404 Not Found
404 Not Found
減蛋下降綜合征病毒重組Knob-S基因在桿狀病毒系統中的表達及其免疫原性鑒定
鄭 金, 陳 滔,, 陸吉虎, 高 峰,, 候繼波, 黎滿香, 唐應華
(1.湖南農業大學動物醫學院,湖南 長沙 410128;2.江蘇省農業科學院/國家獸用生物制品工程技術研究中心,江蘇 南京210014)
減蛋下降綜合征(EDS)是由減蛋下降綜合征病毒(EDS virus,EDSV)引起的以產蛋率下降為特征的傳染病。本病的主要傳播方式是垂直傳播,也可通過呼吸道水平傳播。所有年齡的雞均可感染,但幼雞感染后不表現任何臨床癥狀,母雞只是在產蛋高峰期表現明顯[1]。EDSV由11條大小不等的蛋白質組成,纖突蛋白質是EDSV的主要衣殼蛋白質,纖突蛋白質包括3個結構域,即Tail、Shaft和Knob區,Knob區不但可識別宿主細胞受體而且能增強病毒的吸附能力[2],Knob-S蛋白質存在良好的免疫原性[3]。
本試驗選用昆蟲桿狀病毒表達系統,對EDSV的Knob-S蛋白質進行真核表達、純化,經Western blotting和間接免疫熒光分析,確定表達的蛋白質具有免疫原性,為研究Knob-S蛋白質的生物學活性及相關疫苗奠定基礎。
1 材料與方法
1.1 質粒、菌株、細胞、試驗動物
大腸桿菌DH5α、pFastBac-1、轉座用大腸桿菌宿主菌 DH10Bac、pMD18-T-Knob-S、草地貪夜蛾卵巢細胞(Spodoptera frugiperda)均由江蘇省農業科學院保存。6周齡非免海蘭褐雛雞,購自北京華都峪口禽業集團。
1.2 主要試劑
EDSV-76由江蘇省農業科學院國家獸用生物制品工程技術研究中心保存。BamHⅠ、HindⅢ工具酶、Ex Taq DNA 聚合酶、IPTG、DNA marker DL2000、DL15000、蛋白質 Marker、X-gal、T4 DNA 連接酶、DNA膠回收試劑盒和質粒小提試劑盒均購自寶生物工程有限公司,優級胎牛血清、Grace’s培養基、Sf-900ⅡSFM培養液、LipofectamineTM2000均購自Invitrogen公司,FITC標記的羊抗雞的抗體為Promega公司產品,DMEM與RPMI1640培養基為GIBCO公司產品,細胞培養板為Costar公司產品,其余試劑均為進口或國產分析純。
1.3 引物設計與合成
以EDSV-76疫苗株為模版,根據已發表的纖突蛋白質Knob-S氨基酸序列,利用Primer Premier 5.0生物軟件設計1對引物,于上、下游引物分別引入BamHⅠ、HindⅢ酶切位點,F:5'-GCGGGATCCCA TGTTGACTTTGGCTTATGATTCCAC-3',R:5'-ATAAAGCTTCTACTGTGCTCCAACATATG-3',下劃線部分為酶切位點,擴增片段長為708 bp,引物由南京金斯瑞生物科技公司合成。
1.4 Knob-S基因的擴增、克隆
提取pMD18-T-Knob-S質粒,作為PCR擴增的模板,產物經10 g/L的瓊脂糖凝膠電泳鑒定和切膠回收后連接到pMD18-T中,轉化于DH5α感受態細胞,挑取白斑,提取質粒進行PCR和酶切鑒定。鑒定正確的質粒送上海美吉生物技術有限公司測序,測序正確的質粒命名為p-Knob-S。
1.5 重組桿狀病毒的構建
