高郵鴨群體 MSTN、MyoD1和 MyoG基因外顯子中SNP位點的分析
2016-01-15 06:01:33劉宏祥,徐文娟,宋衛濤等
江蘇農業學報 2015年3期
高郵鴨群體MSTN、MyoD1和MyoG基因外顯子中SNP位點的分析
劉宏祥,徐文娟,宋衛濤,朱文奇,胡艷,姬改革,李慧芳
(江蘇省家禽科學研究所,江蘇揚州225125)

摘要:MSTN、MyoD1和MyoG基因與禽類骨骼肌的生長發育密切相關。本試驗以高郵鴨為素材,對MSTN、MyoD1和MyoG 3個基因的外顯子單核苷酸多態性(SNP)位點進行了鑒定,并對這些位點對蛋白質的影響及群體遺傳信息進行了分析。結果顯示,MSTN、MyoD1和MyoG 3個基因的外顯子分別有9個、4個和7個SNP位點,其中具有中度多態的位點分別為8個、1個和5個,有義突變各1個,這些有義突變均改變了相應蛋白質的理化性質以及二級結構。
關鍵詞:MSTN;MyoD1;MyoG;外顯子;單核苷酸多態性;鴨
doi:10.3969/j.issn.1000-4440.2015.03.023
收稿日期:2014-12-30
基金項目:國家自然科學基金項目(31172194);江蘇省科技支撐計劃項目(BE2011329、BE2012460);現代農業品種創新項目[CX(11)1030]
作者簡介:劉宏祥(1985-),男,江蘇揚州人,碩士,主要從事家禽遺傳育種與資源保護研究。(Tel)13665203762;(E-mail)liuhongxiangzai@163.com
中圖分類號:S834`+.89
文獻標識碼:A
文章編號:1000-4440(2015)03-0604-09
Abstract:There are strong relations between MSTN, MyoD1, MyoG genes and development of skeletal muscle. The single nucleotide polymorphisms (SNPs) in the exons of MSTN, MyoD1 and MyoG genes of Gaoyou duck were detected, and their influences on proteins and population genetics information were studied. There were nine, four SNPs and seven SNPs in the exons of MSTN, MyoD1 and MyoG genes, respectively, among which, eight, one and five SNPs were moderate polymorphic. One sense mutation detected in each gene changed the protein physico-chemical characteristics and second structure.
Analysis of single nucleotide polymorphisms (SNPs) in exons ofMSTN, MyoD1 andMyoGgenes of Gaoyou duck
LIU Hong-xiang,XU Wen-juan,SONG Wei-tao,ZHU Wen-qi,HU Yan,JI Gai-ge,LI Hui-fang
(JiangsuInstituteofPoultryScience,Yangzhou225125,China)
Key words:MSTN;MyoD1;MyoG;exon;single nucleotide polymorphism (SNP);duck
