荒漠草原不同植物根際與非根際土壤養分及微生物量分布特征
2015-02-05 06:15:00劉秉儒
生態學報 2015年22期
關鍵詞:植物
楊 陽, 劉秉儒
寧夏大學, 西北退化生態系統恢復與重建教育部重點實驗室, 銀川 750021
荒漠草原不同植物根際與非根際土壤養分及微生物量分布特征
楊 陽, 劉秉儒*
寧夏大學, 西北退化生態系統恢復與重建教育部重點實驗室, 銀川 750021
通過對寧夏荒漠草原6種地帶性優勢物種長芒草、蒙古冰草、甘草、牛心樸子、黑沙蒿和苦豆子植物根際與非根際土壤養分和微生物量分布特征進行研究,探討不同植物根際養分的富集的相關性和差異性。研究結果表明:6種植物根際土壤養分和微生物量均表現出明顯的富集效應,根際富集率大小依次為菊科(黑沙蒿)>豆科(苦豆子、甘草)>禾本科(長芒草、蒙古冰草)>蘿藦科(牛心樸子);全磷(TP)在根際和非根際中無顯著差異(P>0.05),其它土壤養分及理化指標在根際中均表現出顯著富集(P<0.05),土壤養分中以有機碳(SOC)的富集作用最為明顯;土壤有效態養分較全量養分對植物根際微小的變化響應更為靈敏;不同荒漠植物根際與非根際SOC與全氮(TN)呈極顯著線性關系(P< 0. 01),TN與堿解氮之間呈極顯著線性關系(P<0.01),TP與有效磷(AP)沒有顯著的相關性(P>0.05)。荒漠植物土壤有效養分在根際存在一定的富集,灌木和豆科植物的根際效應的大于禾本科植物,它們通過降低根際pH值可以提高根際養分,有利于在脆弱環境下對土壤養分的有效利用。
荒漠草原;植物根際;土壤養分;土壤微生物量;空間分布
德國微生物學家Lorenz Hiltner將根際土(Rhizosphere Soil)定義為根系周圍、受根系生長影響并且能夠從微環境中吸收大量養分的土體[1- 3]。根際土是圍繞根系進行生物地球化學循環最活躍的區域,是土壤-植物根系-微生物三者相互作用的場所和各種物質循環和能量流動的門戶,對生態系統養分動態分布與循環、植物種間作用等發揮重要作用[3- 6],根系誘導產生根際土壤養分的變化已被證實[1- 3]。由于根際是土壤-植物根系-微生物生態系統物質交換的活躍界面,其分泌物、微生物活動以及水分的差異具有較大差異,根際過程對于認識植物對土壤中養分的利用及適應顯得非常重要。
荒漠區土壤養分含量較貧瘠,這一區域植物“根際效應”及根際對養分的截留效應較為明顯[7- 9]。荒漠植物根際土壤特征可能是有效吸收、利用土壤養分和適應脆弱環境的最直接表征之一,而荒漠根際土壤特征及對環境的適應中所起的作用等科學問題尚未得到充分的認識[10- 12]。國內對根際的研究開始于80年代初,大部分集中在農作物和森林等生態系統和對農作物和樹木的根際養分、毒害離子、根際微生物、根際分泌物、簇根等方面,而對于干旱、養分貧瘠、氣候脆弱的荒漠植物根際土壤養分、微生物量以及根際在土壤-根際-植物系統中的作用還少見報道[13- 15]。研究荒漠草原不同植物根際與非根際土壤養分分布特征與動態變化及其根際養分的富集作用,對于揭示荒漠植物對土壤養分的保護、有效利用機制及其演替、恢復、重建等具有重要生態學意義[1,5- 7,13- 15]。鑒于此,本文研究了荒漠草原6種不同植物根際土壤養分、理化因子和微生物量等指標,旨在探明荒漠植物根際養分的差異性以及對土壤養分利用對策和保護效應,以期揭示荒漠植物對干旱、貧瘠、脆弱環境的適應機理和策略及其在土壤-植物恢復中的作用。
1 材料與方法
1.1 研究區自然概況
