群落密度和生物量的季節變化見表4,以季節和人工林類型為因素的密度和生物量無重復雙因素方差分析顯示雷州市附城紅樹林區大型底棲動物密度在3種人工林間差異極顯著(F=766.10,P< 0.01),季節間差異也顯著(F=89.84,P< 0.01);生物量在3種人工林間差異顯著(F=456.50,P< 0.01),季節間差異也顯著(F=56.35,P< 0.01)。珠海市淇澳島紅樹林區大型底棲動物密度在3種人工林間差異不顯著(F=4.38,P> 0.05),季節間差異顯著(F=9.66,P< 0.01);生物量在3種人工林間差異顯著(F=9.41,P< 0.01),季節間差異也顯著(F=8.12,P< 0.05)。兩個研究地均顯示混交林林地大型底棲動物生物量要比同林齡的無瓣海桑純林高,尤其是1年無瓣海桑+木欖混交林底棲動物生物量最高。
2.4 兩個研究地3種人工林大型底棲動物物種多樣性
兩個研究地3種類型人工林大型底棲動物群落物種多樣性指數的變化見表5,以季節和人工林類型為因素的多樣性指數無重復雙因素方差分析顯示大型底棲動物Shannon-Wiener多樣性指數在雷州市附城紅樹林區3種人工林間差異極顯著(F=1179.02,P< 0.01),季節間差異也顯著(F=7.96,P< 0.05);Pielou均勻度指數不同人工林間差異極顯著(F=311.47,P< 0.01),季節間差異不顯著(F=2.42,P> 0.05);Margalef豐富度指數不同人工林間差異極顯著(F=83.89,P< 0.01),季節間差異不顯著(F=0.39,P> 0.05)。分析顯示大型底棲動物Shannon-Wiener多樣性指數在珠海市淇澳島紅樹林區3種人工林間差異極顯著(F=35.30,P< 0.01),季節間差異也顯著(F=0.03,P< 0.05);Pielou均勻度指數不同人工林間差異極顯著(F=17.54,P< 0.01),季節間差異不顯著(F=0.10,P> 0.05);Margalef豐富度指數不同人工林間差異極顯著(F=27.28,P< 0.01),季節間差異不顯著(F=0.37,P> 0.05)。兩個研究地點的3種人工林中,1年無瓣海桑+木欖混交林和1年無瓣海桑+紅海欖混交林的Shannon-Wiener多樣性指數、Margalef豐富度指數和Pielou均勻度指數均大于同林齡無瓣海桑純林;而且均是1年無瓣海桑+木欖混交林的這3個物種多樣性指數最大,可見,在1年無瓣海桑林下混交木欖可最大幅度地增加林地大型底棲動物的物種多樣性。
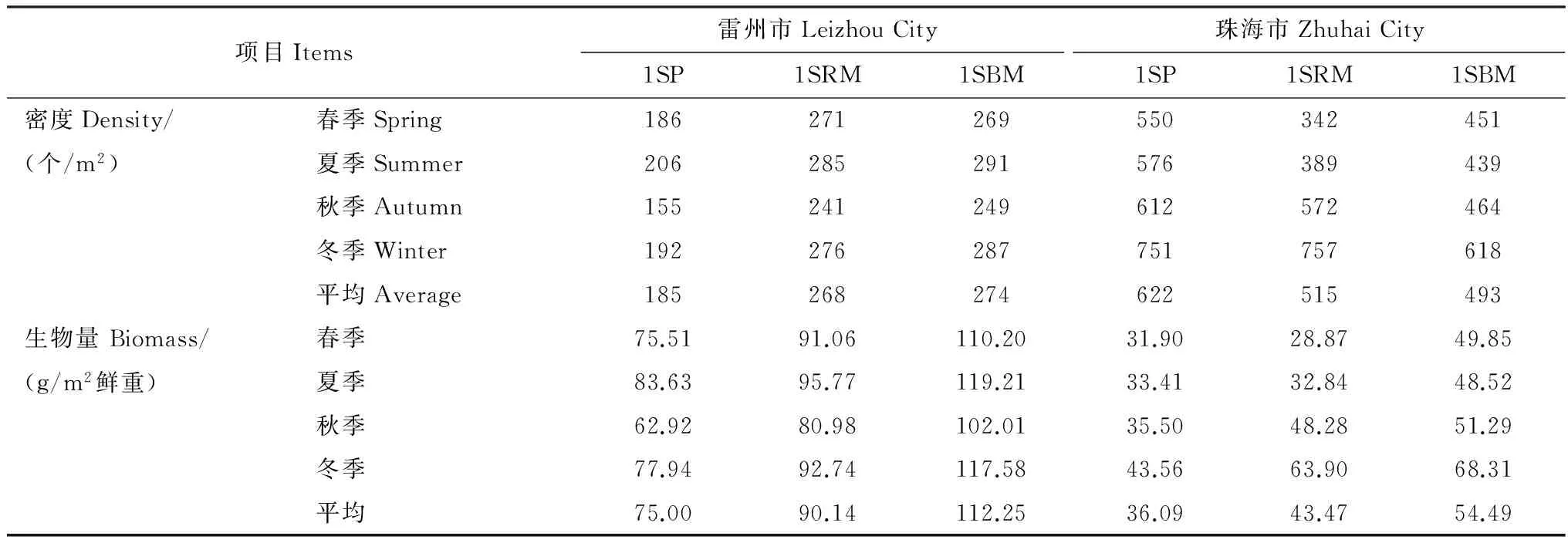
表4 兩個研究地點3種人工林大型底棲動物群落不同季節的密度和生物量Table 4 Density and biomass of the macrofauna communities in the three kinds of plantations across seasons in the two research sites

表5 兩個研究地點3種人工林大型底棲動物群落不同季節的物種多樣性指數Table 5 Species diversity indices of the macrofauna communities in the three kinds of plantations across seasons in the two research sites