動物機體的生長是一個復雜的生理過程,對肌肉生長具有調節作用的因子很多,其中重要的有MRFs(Muscle regulation factors)家族的MyoD1基因、MyoG基因,它們對肌肉生長具有正向調節功能;而TGFβ家族中的MSTN基因對肌肉生長具有負調控作用,其功能的缺失在哺乳動物中能夠引起肌肉的顯著增大[2-4]。
具有編碼蛋白質功能的基因在DNA水平上的堿基序列變化將傳遞給蛋白質,而蛋白質序列的改變將影響蛋白質二級結構以及高級結構,甚至改變蛋白質的功能。基因組水平上的單個核苷酸的變異(SNP)在染色體上數量多、分布廣,而且能夠穩定遺傳。因此在動物遺傳育種中,對功能基因,尤其直接編碼蛋白質的外顯子部分的SNP研究具有重要意義。
已有多個研究報道了哺乳動物(比如牛[3, 5]、小鼠、綿羊[7-8]、狗)MSTN基因外顯子的錯義突變導致了機體肌肉的顯著增長。張跟喜等[10]在2011年對邊雞的MSTN基因外顯子1進行了SNP篩查,發現一個對邊雞生長有顯著效應的SNP位點。
高郵鴨屬于肉蛋兼用型地方品種,經過多年選育,目前高郵鴨群體性能已基本穩定。本試驗運用單重PCR方法對高郵鴨群體的MSTN、MyoD1和MyoG3個基因的外顯子分別進行了SNP篩查,并分析篩查到有義突變導致氨基酸變化以及對蛋白質二級結構的影響,為高郵鴨群體后期的分群以及開發利用提供理論依據。
1材料與方法
1.1 試驗動物
高郵鴨試驗群體來源于江蘇省高郵鴨集團,性成熟時從群體中隨機選擇30個體質量差異較小的個體進行試驗。
1.2 主要儀器和試劑
主要儀器有:PCR儀(型號:Gene Amp PCR,廠家:Norwalk,CT.06859 USA)、電泳儀(型號:JY600+,廠家:北京君意東方電泳設備有限公司)、全自動紫外與可見分析裝置(型號:FR-200A,廠家:上海復日科技有限公司)、生物電泳圖像分析系統(廠家:上海復日科技有限公司)。
主要試劑有:PCR引物(由上海瀚宇生物工程有限公司合成,PAGE級純化)、PCR反應試劑(上海有漁生物工程有限公司提供)。
1.3 DNA的提取
對30個高郵鴨個體翅靜脈采血,用酚-氯仿法抽提DNA并溶于超純水中。將DNA原液 1∶100稀釋并進行分光光度計檢測,OD值為 1.85~1.94。
1.4 引物設計
分別在MSTN基因3個外顯子、MyoD1基因4個外顯子、MyoG基因5個外顯子區域附近設計合適的引物,其中MSTN、MyoD1和MyoG基因分別設計了4、4和3對引物。具體信息見表 1。
表1針對MSTN、MyoD1和MyoG基因外顯子區域設計的引物
Table 1The primers for the exons ofMSTN, MyoD1 andMyoGgenes
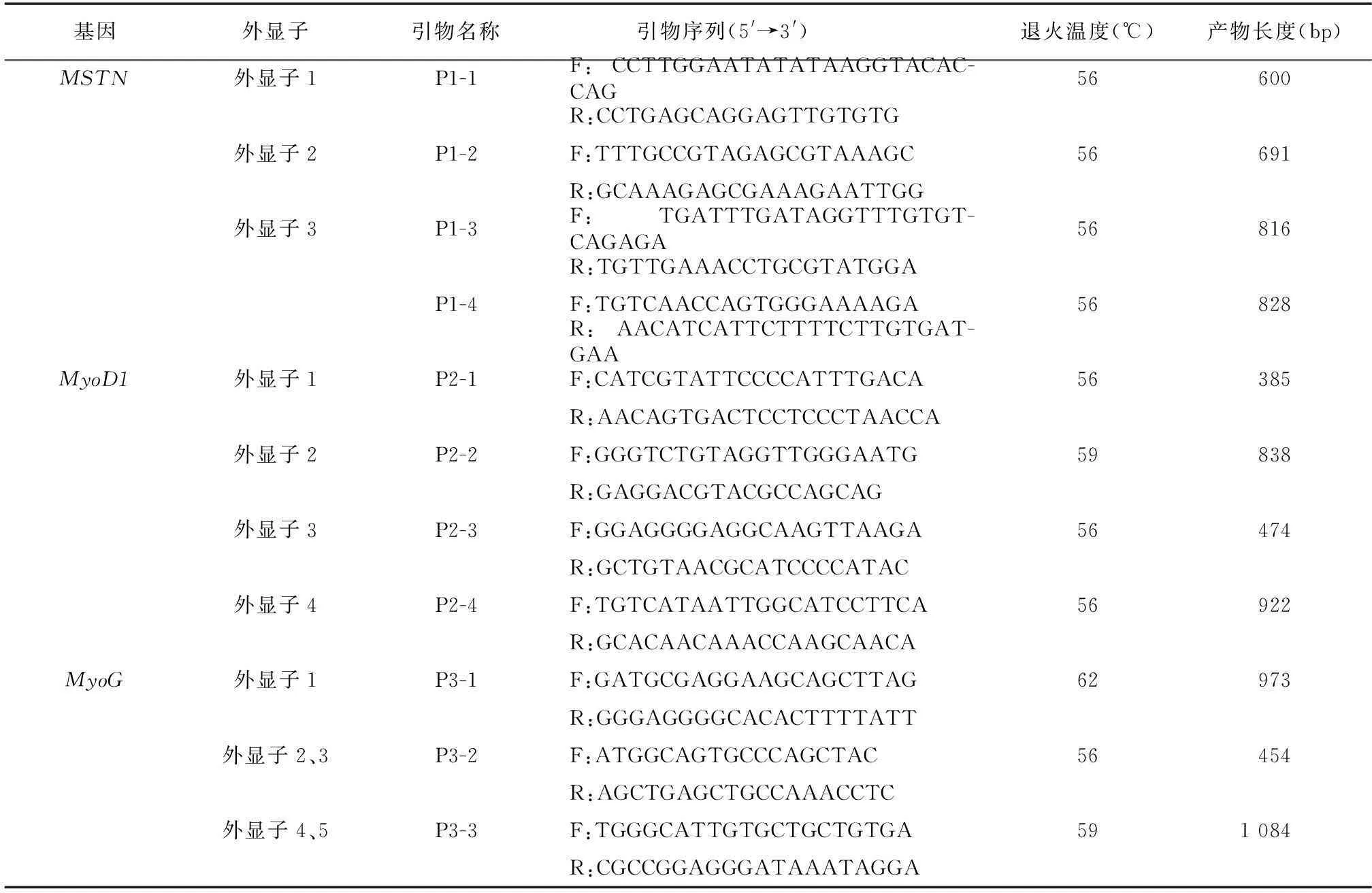
基因外顯子引物名稱 引物序列(5'→3')退火溫度(℃)產物長度(bp)MSTN外顯子1P1-1F:CCTTGGAATATATAAGGTACAC-CAG56600R:CCTGAGCAGGAGTTGTGTG外顯子2P1-2F:TTTGCCGTAGAGCGTAAAGC56691R:GCAAAGAGCGAAAGAATTGG外顯子3P1-3F:TGATTTGATAGGTTTGTGT-CAGAGA56816R:TGTTGAAACCTGCGTATGGAP1-4F:TGTCAACCAGTGGGAAAAGA56828R:AACATCATTCTTTTCTTGTGAT-GAAMyoD1外顯子1P2-1F:CATCGTATTCCCCATTTGACA56385R:AACAGTGACTCCTCCCTAACCA外顯子2P2-2F:GGGTCTGTAGGTTGGGAATG59838R:GAGGACGTACGCCAGCAG外顯子3P2-3F:GGAGGGGAGGCAAGTTAAGA56474R:GCTGTAACGCATCCCCATAC外顯子4P2-4F:TGTCATAATTGGCATCCTTCA56922R:GCACAACAAACCAAGCAACAMyoG外顯子1P3-1F:GATGCGAGGAAGCAGCTTAG62973R:GGGAGGGGCACACTTTTATT外顯子2、3P3-2F:ATGGCAGTGCCCAGCTAC56454R:AGCTGAGCTGCCAAACCTC外顯子4、5P3-3F:TGGGCATTGTGCTGCTGTGA591084R:CGCCGGAGGGATAAATAGGA