將重組質粒p-Knob-S和pFastBac-1質粒用BamHⅠ和HindⅢ雙酶切,回收目的片段連接獲得重組轉座載體pFast-Knob-S。將重組質粒pFast-Knob-S轉化入DH10Bac感受態細胞,獲得插入Knob-S基因的重組桿狀病毒穿梭載體rBac-Knob-S。3次藍白斑篩選純化后,按 Bac-to-Bac桿狀病毒表達系統說明書中改良的堿裂解法提取重組Bacmid的DNA,用M13上、下游通用引物進行PCR擴增,確定目的基因插入是否成功。將rBac-Knob-S在脂質體介導下轉染Sf9細胞,感染4~5 d時,收集出現明顯感染癥狀的細胞上清液,作為第1代重組病毒,傳代3次后篩選到含有Knob-S基因的重組桿狀病毒rAcV-Knob-S種子液。
1.6 重組桿狀病毒滴度的測定
取10支1.5 ml滅菌EP管,每管中加入含有2%FBS 的 Grace’s培養基 900 μl,在第一管中加入100 μl第3代病毒原液,倍比稀釋至第10管,最高稀釋度為1×10-10,棄去已鋪好的96孔細胞培養板內的培養液,用Grace’s培養基清洗1遍,每列細胞板依次加入100 μl所稀釋的病毒滴度,最后兩列為陰性對照,對照內加入 100 μl含有 2%FBS的Grace’s培養基,置于培養箱內,觀察細胞狀態并記錄陽性孔中細胞病變情況,直到陰性細胞脫落老去,結果按Karber法計算。
1.7 Western blotting和間接免疫熒光(IFA)檢測重組Knob-S基因在Sf9細胞中的表達
收集rBac-Knob-S感染后4~5 d的病變細胞,用PBS洗滌3次后,再用 PBS重懸,進行 Western blotting[4]。收集 rBac-Knob-S感染后完全病變的細胞,同時以未感染的Sf9昆蟲細胞和與空桿狀病毒感染的Sf9昆蟲細胞為對照,固定后用雞抗EDSV陽性血清和FITC標記的羊抗雞二抗檢測表達蛋白質,熒光顯微鏡下觀察結果。
1.8 抗原的制備
重組桿狀病毒調整到濃度為1 ml 1×106.0TCID50后,按每1 ml病毒接種100 ml的細胞(細胞密度約為1 ml 1×106個),共接種200 ml細胞,接毒后96~120 h收獲細胞,PBS洗3次,10 ml PBS重懸(濃縮20倍),超聲裂解,4℃以5 000 r/min離心10 min,棄掉沉淀。
1.9 疫苗的制備及動物免疫
將重組蛋白質經初步離心純化后與吐溫混合制備水相,將此水相與油相按 1∶3(體積比)混合并充分乳化,制成油乳劑疫苗并于4℃冰箱放置。將30只6周齡非免海蘭褐雛雞隨機分成3組,每組10只,桿狀病毒表達蛋白質各組每羽份免疫0.3 ml(含50 μg的重組病毒)。免疫后第2、3、4和5周分別采血檢測抗體效價。另設空Bacmid轉染Sf9細胞對照組和不免疫空白對照組。免疫后第 2、3、4和 5周分別采血,以EDS-76病毒作為檢測抗原,檢測血清血凝抑制抗體(HI)效價。
2 結果
2.1 重組穿梭載體的構建及鑒定
分別以 rBac-Knob-S和 rBac-pFast為模板,用M13通用引物進行PCR,擴增出的重組空載體大小為2 430 bp,重組載體大小為3 000 bp左右,與預期結果相符(圖1)。
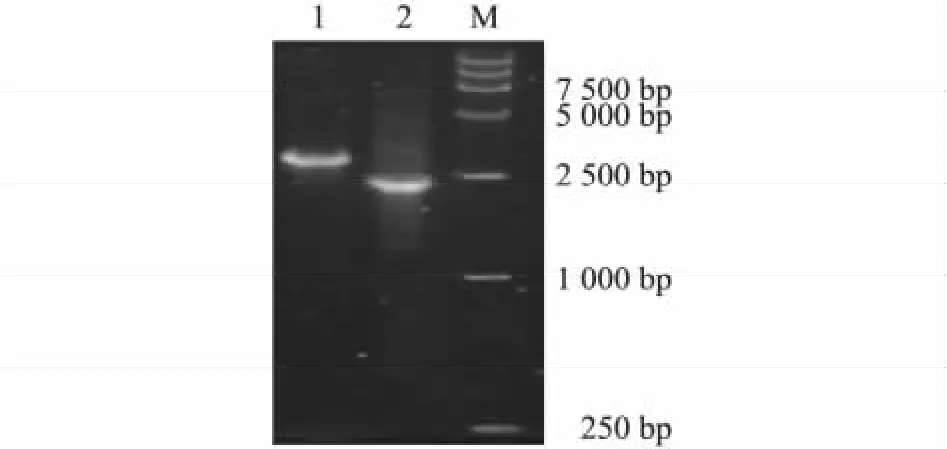
圖1 重組rBac-Knob-S PCR鑒定Fig.1 Ⅰdentification for the recombinant rBac-Knob-S with PCR
2.2 重組桿狀病毒滴度的測定