研究區位于寧夏中東部鹽池縣毛烏素沙地南緣和黃土高原過渡帶之間,屬干中溫帶半干旱區、歐亞草原區、中部草原區的過度地帶,是典型的鄂爾多斯臺地,該區屬于典型的大陸性氣候,熱量豐富,年降水量在150—450 mm之間,主要集中7—9月,約占全年的60%以上,年蒸發量在1221.9—2086.5 mm。土壤類型主要是灰鈣土,其次是風沙土、黃綿土,腐殖質層較薄,土壤結構松散,肥力低下,多風沙,植被成分比較單一,主要由禾本科、菊科、百合科、藜科、蒺藜科、豆科、十字花科和鳶尾科等植物組成;植被以旱生多年生草本植物為主,以長芒草(Stipabungeana)、蒙古冰草(AgropyronmongolicumKeng)、甘草(GlycyrrhizauralensisFisch.)、牛心樸子(CynanchumkomaroviiAl.)、黑沙蒿(Artemisiaordosica)和苦豆子(Sophoraalopecuroides)植物為主要建群種的天然草地類型(典型無干擾情況下),具有典型和代表性,占該區域草地面積的80%以上。
1.2 樣地設置與樣品采集
于2013年8月中旬對每種類型的草地在3個不同的區域布設面積為100 m×100 m的樣地,作為3個重復,在每個樣地內隨機選取典型植物,按照其長勢依次分為大、中、小各3株共27株,采樣植物的形態特征和生長狀況列于表1。
不同植物根際土壤樣品采集采用Riley和Barber的抖落法[16- 17]。在上述所選的典型樣地內挖取具有完整根系的土體(根系主要分布的范圍),先輕輕抖落大塊不含根系的土壤,用小刀取下附在根系周圍的土壤為非根際土(S),裝入塑料袋內混勻,然后用刷子刷下粘附在根圍的土壤(距離根圍0—5 mm)作為根際土(R),取根際土盡量減少損害植物根系,對于混雜于根際土中的根系要徹底去除。每個樣地各分別隨機采集3株大、中、小植物根際、非根際土壤(排除植物個體大小對根際的影響),每種植物采集27株,總共采集到植物162株和324個土樣(根際和非根際土),剛采集的新鮮土樣塑封袋盛裝在保溫箱內4℃保存以測定土壤微生物量及含水量,然后風干去除有機碎片,過2 mm篩用以測定土壤理化指標及養分。
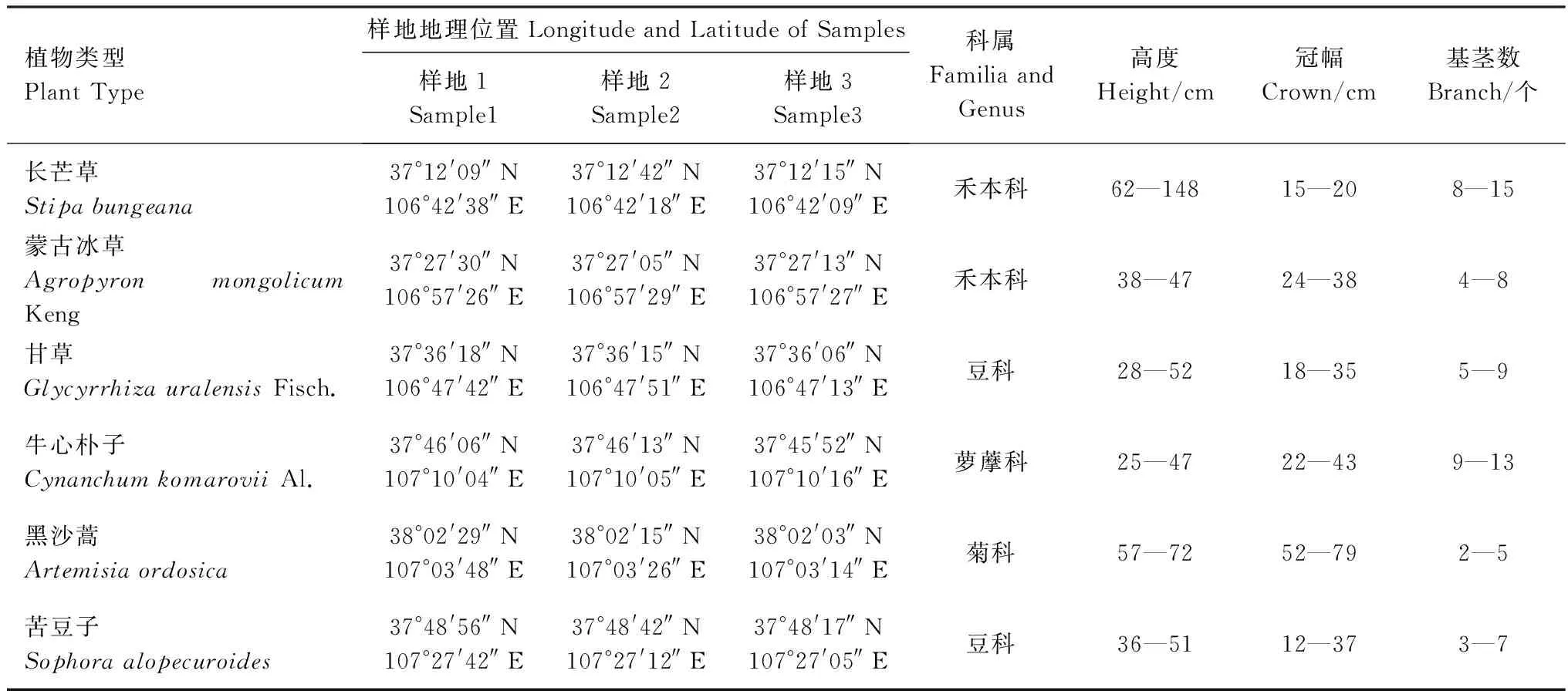
表1 荒漠草原不同植物生長特點Table 1 The basic characteristics of different plants and environmental condition in desert steppe
1.3 測定方法
1.3.1 土壤理化性質的測定
土壤含水量采用烘干法測定;土壤pH采用電極電位法測定(1∶2.5土水比浸提液);電導率EC采樣P4多功能測定儀(Multiline P4 Universal Meter, WTM公司,Germany)測定;土樣研磨過0.5 mm篩用于SOC、TN和TP的分析,SOC采用重鉻酸鉀氧化-外加熱法測定;TP采用NaOH堿溶-鉬銻抗比色法測定(日產UV-1601分光光度計);有效磷采用NaHCO3浸提-鉬銻抗比色法測定;TN采用半微量凱氏定氮法測定(意大利產DK6,UDK140分析儀);堿解氮采用NaOH-H3BO3法測定[18-19]。
1.3.2 土壤微生物量碳、氮的測定
土壤微生物量碳、氮采用氯仿熏蒸-K2SO4浸提法,其中氯仿熏蒸殺死的微生物體中的碳、氮被浸提出來的比例分別為0. 38、0. 45[20]。
1.4 數據處理