1.5 PCR產物擴增與測序
PCR反應體系為20 μl,其中包括:1.0 μl模板(50 ng)、2.0 μl Buffer緩沖液(1×)、0.6 μl鎂離子(2 mmol/L)、2.0 μl dNTP(2 mmol/L)、0.2 μlTaq酶(1 U)、2.0 μl引物混合液(0.5 pmol/L),余下用H2O補足。PCR反應條件為:95 ℃預變性2 min;94 ℃ 30 s,按照表 1中的退火溫度退火1 min 30 s,72 ℃延伸30 s,共40個循環;最后72 ℃后延伸10 min。PCR產物用3%的瓊脂糖凝膠電泳進行檢測擴增效果。最后對產物進行純化,并送由上海翼和應用生物技術有限公司測序。
1.6 統計分析
1.6.3Hardy-Weinberg平衡檢驗假設某群體的某一位點等位基因有A、B兩種,其等位基因頻率分別為p、q。該群體基因型有AA、AB、BB 3種,基因型頻率分別記為D、H和R。根據觀測到的基因型頻率,可以計算出等位基因頻率:
p=D+H/2,q=R+H/2
根據等位基因頻率,可以計算出理論的基因型頻率:
D′=p2,H′=2pq,R′=q2
根據樣本含量、觀測基因型頻率和理論基因型頻率,可以分別計算出觀測基因型頻數和理論基因型頻數。通過卡方檢驗:
χ2=(D-D′)2/D′+(H-H′)2/H′+(R-R′)2/R′
計算出卡方值。如果卡方值對應的概率大于0.05,那么可以認為該等位基因處于Hardy-Weinberg平衡狀態。
1.6.4蛋白質理化性質分析利用ExPASy網站中的ProtParam服務器(http://web.expasy.org/cgi-bin/protparam/)在線計算MSTN、MyoD1和MyoG蛋白質的理論等電點、分子量、正負電位氨基酸數目、不穩定系數、蛋白質親水性及脂肪族氨基酸指數等理化性質。
1.6.5蛋白質二級結構的預測蛋白質二級結構是一級結構和三級結構之間的紐帶,也是通過一級結構預測三級結構的關鍵步驟[11]。對MSTN、MyoD1和MyoG基因的外顯子突變情況進行了篩查,發現3個基因各有1個有義突變導致了氨基酸的變化。SSPro 4.0(http://scratch.proteomics.ics.uci.edu)可以使用神經網絡與同源分析混合進行蛋白質二級結構預測[12],預測的結果較為準確可靠,因此本試驗使用該服務器作為蛋白質二級結構預測的工具。
2結果與分析
2.1 MSTN、MyoD1和 MyoG 3個基因外顯子突變位點分析
擴增產物經測序、對比,MSTN外顯子中發現9個SNP位點,編號為SNP1-1~SNP1-9,具體信息見表2。SNP1-1~SNP1-5位于CDS區,其中SNP1-1導致了甘氨酸(G)到絲氨酸(S)的變化,其余為沉默突變。SNP1-6~SNP1-9位于非CDS區。
MyoD1外顯子中共發現4個SNP位點,編號為SNP2-1~SNP2-4。這4個SNP位點均處于CDS區,其中SNP2-1導致了蛋氨酸(M)向亮氨酸(L)的轉變,其他3個SNP位點均為沉默突變。
MyoG外顯子中共發現了7個SNP位點,編號為SNP3-1~SNP3-7。這7個SNP位點均處于CDS區,其中SNP3-7為有義突變,導致谷氨酸(E)到天冬氨酸(D)的變化,其余均為沉默突變。
2.2 MSTN、MyoD1和 MyoG 3個基因外顯子的遺傳多樣性
對30個高郵鴨個體的MSTN、MyoD1和MyoG基因的基因型頻率以及基因頻率分別進行了統計,統計結果分別見表3、表4和表5。