用Karber法計算病毒滴度,結果表明,前5個病毒稀釋度產生的細胞病變率為 100%,10-6、10-7、10-8、10-9、10-10稀釋的病毒產生的細胞病變率分別為 75.0%、37.5%、0、0 和0。
根據細胞的病變率計算重組桿狀病毒的病毒滴度,病毒滴度用組織培養半數感染量(TCID50)表示。
lgTCID50=L+d(S-0.5)
L:最低稀釋度對數,為-1;d:稀釋度對數之間的差值,為-1;S:陽性孔比率總和,為1+1+1+1+1+0.75+0.375+0=6.125。
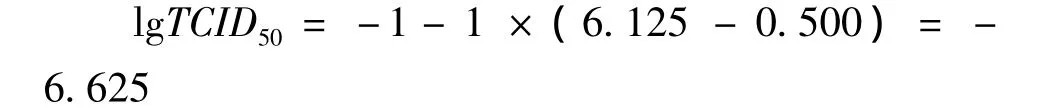
即0.1ml重組桿狀病毒的TCID50為10-6.625。
2.3 重組蛋白質Western blotting鑒定
經Western blotting檢測,重組桿狀病毒表達產物轉印反應后出現特異性蛋白質雜交帶(圖2),表明利用桿狀病毒在昆蟲細胞Sf9內表達的重組蛋白質可與雞陽性血清發生特異性免疫反應,從而證實其具有生物學活性。
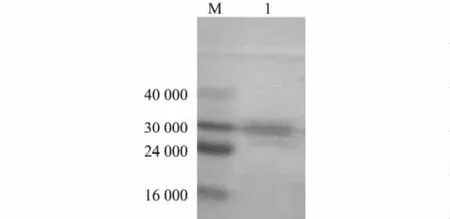
圖2 重組桿狀病毒表達蛋白質的Western blotting鑒定Fig.2 Western blotting analysis of recombinant rBac-Knob-S
2.4 間接免疫熒光檢測
經IFA檢測,rBac-Knob-S感染的Sf9細胞具有很強的特異性熒光(圖3A),而健康Sf9細胞(圖3B)無熒光,桿狀病毒感染的Sf9細胞(圖3C)無熒光,說明Knob-S基因得到表達,并且表達產物位于細胞內。
2.5 血清中HI抗體的檢測
重組Knob-S蛋白質免疫后第2、3、4和5周采血,分離血清,HI檢測的抗體效價見表1,結果顯示,重組Knob-S蛋白免疫后第2周即可產生較高抗體,

圖3 重組病毒的間接免疫熒光(ⅠFA)檢測Fig.3 Ⅰdentification of recombinant bacmid by indirect immunofluorescence assay
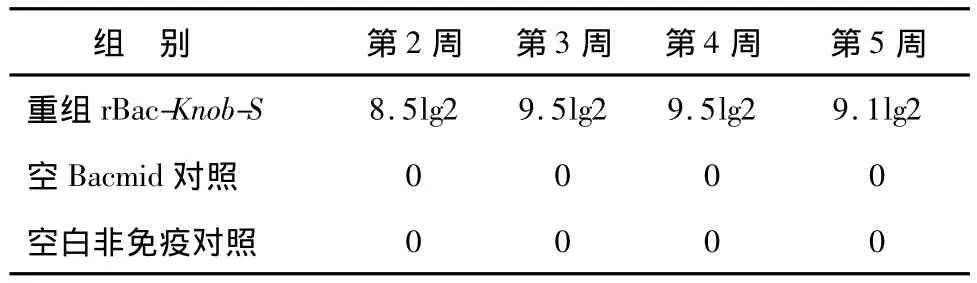
表1 抗EDSV HⅠ抗體效價Table 1 The hemagglutinin inhibition(HⅠ)antibody titers against EDSV
3 討論
目前減蛋下降綜合征病毒(EDSV)的疫苗是EDSV用鴨胚增殖,并滅活制備成油苗,用以預防EDS-76,現階段并無SPF鴨胚用于繁殖病毒,若滅活不完全,則有可能造成巨大的經濟損失,存在很大的安全隱患。利用昆蟲桿狀病毒表達系統(BEVS)體外表達具有生物活性蛋白質國內外均有文獻報道[5-10],該系統具有對外源蛋白質加工后修飾的功能,容易從無血清培養上清液中純化蛋白質,無內毒素污染等優點。該系統與大腸桿菌等原核表達的外源蛋白質相比能更好保持其免疫原性。
本試驗以AV-127株基因組為模板,利用PCR方法擴增纖突蛋白質基因的Knob-S區,將其克隆到真核表達載體pFastBac1中,經雙酶切證實重組質粒構建正確后構建穿梭載體rBac-Knob-S,經三輪篩選后得到陽性菌落,提取病毒基因組轉染Sf9細胞,在細胞中可見很強的亮綠色熒光,說明在Sf9細胞中高效表達了該蛋白質。病毒的滴度測定結果表明,重組Knob-S病毒感染Sf9細胞后,細胞出現病變死亡,說明融合表達的Knob-S蛋白質有細胞毒性。通過Western blotting檢測,確定了Knob-S蛋白質的生物學活性,與預期結果相符,表明表達的蛋白質具有與天然蛋白質相似的構象和相同抗原性。動物免疫試驗結果表明,真核表達重組蛋白質的抗體效價較高,說明該真核表達蛋白質具有較好的免疫原性。