2.5 兩個研究地3種人工林大型底棲動物群落結構
圖1中站點A1到C4取自雷州市附城鎮沿海紅樹林區,D1到F4取自珠海市淇澳島紅樹林自然保護區。聚類分析結果顯示:兩個研究地點各自的12個站點均分成了3個不同的組,可見,同一研究地點的3種人工林的大型底棲動物群落可以區分開來,說明不管是雷州市還是珠海市,在1年無瓣海桑林下混交紅海欖或木欖均會改變大型底棲動物群落結構。
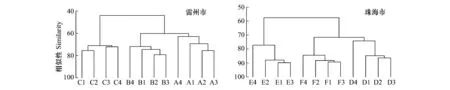
圖1 各站點大型底棲動物群落系統聚類圖Fig.1 Hierarchical cluster dendrogram of the macrofauna communities at the different sampling plots
兩個研究地各站點排序分析結果如圖2所示:雷州市的壓力系數(stress)=0.01,珠海市的壓力系數=0.07,兩個研究地的壓力系數均<0.1,說明兩個結果均支持聚類分析結果。這進一步證明了在1年無瓣海桑林下混交紅海欖或木欖可改變大型底棲動物群落的結構。
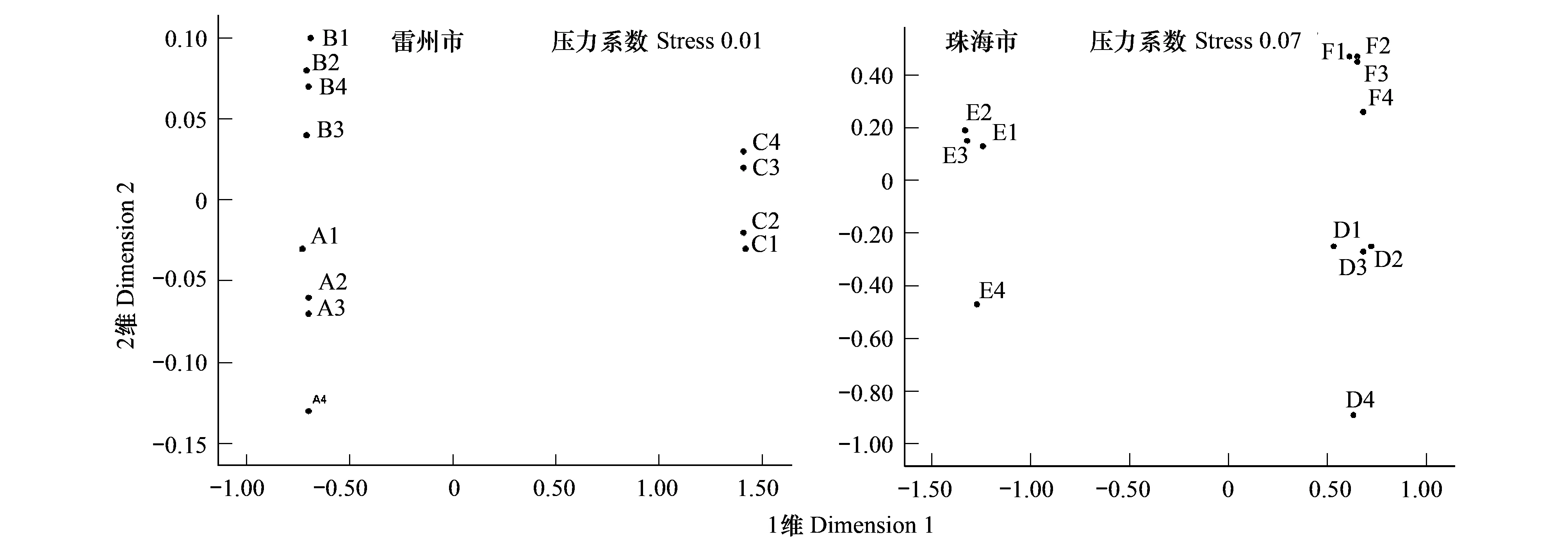
圖2 各站點大型底棲動物群落的非參數多變量標序Fig.2 MDS ordination of the macrofauna communities at the different sampling plots in the Leizhou City or Zhuhai City
用One-way ANOSIM檢驗雷州市附城紅樹林區3種人工林間底棲動物群落結構的差異,結果顯示差異極顯著(GlobalR=0.97,P=0.001)。用One-Way ANOSIM分別檢驗每2種人工林間動物群落結構的差異性,結果顯示:1年無瓣海桑純林和1年無瓣海桑+紅海欖混交林(GlobalR=0.87,P=0.029)、1年無瓣海桑純林和1年無瓣海桑+木欖混交林(GlobalR=1.00,P=0.029)、1年無瓣海桑+紅海欖混交林和1年無瓣海桑+木欖混交林(GlobalR=1.00,P=0.029)之間群落結構均有顯著的差異。采用群落之間相似性百分比分析(SIMPER)來了解表征群落和造成群落差異的物種,結果表明造成1年無瓣海桑純林和1年無瓣海桑+紅海欖混交林間群落結構差異的主要種(Contribution rate>3%)為:扁平擬閉口蟹、黑口擬濱螺、琵琶擬沼螺、紫游螺、螻姑蝦、褶痕相守蟹、短角圍沙蠶、寬身閉口蟹、等齒角沙蠶、瘤擬黑螺,它們對平均非度量系數(Bray-Curtis)不相似性的貢獻分別為:9.09%、9.00%、4.15%、4.08%、3.79%、3.65%、3.33%、3.24%、3.04%、3.00%;造成1年無瓣海桑純林和1年無瓣海桑+木欖混交林間動物群落結構差異的主要種(Contribution