根據各個SNP位點的基因型分布情況,對各個位點進行了Hardy-Weinberg平衡檢驗,結果見表6。MSTN基因外顯子所有突變位點均處于Hardy-Weinberg平衡狀態;MyoD1基因突變位點SNP2-2、SNP2-3和SNP2-4均不處于Hardy-Weinberg平衡狀態;MyoG基因突變位點SNP3-2、SNP3-3和SNP3-5均不處于Hardy-Weinberg平衡狀態。
表2MSTN、MyoD1和MyoG基因突變位點
Table 2SNPs ofMSTN, MyoD1 andMyoGgenes

基因SNP位點外顯子雜合子類型氨基酸變化編碼子位置氨基酸位置優勢堿基MSTNSNP1-1外顯子1GAGGC/AGC(G/S)126AMSTNSNP1-2外顯子1GAACG/ACA(T)343AMSTNSNP1-3外顯子1CTGAC/GAT(D)3108CMSTNSNP1-4外顯子3AGGTA/GTG(V)3327GMSTNSNP1-5外顯子3CAGGC/GGA(G)3334AMSTNSNP1-6外顯子3TC 非CDS區CTMSTNSNP1-7外顯子3GT 非CDS區GMSTNSNP1-8外顯子3AC 非CDS區AMSTNSNP1-9外顯子3AC 非CDS區CMyoD1SNP2-1外顯子1ACATG/CTG(M/L)14AMyoD1SNP2-2外顯子4GAGAG/GAA(E)3190G*MyoD1SNP2-3外顯子4TCACT/ACC(T)3207T*MyoD1SNP2-4外顯子4TCCCT/CCC(P)3247T*MyoGSNP3-1外顯子1CTCTG/TTG(L)143CMyoGSNP3-2外顯子1GCGTG/GTC(V)379G*MyoGSNP3-3外顯子1CGCGC/CGG(R)3132C*MyoGSNP3-4外顯子3CTCAC/CAT(H)3157CTMyoGSNP3-5外顯子4GAACG/ACA(T)3187G*MyoGSNP3-6外顯子5GATCG/TCA(S)3203GMyoGSNP3-7外顯子5GCGAG/GAC(E/D)3222C
氨基酸變化欄中括號前的字母表示三聯密碼子的變化情況,括號中的字母表示氨基酸的變化情況;編碼子位置欄中數字表示三聯密碼子中發生突變的堿基在密碼子中的位置,非CDS區表示該突變不在CDS區中;優勢堿基表示在突變位點的2個等位堿基在群體中出現頻率較大的堿基;*表示該位點的2個堿基在群體中的分布不符合Hardy-Weinberg平衡。
表3MSTN基因SNP位點的群體信息
Table 3The population information about SNPs ofMSTNgene
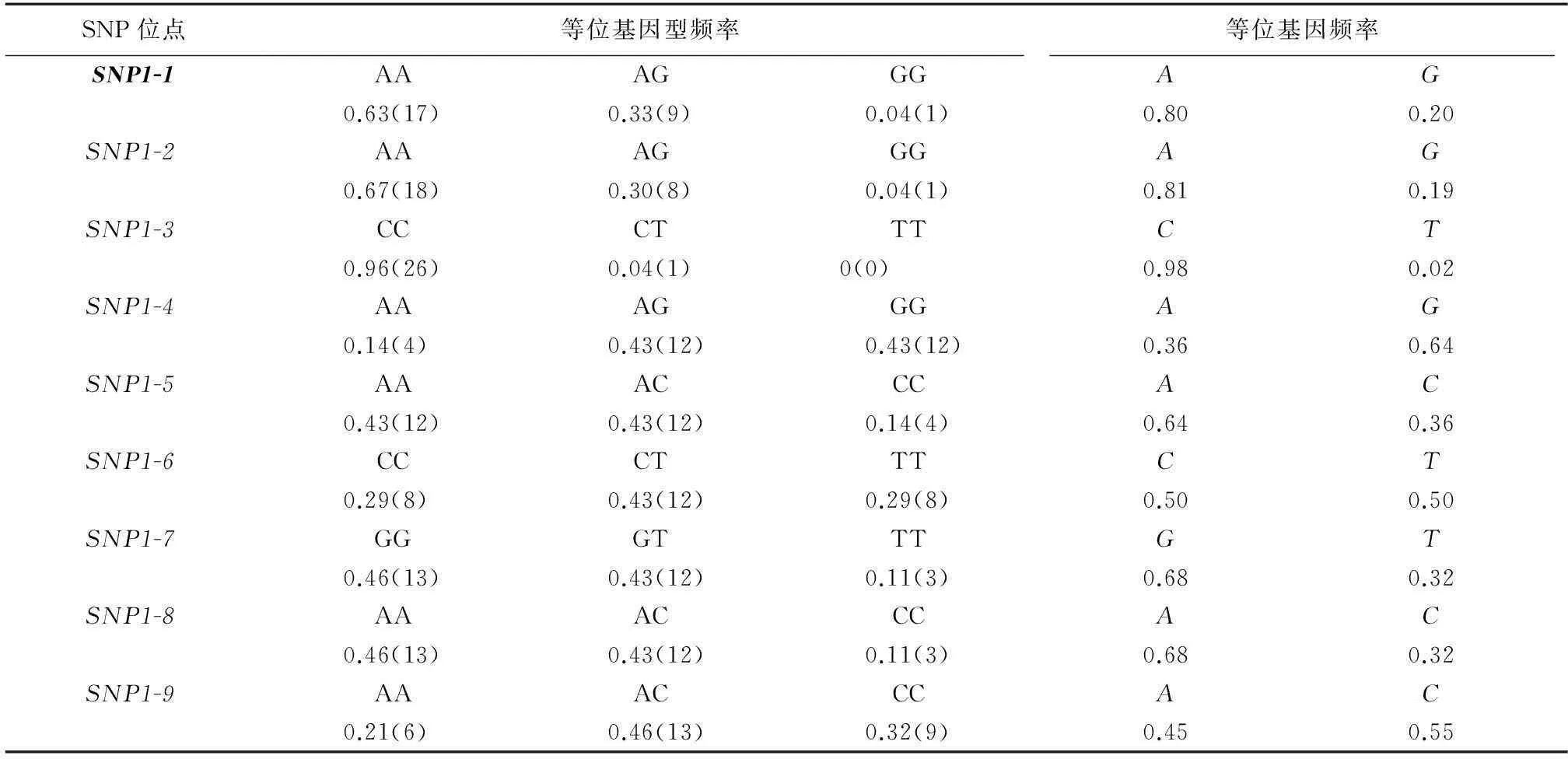
SNP位點等位基因型頻率等位基因頻率SNP1-1AAAGGGAG0.63(17)0.33(9)0.04(1)0.800.20SNP1-2AAAGGGAG0.67(18)0.30(8)0.04(1)0.810.19SNP1-3CCCTTTCT0.96(26)0.04(1)0(0)0.980.02SNP1-4AAAGGGAG0.14(4)0.43(12)0.43(12)0.360.64SNP1-5AAACCCAC0.43(12)0.43(12)0.14(4)0.640.36SNP1-6CCCTTTCT0.29(8)0.43(12)0.29(8)0.500.50SNP1-7GGGTTTGT0.46(13)0.43(12)0.11(3)0.680.32SNP1-8AAACCCAC0.46(13)0.43(12)0.11(3)0.680.32SNP1-9AAACCCAC0.21(6)0.46(13)0.32(9)0.450.55
括號內數字表示該等位基因型在檢測群體中的觀測個數。粗體表示有義突變。
表4MyoD1基因SNP位點的群體信息
Table 4The population information about SNPs of MyoD1 gene