采用SPSS 18.00進行方差分析;Excel 2003.00進行繪圖;多重比較采用LSD法;單因素方差檢驗根際與非根際土壤是否存在顯著差異;所有數據為3次重復的平均值,Pearson相關分析討論土壤理化因子與微生物量之間的相關關系。
根際富集率E(Enrichment ratio)反映養分的富集程度,E值的大小反映土壤性狀值在植物根際的富集程度,同時也反映了植物根際效應的強弱,計算公式如下[14- 15,21]:
E=[(根際含量-非根際含量)/非根際含量 ]×100%
2 結果與分析
2.1 荒漠草原植物根際與非根際土壤養分平均值和富集率
荒漠草原植物根系不斷從外界環境吸收養分,不同的吸收速率使根際養分出現虧缺和富集。從表2可以看出,荒漠草原植物根際與非根際土壤微生物量碳、微生物量氮、有機碳、全氮、堿解氮、有效磷、含水量及pH呈現出顯著的差異(P<0.05),而土壤全磷和電導率沒有明顯的差異(P>0.05)。相對于非根際土壤,根際MBC平均高出30.3%,MBN平均高出72.8%,SOC平均高出43.6%,TN平均高出63.3%,TP平均高出45.7%,TP平均高出41.3%,堿解氮平均高出68.1%,電導率平均高出3.4%,含水量平均高出25.0%,由于根系呼吸釋放CO2、分泌有機酸、微生物活動產生有機酸和CO2等作用導致根際酸化,而根際pH值的降低能夠提高多種礦質養分的有效性和促進養分有效性的轉化和活化,從而保證了其對微量元素的吸收和自身的生長;綜合來看,荒漠植物根際對土壤養分表現出明顯的富集和截存效應。

表2 荒漠草原植物根際與非根際土壤養分平均值和富集率Table 2 Contents of nutrients and enrichment ratio in rhizosphere and non-rhizosphere soil of different desert plant
2.2 荒漠草原植物根際和非根際土壤養分含量變化
荒漠草原6種植物根際SOC富集率大小表現為:黑沙蒿>甘草>苦豆子>牛心樸子>長芒草>蒙古冰草,受豆科固氮作用影響,土壤TN富集率大小表現為:甘草>黑沙蒿>苦豆子>牛心樸子>蒙古冰草>長芒草,土壤TP富集率大小表現為:甘草>黑沙蒿>蒙古冰草>牛心樸子>長芒草>苦豆子,整體上來看(表3),灌叢(黑沙蒿)和豆科植物(苦豆子、甘草)對土壤各養分的富集率均顯著高于禾本科植物(長芒草、蒙古冰草)。
土壤有效成分中,黑沙蒿、苦豆子、甘草和牛心樸子根際中有效磷顯著富集,而在長芒草和蒙古冰草根際中有效磷含量虧缺顯著;除了牛心樸子根際土壤堿解氮含量低于非根際,其他5種植物根際土均表現明顯的富集效應,黑沙蒿、苦豆子和甘草根際土壤堿解氮富集率明顯高于長芒草和蒙古冰草;6種荒漠植物根際土壤pH值均小于非根際,其降低幅度在0.10—0.82個單位之間,降低幅度大小順序為:黑沙蒿>苦豆子>牛心樸子>甘草>長芒草>蒙古冰草,其中黑沙蒿根際pH值最小并且下降幅度最大,達0.82個單位,這說明黑沙蒿比草本植物更容易引起根際土壤 pH值的降低;長芒草、蒙古冰草、甘草和牛心樸子根際電導率顯著富集, 而黑沙蒿和苦豆子根際中電導率虧缺,虧缺率為12.79%和6.52%;含水量在6種荒漠植物中顯著富集,并且黑沙蒿、苦豆子和甘草富集率均顯著高于其他3種植物。
由圖1可知,6種植物根際MBC和MBN含量均高于非根際,其中黑沙蒿、苦豆子和甘草根際MBC極顯著富集(P<0.01),長芒草根際MBC顯著富集(P<0.05),蒙古冰草和牛心樸子根際MBC并沒有達到顯著富集(P>0.05);除了長芒草根際MBN沒有達到顯著富集(P>0.05),其他5種植物根際MBN均達到極顯著富集(P<0.01)。綜合來看,黑沙蒿、苦豆子和甘草根際各成分富集率顯著高于其他3種植物。