rate>3%)為:黑口擬濱螺、節織紋螺、悅目大眼蟹、厥目革囊星蟲、琵琶擬沼螺、珠帶擬蟹守螺、中國綠螂、膽形織紋螺、豆形短眼蟹,它們對平均非度量系數(Bray-Curtis)不相似性的貢獻分別為:5.45%、5.12%、4.46%、4.15%、3.74%、3.31%、3.18%、3.17%、3.03%;造成1年無瓣海桑+紅海欖混交林和1年無瓣海桑+木欖混交林間動物群落結構差異的主要種(Contribution rate>3%)為:節織紋螺、膽形織紋螺、扁平擬閉口蟹、寬身閉口蟹、珠帶擬蟹守螺、悅目大眼蟹、紅樹蜆,它們對平均非度量系數(Bray-Curtis)不相似性的貢獻分別為:4.62%、4.44%、3.80%、3.49%、3.43%、3.32%、3.04%。
同樣,用One-way ANOSIM檢驗珠海市淇澳島紅樹林區3種人工林間底棲動物群落結構的差異,結果顯示差異極顯著(GlobalR=0.944,P=0.001)。用One-Way ANOSIM分別檢驗每2種人工林間動物群落結構的差異性,結果顯示:1年無瓣海桑純林和1年無瓣海桑+紅海欖混交林(GlobalR=1.00,P=0.029)、1年無瓣海桑純林和1年無瓣海桑+木欖混交林(GlobalR=0.88,P=0.029)、1年無瓣海桑+紅海欖混交林和1年無瓣海桑+木欖混交林(GlobalR=1.00,P=0.029)之間群落結構均有顯著的差異。采用群落之間相似性百分比分析(SIMPER)來了解表征群落和造成群落差異的物種,結果表明造成1年無瓣海桑純林和1年無瓣海桑+紅海欖混交林間群落結構差異的主要種(Contribution rate>3%)為:悅目大眼蟹、雙齒相守蟹、小頭蟲、光滑狹口螺、短角圍沙蠶、德氏狹口螺、泥濘擬釘螺、無齒相守蟹、尖錐蟲,它們對平均非度量系數(Bray-Curtis)不相似性的貢獻分別為:9.43%、8.52%、7.21%、5.95%、5.89%、4.51%、4.48%、3.98%、3.13%;造成1年無瓣海桑純林和1年無瓣海桑+木欖混交林間動物群落結構差異的主要種(Contribution rate>3%)為:光滑狹口螺、細螯原足蟲、光滑河藍蛤、德氏狹口螺、雙齒相守蟹、褶痕相守蟹、日本刺沙蠶、長角涵螺,它們對平均非度量系數(Bray-Curtis)不相似性的貢獻分別為:22.76%、7.43%、5.31%、5.06%、4.48%、4.00%、3.86%、3.75%;造成1年無瓣海桑+紅海欖混交林和1年無瓣海桑+木欖混交林間動物群落結構差異的主要種(Contribution rate>3%)為:光滑狹口螺、悅目大眼蟹、小頭蟲、短角圍沙蠶、雙齒相守蟹、細螯原足蟲、泥濘擬釘螺、無齒相守蟹、德氏狹口螺、尖錐蟲,它們對平均非度量系數(Bray-Curtis)不相似性的貢獻分別為:11.39%、10.51%、7.28%、5.97%、5.04%、4.66%、4.60%、4.13%、3.50%、3.12%。可見,兩個研究地的3種人工林間底棲動物群落結構均存在差異。
2.6 兩個研究地大型底棲動物同環境各理化因子的關系
利用BIOENV分析,找出與各站點大型底棲動物豐度最為匹配的環境理化因子,結果見表6,不管是雷州還是珠海,1年無瓣海桑純林與1年無瓣海桑+紅海欖混交林的動物豐度均與紅海欖凋落物量、紅海欖幼苗成活率、紅海欖凋落物量占群落凋落物總量百分比相關;1年無瓣海桑純林與1年無瓣海桑+木欖混交林的動物豐度均與木欖凋落物量、木欖凋落物量占群落凋落物總量百分比、木欖幼苗成活率相關;而1年無瓣海桑+紅海欖混交林與1年無瓣海桑+木欖混交林的動物豐度均與木欖幼苗成活率、紅海欖幼苗成活率相關;RELATE檢驗結果均為顯著。