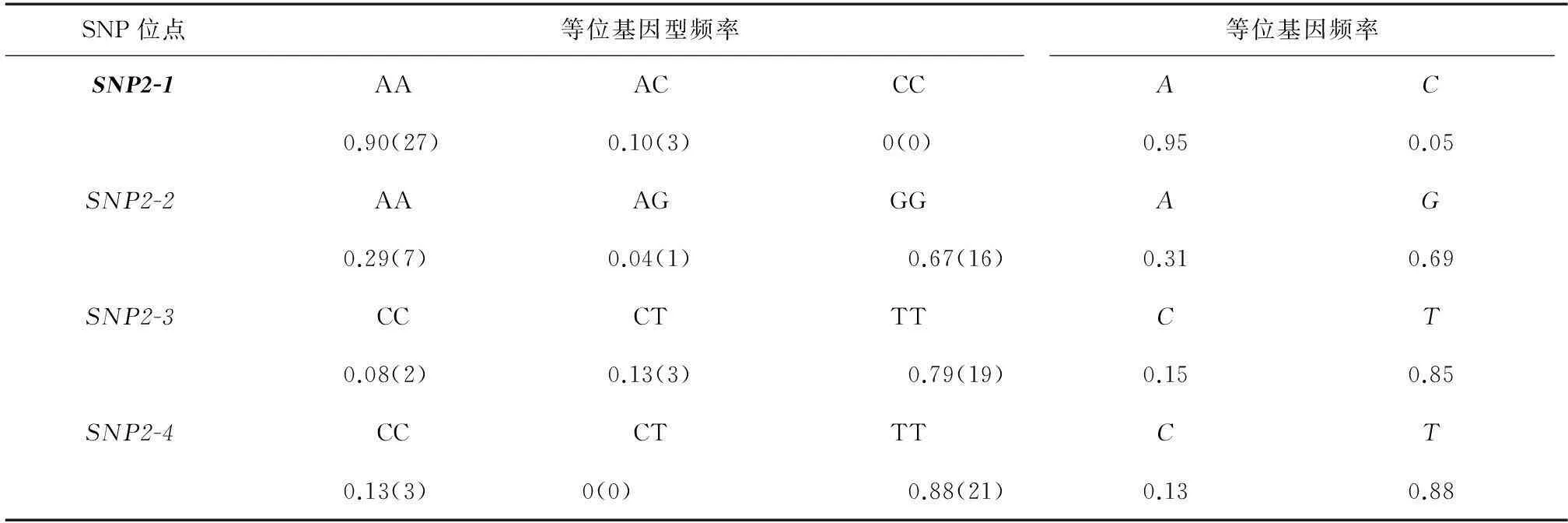
SNP位點等位基因型頻率等位基因頻率SNP2-1AAACCCAC0.90(27)0.10(3)0(0)0.950.05SNP2-2AAAGGGAG0.29(7)0.04(1)0.67(16)0.310.69SNP2-3CCCTTTCT0.08(2)0.13(3)0.79(19)0.150.85SNP2-4CCCTTTCT0.13(3)0(0)0.88(21)0.130.88
括號內數字表示該基因型在檢測群體中的觀測個數。粗體表示有義突變。
表5MyoG基因SNP位點的群體信息
Table 5The population information about SNPs ofMyoGgene

SNP位點等位基因型頻率等位基因頻率SNP3-1CCCTTTCT0.86(25)0.14(4)0(0)0.930.07SNP3-2CCGCGGCG0.10(3)0.34(10)0.55(16)0.280.72SNP3-3CCGCGGCG0.45(13)0.34(10)0.21(6)0.620.38SNP3-4CCCTTTCT0.33(7)0.33(7)0.33(7)0.500.50SNP3-5GGAGAAGA0.87(13)0.13(2)0(0)0.930.07SNP3-6AAAGGGAG0.07(1)0.40(6)0.53(8)0.270.73SNP3-7CCGCGGCG0.40(6)0.27(4)0.33(5)0.530.47
括號內數字表示該等位基因型在檢測群體中的觀測個數。粗體表示有義突變。
根據等位基因頻率計算出群體各個基因突變位點的多態信息含量(PIC)以及有效等位基因數(表 6)。PIC值介于0.25和0.50之間為中度多態,可知MSTN基因外顯子除了SNP1-3外,其他位點均為中度多態;MyoD1基因第4外顯子的1個突變位點SNP2-2為中度多態,MyoG基因突變位點SNP3-2、SNP3-3、SNP3-4、SNP3-6和SNP3-7為中度多態;所有其他突變位點均為低度多態。對于有效等位基因數,MSTN基因SNP1-3位點,MyoD1基因SNP2-1位點、SNP2-3位點和SNP2-4位點,MyoG基因SNP3-1位點和SNP3-5位點有效等位基因數均不超過1.4,與理論等位基因數2.0有較大差異,即這些位點遺傳變異較小。有效等位基因數的分析結果與多態信息含量的結果相似。
表6MSTN、MyoD1和MyoG3個基因的遺傳多樣性
Table 6Genetic diversity ofMSTN, MyoD1 andMyoGgenes

基因SNP位點多態信息含量(PIC)有效等位基因數(Ne)哈代-溫伯平衡(Hardy-Weinberg)MSTNSNP1-10.27181.48020.99MSTNSNP1-20.25621.43221.00MSTNSNP1-30.03571.03771.00MSTNSNP1-40.35381.84910.94MSTNSNP1-50.35381.84910.94MSTNSNP1-60.37502.00000.75MSTNSNP1-70.34111.77381.00MSTNSNP1-80.34111.77381.00MSTNSNP1-90.37211.97730.95MyoD1SNP2-10.09051.10500.96MyoD1SNP2-20.33741.75340*MyoD1SNP2-30.21811.33180.05*MyoD1SNP2-40.19481.28000*MyoGSNP3-10.12021.14730.92MyoGSNP3-20.31971.66530*MyoGSNP3-30.36001.88990*MyoGSNP3-40.37502.00000.31MyoGSNP3-50.11671.14210*MyoGSNP3-60.31461.64231.00MyoGSNP3-70.37391.99120.13
粗體表示有義突變。