2.3 荒漠草原植物根際與非根際土壤養分、微生物量相關分析
由表4可知,土壤根際MBC與MBN、SOC、TN和含水量呈顯著或極顯著正相關(P<0.01),與電導率呈極顯著負相關(P<0.01);MBN與SOC、TN和含水量呈極顯著正相關(P<0.01),與電導率呈顯著負相關(P<0.05);SOC與TN和含水量呈極顯著正相關(P<0.01),與電導率呈極顯著負相關(P<0.01);TN與TP、堿
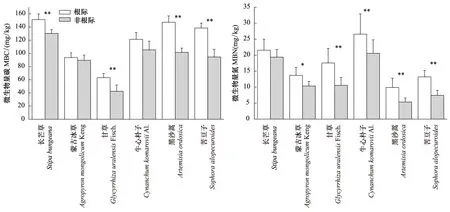
圖1 荒漠草原不同植物根際和非根際土壤微生物量Fig.1 Contents of soil microbial biomass in rhizosphere and non-rhizosphere soil of different desert plant
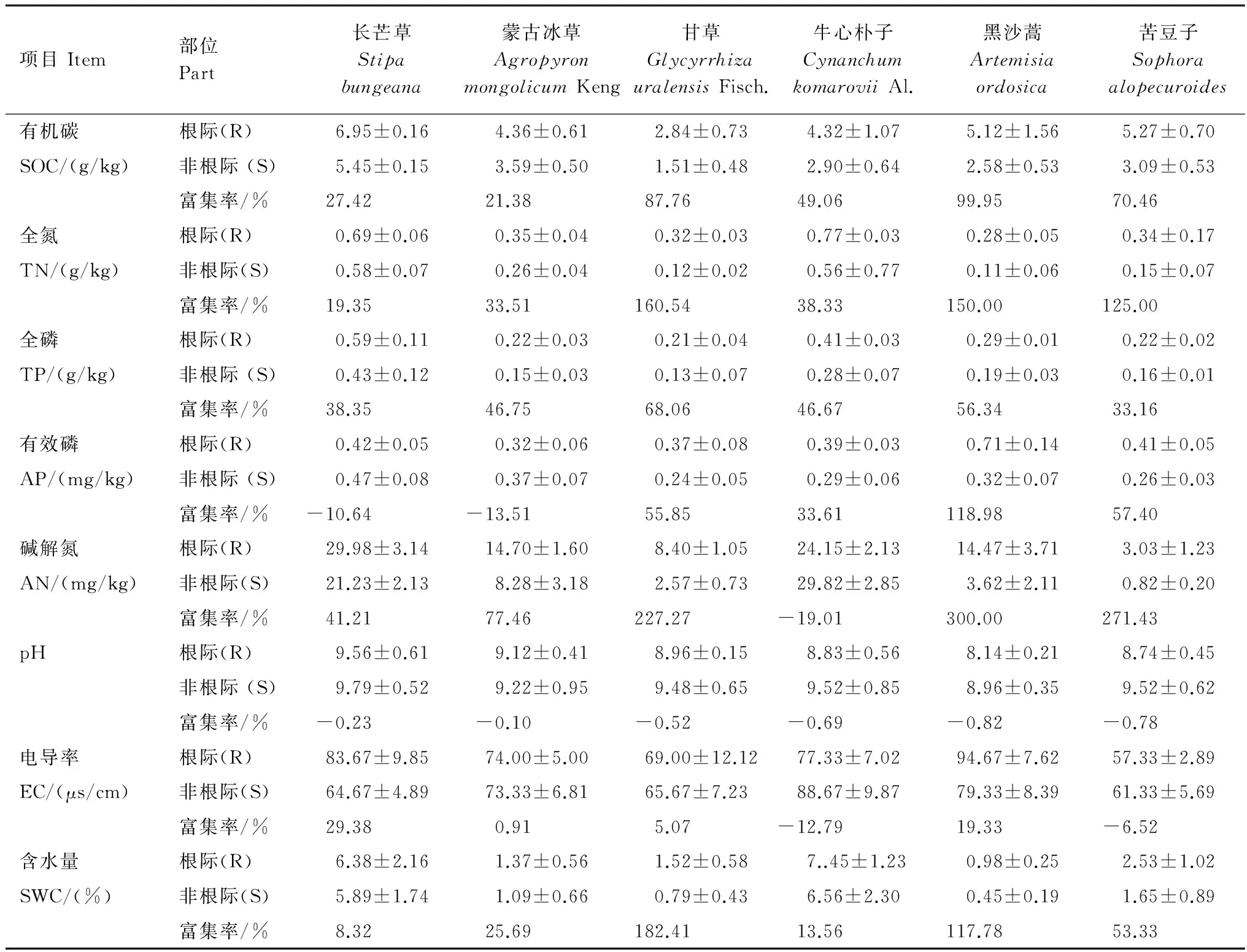
表3 荒漠草原植物根際和非根際土壤養分含量變化Table 3 Contents of soil nutrients in rhizosphere and non-rhizosphere soil of different desert plant
解氮和含水量呈極顯著正相關(P<0.01),與有效磷呈極顯著負相關(P<0.01);TP與堿解氮、pH呈極顯著正相關(P<0.01),與含水量呈顯著正相關(P<0.05);堿解氮與電導率呈顯著正相關(P<0.05);pH與電導率呈顯著正相關(P<0.05);以上結果顯示了根際SOC和TN在荒漠生態系統中相互影響和制約。
由表5可知,土壤非根際MBC與MBN、SOC、TP和含水量呈顯著或極顯著正相關,與電導率呈極顯著負相關(P<0.01);MBN與SOC、TN、含水量和有效磷呈顯著或極顯著正相關,與電導率極顯著負相關(P<0.05);SOC與TN和含水量呈極顯著正相關(P<0.01),與電導率呈顯著負相關(P<0.05);TN與TP、堿解氮和含水量呈極顯著正相關(P<0.01);TP與堿解氮、pH和含水量呈正相關;堿解氮與電導率和含水量呈顯著正相關(P<0.01);以上結果顯示了根際與非根際土壤養分、微生物量變化規律、影響因子之間的相關性。

表4 荒漠草原植物根際土壤養分影響因子Pearson相關性分析Table 4 Pearson′s correlation coefficients among soil nutrient factors in rhizosphere soil of different desert plant
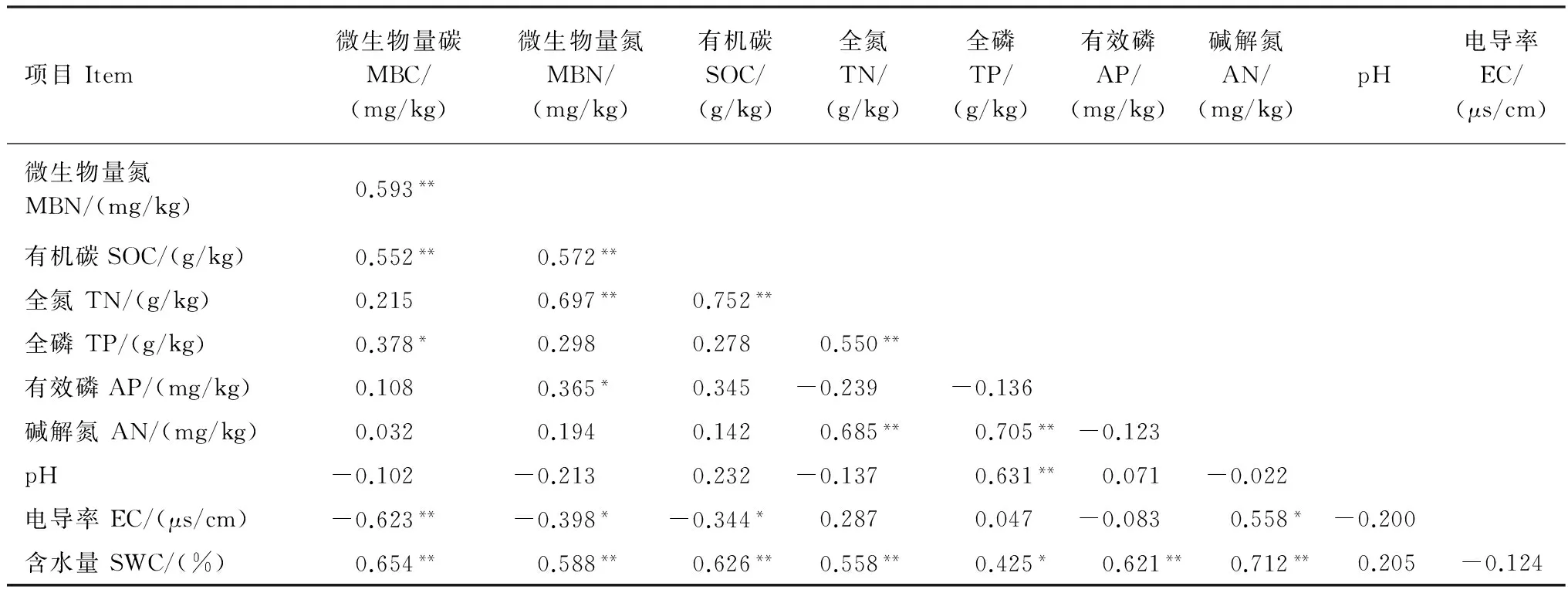
表5 荒漠草原植物非根際土壤養分影響因子Pearson相關性分析Table 5 Pearson’s correlation coefficients among soil nutrient factors in non-rhizosphere soil of different desert plant
由原始數據擬合得到的回歸關系經統計學檢驗得到擬合度參數R2,并在P<0.05水平檢驗相關系數的顯著性,F檢驗結果表明,下表中的線性回歸關系均達到極顯著水平,各回歸關系成立,得以下結果。由表6可知,荒漠草原植物根際和非根際SOC與TN均呈極顯著線性相關(P<0.01);TN與堿解氮在根際與非根際呈顯著線性相關(P<0.01);而TP與有效磷在根際和非根際沒有顯著相關性(P<0.01);MBC與MBN在根際與非根際呈顯著線性相關(P<0.01)。TP和有效磷之間并沒有表現出相關性(P<0.01),說明在荒漠植物凋落物歸還的P不足以維持植物需求,土壤磷庫中P素不斷轉移到地上植物體內。