表6 大型底棲動物群落的豐度與環境理化因子的匹配關系Table 6 Multivariate correlation analysis of the relationship between the abundance of macrofauna communities and environmental parameters
3 討論
3.1 無瓣海桑與鄉土紅樹植物混交對林地大型底棲動物群落的影響
從本研究結果來看,不管是雷州市附城鎮還是珠海市淇澳島,3種人工林間大型底棲動物群落結構差異顯著(結果2.5),說明在1年無瓣海桑純林下種植紅海欖或木欖這2種鄉土紅樹植物,可改變林地大型底棲動物的群落結構。有研究證明紅樹林中的底棲動物優勢物種分布與紅樹林植被有密切關系[11],從表3可以看出,兩個研究地實驗結果均顯示無瓣海桑與這2種鄉土紅樹植物混交后,林地大型底棲動物優勢種明顯發生變化,而且,純林中物種優勢度指數最大的數值,在混交林中明顯下降,尤其是在1年無瓣海桑+木欖混交林中下降幅度最大。比如雷州市附城鎮1年無瓣海桑純林中,琵琶擬沼螺的物種優勢度指數最大,高達0.54,而在1年無瓣海桑+紅海欖混交林中其物種優勢度指數下降為0.27,到了1年無瓣海桑+木欖混交林則下降為0.21;珠海市淇澳島1年無瓣海桑純林中,光滑狹口螺的物種優勢度指數高達0.53,而在1年無瓣海桑+紅海欖混交林中其物種優勢度指數下降為0.42,到了1年無瓣海桑+木欖混交林物種優勢度指數已經不是最大了,而是細螯原足蟲,其物種優勢度指數為0.18。可見,在1年無瓣海桑純林下種植紅海欖或木欖這2種鄉土紅樹植物,在改變林地底棲動物群落結構的同時,還可降低林地最大物種優勢度指數數值,這是混交林大型底棲動物群落Pielou均勻度指數大于同林齡無瓣海桑純林的主要原因(表5)。
兩個研究地各站點大型底棲動物群落系統聚類結構均顯示按紅樹群落類型進行聚類。比如在雷州市附城鎮,取樣于1年無瓣海桑純林的 A1—C4聚為一組,而取樣于1年無瓣海桑+紅海欖混交林的B1—B4、取樣于1年無瓣海桑+木欖混交林的C1—C4也分別聚為一組。同樣,取樣于珠海市淇澳島紅樹林區1年無瓣海桑純林的 D1—D4、取樣于1年無瓣海桑+紅海欖混交林的E1—E4、取樣于1年無瓣海桑+木欖混交林的F1—F4都分別聚為一組(圖1)。兩個研究地各站點大型底棲動物群落的非參數多變量標序結果也顯示,取樣于不同人工林的站點可辨別開來,而且其壓力系數stress小于 0.1,說明標序結果支持聚類分析結果。這進一步證明了在1年無瓣海桑林下混交紅海欖或木欖可改變大型底棲動物群落的結構(圖2)。