2.3 突變導致的蛋白質理化性質的改變
MSTN、MyoD1和MyoG野生型蛋白質及突變型蛋白質的理化性質分析結果見表7。MSTN突變型蛋白質與野生型蛋白質相比,除了分子量變化之外,其不穩定系數變大(由44.80變為45.83),親水性變強(疏水性指數由-0.405變為-0.406),其他如理論等電點、正負電位點數以及脂肪族系數均沒有變化。MyoD1突變型蛋白質與野生型蛋白質相比,分子量變小,親水性變弱(疏水性指數由-0.917變為-0.910),脂肪族系數變大。MyoG突變型蛋白質與野生型蛋白質相比,除了分子量變小之外,其理論等電點與不穩定系數均變小,疏水性沒有變化。
表7蛋白質理化性質
Table 7Protein physico-chemical properties

蛋白質分子量理論等電點正電位點數負電位點數不穩定系數總平均疏水性脂肪族系數MSTN-RAW42817.16.27464844.80-0.40580.83MSTN-SNP42847.16.27464845.83-0.40680.83MyoD1-RAW28133.16.94293074.85-0.91756.15MyoD1-SNP28115.16.94293074.85-0.91057.70MyoG-RAW25793.05.19273667.37-0.64075.68MyoG-SNP25779.05.18273665.93-0.64075.68
RAW表示野生型,SNP表示突變型。
2.4 突變導致蛋白質二級結構的變化
2.4.1MSTN蛋白質二級結構的變化MSTN基因外顯子1中篩查到的有義突變導致了第26位氨基酸由甘氨酸到絲氨酸的變化。通過軟件分析,MSTN蛋白質在5~8位置由折疊變成了螺旋,在17~18位置、19~22位置由卷曲變成了螺旋,在136、190位置由卷曲變成了折疊(圖1)。折疊、螺旋、卷曲的比例由野生型的26.13%、15.73%、58.14%分別變成突變型的25.87%、18.13%、56.00%。

RAW表示野生型;SNP表示突變型。黑色背景表示突變型與野生型蛋白質二級結構間有變化。H表示螺旋,E表示拉長的折疊股,C表示卷曲。括號中的數字表示每一行最后的氨基酸在蛋白質中的位置。 圖1 MSTN野生型與突變型蛋白質二級結構的比較 Fig.1 The alignment of wild-type and mutated MSTN protein secondary structures
2.4.2MyoD1蛋白質二級結構的變化MyoD1基因外顯子1中的有義突變導致了第4位氨基酸從蛋氨酸到亮氨酸的變化。通過軟件分析,MyoD1蛋白質在4~5位置由野生型的卷曲變成突變型的螺旋,在30位置由野生型的折疊變成突變型的卷曲(圖2)。折疊、螺旋、卷曲的比例由野生型的2.78%、26.98%、70.24%分別變成突變型的2.38%、27.78%、69.84%。
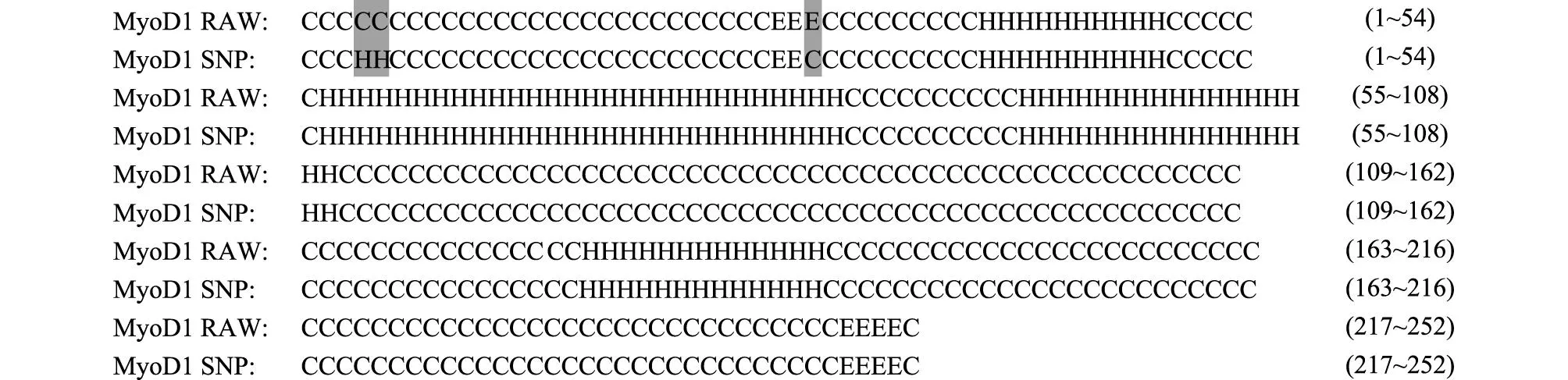
RAW表示野生型;SNP表示突變型。黑色背景表示突變型與野生型蛋白質二級結構間有變化。H表示螺旋,E表示拉長的折疊股,C表示卷曲。括號中的數字表示每一行最后的氨基酸在蛋白質中的位置。 圖2 MyoD1野生型與突變型蛋白質二級結構的比較 Fig.2 The alignment of wild-type and mutated MyoD1 protein secondary structures
2.4.3MyoG蛋白質二級結構的變化MyoG基因的第5外顯子的一處突變導致了第222位氨基酸從谷氨酸到天冬氨酸的改變。通過軟件分析,MyoG蛋白質在149、200、211位置由野生型的螺旋變成突變型的卷曲(圖3)。突變型的折疊所占比例0.88%沒有變化,螺旋、卷曲的比例分別從野生型的30.40%、68.72%變成突變型的29.07%、70.04%。