表6 荒漠草原植物根際養分與非根際養分的線性回歸關系Table 6 The relations of linear regression in rhizosphere and non-rhizosphere soil nutrients of different desert plants
3 討論與結論
3.1 荒漠植物根際與非根際土壤養分和富集率分析
植物根際研究文獻表明,各種元素含量在根際與非根際土壤中存在較大差異[22- 24]。本研究中不同荒漠植物根際與非根際土壤養分、微生物量及理化性質等均存在明顯差異(表2),主要是由于植物根系不斷從外界攝取養分及其分泌物向根表的遷移,使得根際微區域內的養分形態、濃度和分布等方面較為復雜[13- 15,21,25],這充分體現了荒漠植物在定居和發育過程中對養分的需求變化[21,26]。由表1和2可知,以黑沙蒿根際對土壤各養分的富集率最大,苦豆子和甘草次之,黑沙蒿灌叢適應脆弱環境需要更多的營養維持根系的生長和發育,因而出現更多的根系沉淀和養分富集[13- 15,21,25- 26];甘草和苦豆子為豆科植物,根瘤菌有利于根系吸收營養物質,因此根際養分含量相對偏高;蒙古冰草和牛心樸子屬于矮小草本植物,根系及枯落物對土壤養分的吸收貢獻較小。綜合比較可知,黑沙蒿(灌叢)根際富集率明顯優于其他草本植物,依次表現為菊科(黑沙蒿)>豆科(苦豆子、甘草)>禾本科(長芒草、蒙古冰草)>蘿藦科(牛心樸子),主要是由于黑沙蒿灌叢根系發達、生物量大、枯落物豐富,伴有養分富集的“肥島效應”和種內營養補償效應[27- 29],促進了根際微生物的旺盛生長,因此表現出較為明顯的根際效應[7- 9,13- 15,21,30]。從該結果可以看出,黑沙蒿灌叢可作為改良土壤、防風固沙及退化荒漠生態區植物恢復與重建關鍵物種。
3.2 不同荒漠植物根際與非根際養分比較分析
表1和表2表明6種荒漠植物根際養分均表現明顯的富集作用,其中以SOC的富集作用最為明顯,黑沙蒿灌叢根際對SOC的富集作用遠高于其他草本植物,可能是由于黑沙蒿灌叢對土壤風蝕物質、降塵和凋落物的截獲,通過發達根系以根際沉積的形式顯著聚集于根際[13- 15,21],此外,植物根系可通過一系列措施來減少根際碳損失、緩解環境的脅迫,也能通過地下根系的可塑性分配適應土壤養分有效性的改變[30- 31]。堿解氮在牛心樸子根際虧缺可能是由于牛心樸子生長對N素的消耗、以及根系生理代謝功能強、根際微生物活動旺盛等促進了N素向堿解氮的轉化,這種現象是荒漠植物對N素缺乏的一種特殊適應策略;其他5種植物N素營養利用上不同于牛心樸子,以苦豆子和甘草的根際N素富集作用最大,其根系與根瘤菌共生可能促進了N素的固定和營養的吸收。而土壤TP在根際和非根際中差異并不顯著,但黑沙蒿灌叢的根際富集效果較為明顯,說明其根系具有較強的吸附P素能力,由表4可知荒漠植物根際TP與pH呈極顯著的正相關關系(P<0.01),表明根系分泌的有機酸對難溶性P起到一定的活化作用[4,32- 33],pH值的降低能夠提高P素的轉化與活化,以此抵御脆弱環境和土壤養分貧瘠等[1- 3,5- 7]。由于黑沙蒿強大根系呼吸釋放CO2、分泌有機酸能力,造成其根際土壤pH值均低于其他草本植物。綜上所述,本研究初步表明荒漠植物土壤有效養分在根際存在一定的富集,它們通過降低根際pH值可以提高根際養分,有利于在脆弱環境下對土壤養分的有效利用。