影響大型底棲動物分布的環境因素包括海水鹽度、潮位和土壤特性等,但在小范圍區域,林內動物的分布更多地與紅樹林植被特性和潮位有關[12]。從表1可以看出,每個研究地的3種人工林間土壤理化因子差異不顯著,并且林地的上層木均為1年生的無瓣海桑,這說明大型底棲動物群落結構與紅海欖或木欖這些鄉土樹種的種植密切相關。有研究發現在泰國Ronang紅樹林中,相手蟹屬蟹類的分布與紅樹林土壤類型沒有密切的相關性[13],而與紅樹林植被的類型則有明顯的關系[14]。根據本研究結果,利用BIOENV分析找出與1年無瓣海桑純林和1年無瓣海桑+紅海欖混交林中大型底棲動物豐度最為匹配的環境理化因子,兩個研究地均包含紅海欖凋落物量、紅海欖凋落物量占群落凋落物總量百分比這2個因子;同樣分析1年無瓣海桑純林與1年無瓣海桑+木欖混交林中大型底棲動物豐度最為匹配的環境理化因子中,兩個研究地均包含木欖凋落物量、木欖凋落物量占群落凋落物總量百分比這2個因子;RELATE檢驗結果也均為顯著(表6)。這進一步說明大型底棲動物分布與紅海欖或木欖這些鄉土樹種的凋落物量密切相關。這也進一步證實了紅樹種植種類對紅樹林底棲動物的多樣性和分布的重要作用[3- 4,14- 19]。實際上,紅樹林除了為底棲動物提供棲息繁育場所外[20],還有一個重要的作用,就是直接或間接為其提供食物[21],而不同動物對不同紅樹植物殘體尤其是落葉啃食作用可能不同,即具有選擇性[18]。在香港米埔紅樹林的調查表明,兩個相距很近的秋茄林和白骨壤林內相手蟹的種類和數量有很大差異[14]。這可能是由于不同的蟹對不同紅樹植物凋落腐解葉在啃食上有不同的“選擇性”。最近國外的研究表明,某些蟹類和螺類對紅樹植物葉片的取食具有很強的選擇性,方蟹科種類Aratuspisonni對3種紅樹植物即大紅樹(Rhizophoramangle)、亮葉白骨壤(Avicenniagerminans)和假紅樹(Lagunculariaracemosa)葉片啃食的偏愛程度也不同[22]。螺類通過嗅覺感受到殘缺葉片所散發的氣味而攝食紅樹植物,不同種類和狀態的葉片因散發的氣味的差異而被螺類選擇性攝食[23]。國內李旭林等通過設置室內模擬飼喂實驗,研究了華南沿海紅樹林大型底棲動物的常見種無齒相手蟹(Sesarmadehaani)和雙齒相手蟹(S.bidens)對外來紅樹植物無瓣海桑和鄉土紅樹植物秋茄、桐花樹凋落葉片的取食特性。結果表明,兩種相手蟹種間的取食特性差異不顯著,但對不同樹種的取食上卻存在顯著差異,其原因是不同紅樹植物不同狀態的葉片在營養水平(如C/N比)和單寧含量上有差異[24]。從本研究結果來看,在無瓣海桑純林下種植紅海欖或木欖這些鄉土樹種,推測是增加了鄉土紅樹植物凋落物等“食物源”,促進了一些“偏好”鄉土紅樹植物凋落物的本地大型底棲動物的遷入、繁育和生長,改變了林地底棲動物優勢種,以及提高了底棲動物生物量和物種多樣性。
3.2 在無瓣海桑人工林下混交紅海欖或木欖的效果比較
兩個研究地的研究結果均顯示在無瓣海桑林下種植木欖的效果明顯優于紅海欖,表現在木欖的平均苗高、凋落物量、凋落物量占群落凋落物總量百分比和幼苗成活率均高于紅海欖(表1)。而且,1年無瓣海桑+木欖人工林中大型底棲動物生物量和物種多樣性明顯高于1年無瓣海桑+紅海欖人工林(表5)。利用BIOENV分析,找出與1年無瓣海桑+紅海欖混交林和1年無瓣海桑+木欖混交林中大型底棲動物豐度最為匹配的環境理化因子中,兩個研究地均包含木欖幼苗成活率、紅海欖幼苗成活率這兩個因子,而幼苗成活率高低直接關系到凋落物量的多少,這說明在1年無瓣海桑林下種植木欖的效果要優于紅海欖,推測是由于木欖幼苗成活率明顯高于紅海欖,從而為本地大型底棲動物提供了更多在營養成份、粗纖維和單寧含量以及 C/N 比等性質方面都“適合”的凋落物“食物源”[24]。