RAW表示野生型;SNP表示突變型。黑色背景表示突變型與野生型蛋白質二級結構間有變化。H表示螺旋,E表示拉長的折疊股,C表示卷曲。括號中的數字表示每一行最后的氨基酸在蛋白質中的位置。 圖3 MyoG野生型與突變型蛋白質二級結構的比較 Fig.3 The alignment of wild-type and mutated MyoG protein secondary structures
3討論
禽類肌肉組織肌纖維的數量在胚胎期即已確定[13],出殼后主要是通過已有肌纖維的衛星細胞相互融合而變得粗大[14]。肌纖維的形成與發育受到MRF家族基因的正向調控,MyoD1基因能夠將多種類型的細胞轉換為成肌細胞并促進肌細胞融合成肌管[15],MyoG基因的表達可終止成肌細胞的增殖,促進單核成肌細胞融合成多核成肌細胞[16-17]。作為肌肉的負調控因子,MSTN可以抑制成肌細胞的增殖和分化[18]。MSTN不僅可以下調MyoD1基因的表達[19-20],而且還是MyoD1的下游靶基因[21]。MSTN、MyoD1和MyoG基因三者對肌肉的調節是相互協調保持相對的平衡,因此開展MSTN、MyoD1和MyoG3個基因的SNP位點篩查對于生長發育有重要的科學意義。
本研究發現高郵鴨群體MSTN、MyoD1和MyoG基因外顯子中分別有9、4和7個SNP位點。其中MSTN基因的所有7個SNP位點均處于Hardy-Weinberg平衡狀態。鴨MyoD1基因和MyoG基因的多態性報道較少。Xu等在北京鴨群體中發現MSTN基因CDS區129位置有1個T/C突變[22],與本試驗發現的SNP1-2位點(CDS區第129位置)A/G突變一致。該突變位點2個等位基因在北京鴨上頻率接近,而在高郵鴨中A等位基因為優勢基因,這可能與鴨不同遺傳背景有關。除了本試驗篩查到的MSTN基因外顯子中的7個SNP位點,其他研究者也發現了其他多個SNP位點[23-25],說明MSTN基因的多態性位點在鴨上非常豐富。
寇潔等[26]對鴨MyoD1基因外顯子1進行多態性檢測,發現1處C/G的突變引起了氨基酸的變化,并且導致鴨屠體腿肌率的顯著增加。王瓊等[27]對優質肉雞研究結果表明,MyoG基因5′調控區存在影響肌纖維生長發育的SNP位點。這些說明MSTN、MyoD1和MyoG基因是影響禽類肌肉生長的潛在候選基因。
氨基酸的總平均疏水性可以體現蛋白質的親疏水性質,數值越高代表疏水能力越強,數值為負值表明該蛋白質為親水性蛋白質[28]。本研究發現,MyoD1突變蛋白質相比原始蛋白質,總平均親水性變小,即疏水性變強,這可能對蛋白質的折疊以及功能的發揮有一定的影響。蛋白質的不穩定系數小于40,該蛋白質可認為是穩定的;不穩定系數大于40,那么該蛋白質為不穩定蛋白質[29]。MSTN、MyoD1和MyoG野生型蛋白質和突變型蛋白質不穩定系數均大于40,說明這些蛋白質均為不穩定蛋白質,在動物機體內的半衰期時間較短。
基因外顯子水平上的有義突變可以影響蛋白質的二級結構。本試驗涉及的3個基因各有1處有義突變,通過蛋白質二級結構預測發現,這些有義突變導致的氨基酸變化均影響了蛋白質的二級結構。其中MSTN蛋白質2處連續的折疊和無規則卷曲均變成了螺旋結構;MyoD1蛋白質1處無規則卷曲變成了螺旋結構,1處折疊變成了無規則卷曲;MyoG蛋白質3處螺旋結構均變成了無規則卷曲。
參考文獻:
[1]RUDNICKI M, SCHNEGELSBERG P, STEAD R, et al. MyoD or Myf-5 is required for the formation of skeletal muscle. Cell, 1993, 75(7): 1351-1359.
[2]GROBET L, MARTIN L, PONCELET D, et al. A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle. Nature Genetics,1997, 17(1): 71-74.