由于根系分泌物種類和數量繁多、影響因素相對復雜,不同分泌物之間可能存在相互替代和拮抗作用等,使得植物根系分泌物對土壤養分有效性有多重影響[34- 36]。本研究荒漠植物根際土壤有效養分(堿解氮和速效磷)的變化幅度高于全量養分,說明荒漠植物在土壤養分循環和吸收過程中,有效態養分較全量養分對植物根際微小的變化響應更為靈敏。其中長芒草和蒙古冰草有效磷在根際中虧缺可能是由于二者生長過程中對P素的需求量大,并且對P素的吸附和緩沖能力較弱等。因此,保持土壤中較低的有效磷可能是土壤P素持續利用的有效對策。
3.3 不同荒漠植物根際與非根際微生物量比較分析
荒漠植物根際MBC和MBN均比非根際高,根系較為龐大的植物,其土壤微生物量也較高。由圖2可知,黑沙蒿、苦豆子和甘草深根系植物根際微生物量富集作用高于蒙古冰草、長芒草和牛心樸子淺根系植物,與前人的研究結果一致[37- 39],蒙古冰草、長芒草和牛心樸子土壤微生物量較低,原因可能與它們發育階段的物質分配有關,地上積累的物質大多用于發育繁殖器官,地下生物量表現負累積。雖然MBC和MBN只占SOC、TN的3%左右,但它們是植物養分轉化、SOC代謝及降解的驅動力。此外,MBC和MBN差異主要與植物種群、生物學特性、土壤理化特性、凋落物質量和數量以及生長節律有關,是各種因素綜合作用的結果,還存在較多的不確定性因素[37- 39]。
3.4 荒漠草原植物根際與非根際土壤養分、微生物量的相關分析
由表4和5可知,荒漠植物根際、非根際MBC與MBN之間達到顯著或極顯著相關性,MBC、MBN與SOC、TN、堿解氮和有效磷等土壤養分之間相關性也達到顯著或極顯著水平,表明MBC和MBN具有協同性并且可以作為判斷荒漠草原土壤肥力性狀的生物學指標,這與許多學者的研究結果基本一致[37- 39]。SOC主要來源于根系分泌物和凋落物等,N素除微生物固氮或少量大氣沉降外,大多來源于植物凋落物的返還、死亡的細根和根系分泌的有機物[1- 3,13- 15],因此根際和非根際土壤TN與SOC之間呈現出極顯著相關性;本研究表明荒漠植物根際和非根際土壤養分、微生物量變化趨勢基本保持一致,它們的主導控制因子也基本相同。荒漠植物根際沉積不僅為根際微生物提供了豐富的C源,還對根際土壤養分產生重大影響[13- 15,21,25- 26,30]。因此,認識和調控根際沉積及養分的流量和方向對于建立根際環境中養分的合理分配模式具有重要作用。
致謝:以色列巴伊蘭大學劉任濤博士幫助寫作,宋乃平教授、楊新國副教授在數據測試過程中提供幫助,特此致謝。
[1] Ma B, Zhou Z Y, Zhang C P, Li X R. The character of phosphorus concentrations in rhizosphere soil of super-xerophytic shrubs. Acta Prataculturae Sinica, 2005, 14(3): 106- 110.
[2] Barth R C, Klemmedson J O. Shrub-induced spatial patterns of dry matter, nitrogen, and organic carbon. Soil Science Society of America Journal, 1978, 42(5): 804- 809.