至于紅海欖幼苗成活率低,可能與紅海欖不耐蔭有關[25];還可能與無瓣海桑對紅海欖幼苗的化感作用有關。李玫等以外來種無瓣海桑為供體,通過室內栽培實驗探討了無瓣海桑各器官水浸液的不同濃度對我國4個鄉土紅樹植物的化感作用,結果表明無瓣海桑對各鄉土紅樹植物均存在化感作用,所有水浸液均表現出高濃度下抑制幼苗生長,而且對不同受體紅樹植物的幼苗生長抑制強度不同[26]。是否無瓣海桑對紅海欖化感作用強于木欖,這種化感作用強度是否能導致紅海欖無法與無瓣海桑混交及長期效應研究都有待深入研究。
[1] 廖寶文, 李玫, 鄭松發, 陳玉軍, 鄭馨仁. 外來種無瓣海桑種內、種間競爭關系研究. 林業科學研究, 2003, 16(4): 418- 422.
[2] 彭友貴, 徐正春, 劉敏超. 外來紅樹植物無瓣海桑引種及其生態影響. 生態學報, 2012, 32(7): 2259- 2270.
[3] Tang Y J, Fang Z Q, Chen K, Zhang Z W, Zhong Y T, An D, Yang X B, Liao B W. Ecological influence of exotic plants ofSonneratiaapetalaon understory macrofauna. Acta Oceanologica Sinica, 2012, 31(5): 115- 125.
[4] 唐以杰, 方展強, 鐘燕婷, 張再旺, 陳康, 安東, 楊雄邦, 廖寶文. 不同生態恢復階段無瓣海桑人工林濕地中大型底棲動物群落的演替. 生態學報, 2012, 32(10): 3160- 3169.
[5] Ashton E C. Mangrove sesarmid crab feeding experiments inPeninsularmalaysia. Journal of Experimental and Marine Biology and Ecology, 2002, 273: 97- 119.
[6] Lee S Y.Tropical mangrove ecology: physical and biotic factors influencing ecosystem structure and function.Australian Journal of Ecology, 1999, 24(4): 355- 366.
[7] 許方宏, 吳曉東, 陳粵超, 潘偉生, 劉一鳴, 張葦. 雷州半島前沿惡劣潮間灘涂紅樹林造林樹種的選擇. 防護林科技, 2013, (6): 7- 9.
[8] 管偉, 廖寶文, 邱鳳英, 谷興華, 韓靜, 田廣紅, 楊雄邦. 利用無瓣海桑控制入侵種互花米草的初步研究. 林業科學研究, 2009, 22(4): 603- 607.
[9] 廖一波, 壽鹿, 曾江寧, 高愛根, 江志兵. 象山港不同養殖類型海域大型底棲動物群落比較研究. 生態學報, 2011, 31(3): 646- 653.