[3]KAMBADUR R, SHARMA M, SMITH T. Mutations in myostatin (GDF8) in double muscled Belgian blue and Piedmon tese cattle. Genome Research, 1997(7): 910-916.
[4]WHITTEMORE L, SONG K, LI X, et al. Inhibition of myostatin in adult mice increases skeletal muscle mass and strength. Biochemical and Biophysical Research Communications, 2003, 300(4): 965-971.
[5]MCPHERRON A C, LEE S J. Double muscling in cattle due to mutations in the myostatin gene. Proc Natl Acad Sci USA, 1997(94): 12457-12461.
[6]NISHI M, YASUE A, NISHIMATU S, et al. A missense mutant myostatin causes hyperplasia without hypertrophy in the mouse muscle. Biochemical and Biophysical Research Communications, 2002, 293(1): 247-251.
[7]CLOP A, MARCQ F, TAKEDA H, et al. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nature Genetics, 2006, 38: 813-818.
[8]BOMAN I A, KLEMETSDAL G, NAFSTAD O, et al. Impact of two myostatin (MSTN) mutations on weight gain and lamb carcass classification in Norwegian white sheep (Ovisaries). Genetics Selection Evolution, 2010, 42(1): 4.
[9]MOSHER D S, QUIGNON P, BUSTAMANTE C D, et al. A mutation in the myostatin gene increases muscle mass and enhancesracing performance in heterozygote dogs. Plos Genetics, 2007, 3(5): 779-785.
[10]張跟喜,丁馥香,是燕萍,等. 肌肉生長抑制素基因 (MSTN) 外顯子1的多態性及其與邊雞生長性狀的關聯分析. 農業生物技術學報, 2011, 19(1): 122-127.
[11]王志新. 蛋白質結構預測的現狀與展望. 生命的化學, 1998, 18(6): 19-22.
[12]吳祖建,高芳鑾,沈建國. 生物信息學分析實踐. 北京: 科學出版社, 2010.
[13]REMIGNON H, GARDAHAUT M F, MARCHE G, et al. Selection for rapid growth increases the number and the size of muscle fibres without changing their typing in chickens. J Muscle Res Cell Motil, 1995, 16(2): 95-102.
[14]GROUNDS M D. Towards understanding skeletal muscle regeneration. Pathol Res Pract, 1991, 187(1): 1-22.
[15]WEINTRAUB H, TAPSCOTT S J, DAVIS R L, et al. Activation of muscle-specific genes in pigment, nerve, fat, liver, and fibroblast cell lines by forced expression of MyoD. PNAS,1989, 86(14): 5434-5438.
[16]NAIDU P S, LUDOLPH D C, TO R Q, et al. Myogenin and MEF2 function synergistically to activate the MRF4 promoter during myogenesis. Mol Cell Biol,1995, 15(5): 2707-2718.
[17]OLSON E N, BRENNAN T J, CHAKRABORTY T, et al. Molecular control of myogenesis: antagonism between growth and differentiation. Mol Cell Biochem, 1991, 104(1-2): 7-13.
[18]JOULIA D, BERNARDI H, GARANDEL V, et al. Mechanisms involved in the inhibition of myoblast proliferation and differentiation by myostatin. Experimental Cell Research, 2003, 286(2): 263-275.
[19]LANGLEY B, THOMAS M, BISHOP A. Myostatin inhibits myoblast differentiation by down-regulating MyoD expression. Journal of Biological Chemistry, 2002, 277(51): 49831-49840.
[20]阮井玲,甄鑫,劉娣,等. Myostatin通過Smad3下調MyoD的表達來抑制骨骼肌衛星細胞的分化. 中國生物工程雜志, 2008, 28(5): 99-103.
[21]SPILLER M P, KAMBADUR R, JEANPLONG F, et al. The myostatin gene is a downstream target gene of basic helix-loop-helix transcription factorMyoD. Molecular and Cellular Biology, 2002, 22(20): 7066-7082.
[22]XU T S, GU L H, ZHANG X H, et al. Characterization of myostatin gene (MSTN) of Pekin duck and the association of its polymorphism with breast muscle traits. Genet Mol Res, 2013, 12(3): 3166-3177.
[23]LU J, HOU S, HUANG W, et al. Polymorphisms in the myostatin gene and their association with growth and carcass traits in duck. African Journal of Biotechnology, 2011, 10(54): 11309-11312.
[24]盧俊清. 北京鴨肌肉生長抑制素基因(MSTN)多態性與骨骼肌和脂肪生長發育性狀的關聯性分析. 中國: 中國農業科學院, 2008.
[25]劉倩,陳永華,蔡鳳仙,等. 高郵鴨生長抑制素基因外顯子3多態性與腹脂率的關聯分析. 中國家禽, 2012, 34(23): 27-30.
[26]寇潔,李亮,王繼文. 5個鴨品種MyoD基因外顯子1的多態性及其與屠宰性狀的關聯分析. 中國畜牧雜志, 2011, 47(19): 10-12.
[27]王瓊,劉益平,蔣小松,等.MyoG基因多態性與優質肉雞屠宰性狀和肉質性狀的相關性分析. 遺傳, 2007, 29(9): 1089-1096.
[28]薛慶中. DNA和蛋白質序列數據分析工具. 3版. 北京: 科學出版社, 2012.
[29]WALKER J M. The proteomics protocols handbook.Totowar: Humana Press, 2005.
(責任編輯:袁偉)
胡成,朱善元,左偉勇,等. 鴨I型肝炎病毒一步法RT-LAMP可視化快速檢測方法的建立[J].江蘇農業學報,2015,31(3):613-618.