[3] Curl E A, Truelove B. The Rhizosphere. Berlin-Heidelberg: Springer-Verlag, 1986.
[4] Jones D L. Organic acids in the rhizosphere-a critical review. Plant and Soil, 1998, 205(1): 25- 44.
[5] Weller D M. Biological control of soilborne plant pathogens in the rhizosphere with bacteria. Annual Review of Phytopathology, 1988, 26(1): 379- 407.
[6] Van Loon L C, Bakker P A H M, Pieterse C M J. Systemic resistance induced by rhizosphere bacteria. Annual Review of Phytopathology, 1998, 36(1): 453- 483.
[7] 弋良朋, 馬健, 李彥. 荒漠鹽生植物根際土壤鹽分和養分特征. 生態學報, 2007, 27(9):3564- 3571.
[8] 董利蘋, 曹靖, 李先婷, 代立蘭, 蘇怡兵. 蘇怡兵不同耐鹽植物根際土壤鹽分的動態變化. 生態學報, 2011, 31(10): 2813- 2821.
[9] 詹媛媛, 薛梓瑜, 任偉, 周志宇. 干旱荒漠區不同灌木根際與非根際土壤氮素的含量特征. 生態學報, 2009, 29(1): 59- 66.
[10] 李國輝, 陳慶芳, 黃懿梅, 安韶山, 鄭粉莉, 陳利頂. 黃土高原典型植物根際對土壤微生物生物量碳、氮、磷和基礎呼吸的影響. 生態學報, 2010, 30(4): 976- 983.
[11] 謝添, 李戀卿, 潘根興, 杜有新. 不同退化程度喀斯特生態系統根際土壤的養分分布特 征. 生態環境學報, 2011, 20(2): 276- 280.
[12] 陳立新, 段文標, 喬璐. 落葉松人工林根際與非根際土壤養分特征及酸度研究. 水土保持學報, 2011, 25(3): 131- 135.
[13] 李從娟, 馬健, 李彥. 五種沙生植物根際土壤的鹽分狀況. 生態學報, 2009, 29(9):4549- 4655.
[14] 李從娟, 李彥, 馬健, 范連連, 王吉利. 干旱區植物根際土壤養分狀況的對比研究. 干旱區地理, 2011, 34(2): 222- 228.
[15] 王衛霞, 羅明, 潘存德. 塔里木河下游幾種荒漠植物根際土壤微生物及其活性. 中國 沙漠, 2010, 30(3): 571- 576.
[16] Riley D, Barber S A. Bicarbonate accumulation and pH changes at the soybean (Glycinemax(L.) Merr.) root-soil interface. Soil Science Society of America Journal, 1969, 33(6): 905- 908.
[17] Riley D, Barber S A. Salt accumulation at the soybean (Glycinemax. (L.) Merr.) root-soil interface. Soil Science Society of America Journal, 1970, 34(1): 154- 155.
[18] Barber S A. Soil Nutrient Bioavailability: a Mechanistic Approach. New York: John Wiley & Sons, 1995.
[19] Cleveland C C, Townsend A R, Taylor P, Alvarez-Clare S, Bustamante M M C, Chuyong G, Dobrowski S Z, Grierson P, Harms K E, Houlton B Z, Marklein A, Parton W, Porder S, Reed S C, Sierra C A, Silver W L, Tanner E V J, Wieder, W R. Relationships among net primary productivity, nutrients and climate in tropical rain forest: a pan-tropical analysis. Ecology Letters, 2011, 14(9): 939- 947.
[20] Brookes P C, Landman A, Pruden G, Jenkinson D S. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biology and Biochemistry, 1985, 17(6): 837-842.
[21] Rovira A D. Plant root excretions in relation to the rhizosphere effect. Plant and Soil, 1956, 7(2): 178- 194.
[22] Ruan J, Ma L F, Shi Y Z, Zhang F S. Effects of litter incorporation and nitrogen fertilization on the contents of extractable aluminium in the rhizosphere soil of tea plant (Camalliasinensis(L.) O. Kuntze). Plant and Soil, 2004, 263(1): 283- 296.
[23] Dieffenbach A, Matzner E. Insitusoil solution chemistry in the rhizosphere of mature Norway spruce (Piceaabies[L.]Karst.) trees. Plant and Soil, 2000, 222(1- 2): 149- 161.
[24] Shi W M, Wang X C, Yan W D. Distribution patterns of available P and K in rape rhizosphere in relation to genotypic difference. Plant and Soil, 2004, 261(1- 2): 11- 16.
[25] Rouatt J W, Katznelson H, Payne T M B. Statistical evaluation of the rhizosphere effect. Soil Science Society of America Journal, 1960, 24(4): 271- 273.
[26] Valiela I, Foreman K, LaMontagne M, Hersh D, Costa J, Peckol P, DeMeo-Andreson B, D’Avanzo C, Babione M, Sham C H, Brawley J, Lajtha K. Couplings of watersheds and coastal waters: sources and consequences of nutrient enrichment in Waquoit Bay, Massachusetts. Estuaries and Coasts, 1992, 15(4): 443- 457.
[27] Li J, Zhao C, Zhu H, Li Y, Wang F. Effect of plant species on shrub fertile island at an oasis-desert ecotone in the South Junggar Basin, China. Journal of Arid Environments, 2007, 71(4): 350- 361.