[10] Clarke K R, Gorley R N. PRIMER v5: User Manual/Tutorial. Plymouth: PRIMER-E Ltd., 2001: 91- 91.
[11] Nobbs M. Effects of vegetation differ among three species of fiddler crabs (Ucaspp.). Journal of Experimental Marine Biology and Ecology, 2003, 284(1- 2): 41- 50.
[12] 陳光程, 余丹, 葉勇, 陳彬. 紅樹林植被對大型底棲動物群落的影響. 生態學報, 2013, 33(2): 327- 336.
[13] Macnae W. A general account of the fauna and flora of mangrove swamps and forests in the Indo-West-Pacific region. Advance in Marine Biology, 1969, 6: 73-103, 104a-104a, 104b-104b, 105- 270.
[14] Lee C Y, Kwok P W. The importance of mangrove species association to the population biology of sesarmine crabsParasesarmaaffinisandPerisesarmabidens. Wetlands Ecology and Management, 2002, 10(3): 215- 226.
[15] 唐以杰, 余世孝. 廣東湛江紅樹林保護區大型底棲動物群落的空間分帶. 生態學報, 2007, 27(5): 1703- 1714.
[16] Lee S Y. The ecological role of grapsid crabs in mangrove ecosystems: implications for conservation. Marine and Freshwater Research, 1998, 49(4): 335- 343.
[17] Roach A C, Lim R P. Variation in the population dynamics of the intertidal pulmonate gastropodSalinatorsolidaMartens (Gastropoda: Amphibolidae) at Towra Point, NSW, Australia. Wetlands Ecology and Management, 2000, 8(1): 53- 69.
[18] 葉勇, 翁勁, 盧昌義, 陳光程. 紅樹林生物多樣性恢復. 生態學報, 2006, 26(4): 1243- 1250.
[19] 魏德重, 項長友, 鮑毅新, 李良峰, 章書聲. 紅樹林種植對大型底棲動物群落結構及功能群的影響. 浙江師范大學學報: 自然科學版, 2012, 35(2): 195- 202.
[20] Robertson A I, Duke N C. Mangroves as nursery sites: comparisons of the abundance of fish and crustaceans in the mangroves and other nearshore habitats in tropical Australia. Marine Biology, 1987, 96(2): 193- 205.
[21] Lee S Y. Potential trophic importance of the faecal material of the mangrove Sesarmine crabSesarmamessa. Marine Ecology Progress Series, 1997, 159: 275- 284.
[22] Erickson A A, Saltis M, Bell S S, Dawes C J. Herbivore feeding preferences as measured by leaf damage and stomatal ingestion: a mangrove crab example. Journal of Experimental and Marine Biology and Ecology, 2003, 289(1): 123- 138.
[23] Fratini S, Cannicci S, Vannini M. Feeding clusters and olfaction in the mangrove snailTerebraliapalustris(Linnaeus) (Potamididae: Gastropoda). Journal of Experimental and Marine Biology and Ecology, 2001, 261(2): 173- 183.
[24] 李旭林, 彭逸生, 萬如, 伍卡蘭, 陳桂珠. 兩種相手蟹對不同紅樹植物葉片取食的偏好性. 生態學報, 2010, 30(14): 3752- 3759.
[25] 曾雯珺, 廖寶文, 陳先仁, 粟娟, 馬蘇全, 管偉. 無瓣海桑與三種鄉土紅樹植物混交的生態效應. 生態科學, 2008, 27(1): 31- 37.
[26] 李玫, 廖寶文, 鄭松發, 陳玉軍. 外來種無瓣海桑化感作用研究初報. 生態科學, 2002, 21(3): 197- 200.