[1] W卡尼爾B.禽病學[M].10版.北京:中國農業大學出版社,1999.
[2] TOMKO R P,JOHANSSON C B,TOTROV M,et al.Expression of the adenovirus receptor and its interaction with the fiber knob[J].Experimental Cell Research,2000,255(1):47-55.
[3] FINGERUT E,GUTTER B,GALLILI G,et al.A subunit vaccine against the adenovirus egg-drop syndrome using part of its fiber protein[J].Vaccine,2003,21(21):2761-2766.
[4] 陳 滔,唐應華,陸吉虎,等.產蛋下降綜合征病毒纖突蛋白Knob-S區原核表達及其活性分析[J].動物醫學進展,2012,33(6):41-44.
[5] KOST T A,CONDREAY J P,JARVIS D L.Baculovirus as versatile vectors for protein expression in insect and mammalian cells[J].Nature Biotechnology,2005,23(5):567-575.
[6] SHEN X,HU G,JIANG S,et al.Engineering and characterization of a baculovirus-expressed mouse/human chimeric antibody against transferrin receptor[J].Protein Engineering Design and Selection,2009,22(12):723-731.
[7] WANG Y,SUN Y,TIAN Z,et al.Protection of chickens against infectious bronchitis by a recombinant fowlpox virus co-expressing IBV-S1 and chicken IFNγ[J].Vaccine,2009,27(50):7046-7052.
[8] NAGATA T,ISHIKAWA S,SHIMOKAWA E,et al.High level expression and purification of bioactive bovine interleukin-18 using a baculovirus system[J].Veterinary Immunology and Immunopathology,2002,87(1):65-72.
[9] 唐應華,吳培培,宮玉珍,等.腎型傳染性支氣管炎病毒的鑒定和結構蛋白的表達[J].中國獸醫科學,2011,41(10):1034-1040.
[10]李東衛,郭瑩瑩,劉在斯,等.禽流感病毒 NS1蛋白在桿狀病毒表達系統中的表達[J].動物醫學進展,2012,33(6):27-31.