[28] Smith S D, Huxman T E, Zitzer S F, Charlet T N, Housman D C, Coleman J S, Fenstermaker L K, Seemann J R, Nowak R S. Elevated CO2increases productivity and invasive species success in an arid ecosystem. Nature, 2000, 408(6808): 79- 82.
[29] Wezel A, Rajot J L, Herbrig C. Influence of shrubs on soil characteristics and their function in Sahelian agro-ecosystems in semi-arid Niger. Journal of Arid Environments, 2000, 44(4): 383- 398.
[30] Bell D L, Sultan S E. Dynamic phenotypic plasticity for root growth in Polygonum: a comparative study. American Journal of Botany, 1999, 86(6): 807- 819.
[31] Dinkelaker B, R?mheld V, Marschner H. Citric acid excretion and precipitation of calcium citrate in the rhizosphere of white lupin (LupinusalbusL.). Plant, Cell & Environment, 1989, 12(3): 285- 292.
[32] Marschner P, Neumann G, Kania A, Weiskopf L, Lieberei R. Spatial and temporal dynamics of the microbial community structure in the rhizosphere of cluster roots of white lupin (LupinusalbusL.). Plant and Soil, 2002, 246(2): 167- 174.
[33] Bowen G D, Rovira A D. The rhizosphere: the hidden half of the hidden half// Waisel Y, Eshel A, Kafkafi U. Plant Roots: The Hidden Half. New York, USA: Marcel Dekker, 1991: 641- 669.
[34] Itai C, Vaadia Y. Kinetin-like activity in root exudate of water-stressed sunflower plants. Physiologia Plantarum, 1965, 18(4): 941- 944.
[35] Neumann G, R?mheld V. Root excretion of carboxylic acids and protons in phosphorus-deficient plants. Plant and Soil, 1999, 211(1): 121- 130.
[36] Dong J, Mao W H, Zhang G P, Wu F B, Cai Y. Root excretion and plant tolerance to cadmium toxicity-a review. Plant Soil and Environment, 2007, 53(5): 193- 200.
[37] Kraffczyk I, Trolldenier G, Beringer H. Soluble root exudates of maize: influence of potassium supply and rhizosphere microorganisms. Soil Biology and Biochemistry, 1984, 16(4): 315- 322.
[38] Shaw L J, Morris P, Hooker J E, Joner E. Perception and modification of plant flavonoid signals by rhizosphere microorganisms. Environmental Microbiology, 2006, 8(11): 1867- 1880.
[39] Chaudhry Q, Blom-Zandstra M, Gupta S K, Joner E J. Utilising the synergy between plants and rhizosphere microorganisms to enhance breakdown of organic pollutants in the environment (15 pp). Environmental Science and Pollution Research, 2005, 12(1): 34- 48.
Distribution of soil nutrient and microbial biomass in rhizosphereversusnon-rhizosphere area of different plant species in desertified steppe
YANG Yang, LIU Bingru*
KeyLaboratoryofRestorationandReconstructionofDegradedEcosysteminNorthwestChinaofMinistryofEducation,NingxiaUniversity,Yinchuan750021,China
Six plant species,Stipabungeana,AgropyronmongolicumKeng,GlycyrrhizauralensisFisch.,CynanchumkomaroviiAl.,Artemisiaordosica, andSophoraalopecuroidesdominate the desertified steppe ecosystems in Ningxia, northern China. However, the distribution of soil nutrients and microbial biomass in the rhizosphere and non-rhizosphere areas of these six plant species in this region has been largely unknown. This study investigated soil nutrients, microbial biomass, and soil physicochemical properties in the rhizosphere and non-rhizosphere areas of these six desert plant species, together with soil physicochemical properties. The results showed that rhizosphere enrichment followed the order shrub (A.ordosica) > Leguminosae (S.alopecuroides,G.uralensisFisch.) > Gramineae (S.bungeana,A.mongolicum) >Compositae(C.komaroviiAl.). In the rhizosphere area, a significant enrichment effect (P< 0.05) was found on the soil nutrients and microbial biomass, but not on the total phosphorus (TP) (P> 0.05). The rhizosphere enrichment effect on soil organic carbon (SOC) was more pronounced than the other indices (i.e., soil nutrients and physicochemical properties), and soil nutrient availability was more sensitive to the changes in the rhizosphere among plant species. In the rhizosphere and non-rhizosphere soil of different desert plant species, a significant linear correlation was observed (P< 0.01) between SOC and total nitrogen (TN) and a significant linear correlation between TN and alkaline hydrolysis nitrogen (P< 0.01), but no significant correlation was observed between TP and available phosphorus (AP) (P> 0.05). The results suggested that the rhizosphere enrichment effect occurred for many desert plant species. Shrubs and leguminous plants had a significantly higher rhizosphere effect than the other herbaceous species. Rhizosphere enrichment effect was found to increase soil nutrients by reducing soil pH, and this was beneficial for the efficient utilization of soil nutrients in the susceptible desertified steppe ecosystems.
desertified steppe; plant rhizosphere; soil nutrients; soil microbial biomass; spatial distribution
寧夏自然科學基金項目(NZ14015)
2014- 03- 13;
日期:2015- 04- 20
10.5846/stxb201403130440
*通訊作者Corresponding author.E-mail: bingru.liu@163.com
楊陽, 劉秉儒.荒漠草原不同植物根際與非根際土壤養分及微生物量分布特征.生態學報,2015,35(22):7562- 7570.
Yang Y, Liu B R.Distribution of soil nutrient and microbial biomass in rhizosphereversusnon-rhizosphere area of different plant species in desertified steppe.Acta Ecologica Sinica,2015,35(22):7562- 7570.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