Effects of mixed plantations ofSonneratiaapetalaand indigenous mangrove species on understory macrofauna
TANG Yijie1, FANG Zhanqiang2,*, HE Qing1, WU Yingmin1, LU Di1, LI Yanni1, TAN Lijun1, YANG Xuebin1, CHEN Siming1
1DepartmentofBiology,GuangdongUniversityofEducation,Guangzhou510303,China2CollegeofLifeSciences,SouthChinaNormalUniversity,KeyLaboratoryofEcologyandEnvironmentalScienceinGuangdongHigherEducation,Guangzhou510631,China
Sonneratiaapetalawas introduced to China′s Hainan Island from Bangladesh in 1985. In recent years, because of its adaptability and rapid growth,S.apetalahas become the most important species to ensure the forestation of mangroves that are imported to the coastal and intertidal regions of South China. This study was conducted to explore the ecological responses of understory macrofauna communities during the transformation of a pureS.apetalaplantation and the major environmental factors affecting them. Two pure plantations of 1-year-oldS.apetalawere chosen as the study sites: one was at Zhanjiang Island Mangrove Nature Reserve in Leizhou, and the other was at Qi′ao Island Mangrove Nature Reserve in Zhuhai, Guangdong Province, China. Indigenous mangrove seedlings ofRhizophorastylosaorBruguieragymnorrhizawere mixed withS.apetalaby underwood planting. The macrofauna communities of the transformed 1-year-oldS.apetala+R.stylosaand 1-year-oldS.apetala+B.gymnorrhizamixed plantations were compared with those of a similarly aged pureS.apetalaplantation. The results showed differences in the dominant species among the macrofauna communities in the 1-year-old pureS.apetalaplantation and the 1-year-oldS.apetala+R.stylosaand 1-year-oldS.apetala+B.gymnorrhizamixed plantations. The results of a one-way analysis of similarity, hierarchical clustering, and nonmetric multidimensional scaling all showed significant structural differences among the macrofauna communities of the pure plantations and mixed plantations, which had similar forest ages. In addition, the results showed that transforming pureS.apetalaplantations by underwood planting with seedlings ofR.stylosaorB.gymnorrhizamay improve the biomass and species diversity of the understory macrofauna. BIOENV analysis showed that macrofauna distribution was closely related to the amount of indigenous mangrove plant litter ofR.stylosaorB.gymnorrhiza, which further confirmed that underwood planting with indigenous mangrove species played an important role in the distribution and species diversity of the understory macrofauna. Because the levels of the physicochemical factors in the soil in the three forest types were not significantly different and the upper canopy of these forests was entirely comprised ofS.apetala, it can be speculated that underwood planting withR.stylosaorB.gymnorrhizaincreased the understory indigenous mangrove plant litter, the nutrient levels (such as the C/N ratio), and the tannin content that may be adaptive or preferred by indigenous macrofauna. Furthermore, some mobile mollusks and crustaceans move to mangrove habitats that are more suitable for survival, leading to a rise in the biomass and species diversity of understory macrofauna in mixed plantations. However, experimental results from the two research sites showed that the effect of underwood planting in pureS.apetalaplantations withB.gymnorrhizawas superior to that withR.stylosabecause the average seedling height, litter production, percentage of litter production accounting for the communities′ total litter production, and the survival rate of seedlings were all greater for underwood planting withB.gymnorrhizathan withR.stylosa. This also resulted in a higher biomass increase and species diversity of macrofauna in the underwood planting withB.gymnorrhizathan withR.stylosa. This experiment was conducted 3 months after underwood planting withR.stylosaorB.gymnorrhizain the pureS.apetalaplantations. Since the total research period was only 1 year, this study only explored the short-term effects of the experiment; therefore, future studies on its long-term effects are required.
Sonneratiaapetala;Bruguieragymnorrhiza;Rhizophorastylosa;macrofauna;species diversity
廣東省科技計劃項目(NO.2009B030600006); 國家科技支撐計劃項目(NO.2009BADB2B0401-02); 國家星火計劃(2013GA780019); 廣東第二師范學院大學生創新創業訓練計劃項目(1427813116)(142781429); 廣東第二師范學院校級科研項目(2013yjxm03)
2014- 03- 23;
日期:2015- 04- 20
10.5846/stxb201403230515
*通訊作者Corresponding author.E-mail: fangzhq@scnu.edu.cn
唐以杰,方展強,何清,吳映明,陸棣,李燕妮,譚莉君,楊學彬,陳思敏.無瓣海桑與鄉土紅樹植物混交對林地大型底棲動物的影響.生態學報,2015,35(22):7355- 7366.
Tang Y J, Fang Z Q, He Q, Wu Y M, Lu D, Li Y N, Tan L J, Yang X B, Chen S M.Effects of mixed plantations ofSonneratiaapetalaand indigenous mangrove species on understory macrofauna.Acta Ecologica Sinica,2015,35(22):7355- 7366.
猜你喜歡
歌海(2024年6期)2024-03-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
幼兒園(2021年4期)2021-07-28 07:38:04
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
歌海(2018年4期)2018-05-14 12:46:15
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
