小麥產量構成因素的基因效應和雜種優勢分析
2015-01-01 02:14:36姚金保周淼平馬鴻翔楊學明
江蘇農業學報 2015年3期
關鍵詞:質量
姚金保, 楊 丹, 周淼平, 馬鴻翔, 楊學明
(江蘇省農業科學院/江蘇省農業生物學重點實驗室,江蘇 南京 210014)
小麥籽粒產量由單位面積穗數、每穗粒數和千 粒質量3個因素構成[1-3]。育種實踐表明,在保證足夠穗數的基礎上,增加穗粒數和穩定提高千粒質量是今后小麥產量突破的重要途徑[4-5]。近年來,許多學者采用不同方法對小麥每穗粒數和千粒質量的遺傳特性和雜種優勢進行了研究。一般認為小麥每穗粒數和千粒質量的遺傳符合加性-顯性模型,基因作用方式以加性效應為主[6-10],也有以顯性效應為主的報道[11-14]。也有研究者認為小麥每穗粒數或千粒質量的遺傳機制復雜,可能存在上位性效應[15-17]。數量遺傳研究結果表明,小麥的每穗粒數和千粒質量是質量-數量性狀,由主基因和微效基因共同控制[18]。關于小麥每穗粒數和千粒質量雜種優勢的研究結論也不完全一致。多數研究結果表明,小麥每穗粒數和千粒質量的平均雜種優勢在F1代普遍存在一定的正向優勢[19-22]。Inamullah等[23]、Al-Falahy[24]研究發現,每穗粒數和千粒質量的平均雜種優勢變異幅度較大,其優勢強弱因組合而異。許永財等[25]研究認為,千粒質量具有較高的正向超親優勢,但每穗粒數表現負向中親優勢。趙鵬等[26]、史秀秀等[27]的研究結果也證實了每穗粒數表現為負向中親優勢。但 Abdullah等[28]、Ilker等[29]認為,每穗粒數存在明顯的正向超親優勢。由此可見,對于小麥產量構成因素的遺傳特性和雜種優勢還需進一步研究。本試驗以7個產量構成因素差異較大的小麥品種為親本,按n(n-1)/2雙列雜交法配制21個組合,在2種環境下研究每穗粒數和千粒質量2個小麥產量構成因素的基因效應和雜種優勢,旨在為小麥高產育種提供理論依據。
1 材料與方法
1.1 材料
2012年春選用綜合農藝性狀優良、每穗粒數和千粒質量有一定差異的7個冬小麥品種作為親本,7個品種為徐麥25(P1)、矮抗58(P2)、淮麥20(P3)、淮麥33(P4)、煙 2801(P5)、濟麥 22(P6),徐麥 31(P7)。
1.2 試驗設計
按Griffing雙列雜交模型II,配制成21個雜交組合,2013年秋分別在宿遷和南京種植F1及親本,共28個基因型。田間按隨機區組排列,重復3次,每小區2行區,行長1.5 m,行距25 cm,株距4 cm,田間管理措施同大田栽培。
1.3 性狀調查
2014年在小麥成熟前,每小區隨機取30穗,調查每穗粒數;成熟時按小區收獲,脫粒后揚凈、風干,分小區統計千粒質量。
1.4 數據統計分析
雜種優勢測定:平均優勢(%)=[(F1-MP)/MP]×100%,超高親優勢(%)=[(F1-HP)/HP]×100%,超低親優勢(%)=[(F1-LP)/LP]×100%,其中Fl為雜種一代性狀平均值,MP、HP和LP分別表示雙親均值、高值親本值和低值親本值。利用Hayman雙列雜交方法,對2個產量構成因素分試點進行遺傳模型分析并估算遺傳參數[30]。
2 結果與分析
2.1 每穗粒數和千粒質量的遺傳模型分析
2.1.1 方差分析 7個小麥品種及其雙列雜交組配的21個雜交組合的2個產量構成因素在宿遷和南京2個試點上的測定值列于表1。組合間穗粒數在宿遷和南京試點間的變化范圍分別為32.07~47.87粒和 28.63~46.40粒,變異系數分別為11.38%和10.51%;組合間千粒質量在宿遷和南京試點間的變化范圍分別為 38.03~50.33 g和38.07~48.03 g,變異系數分別為6.87%和6.27%,表明所分析的性狀具有較大差異。方差分析結果(表2)表明,每穗粒數和千粒質量基因型間方差均達極顯著水平,而重復間差異均不顯著,說明基因型效應存在顯著差異,即供試材料間的遺傳基礎存在顯著差異,因此可以做進一步的遺傳分析。
2.1.2 加性-顯性模型測驗 每穗粒數系列協方差(Wr)/方差(Vr)回歸分析結果(表3)表明,其直線回歸系數b與0的差異達顯著(宿遷)和極顯著(南京),與1的差異均不顯著,由此可見每穗粒數的遺傳符合加性-顯性模型。由回歸截距α的測驗結果可知,每穗粒數的回歸截距α<0,但與0差異不顯著,說明該性狀屬超顯性遺傳。對南京點試驗的千粒質量Wr、Vr進行回歸分析,得α=0.048,b=0.541。由回歸斜率b的檢驗結果(表3)可以看出,b與0、1間都存在顯著差異,這表明控制千粒質量的基因不僅具有加性、顯性作用,還可能存在基因位點間的互作(上位性效應)。應作Wr-Vr和Wr+Vr方差分析,以進一步確定是否存在上位性效應。由表4可知,千粒質量的Wr+Vr、Wr-Vr值在公共親本間和公共親本內差異均未達到顯著水平,表明不存在上位性效應。宿遷點千粒質量的分析結果(表3)則表明,b與0存在顯著差異,而與1差異不顯著。千粒質量的基因作用方式為加性、顯性作用。綜合2個地點的分析結果,認為千粒質量的遺傳符合加性-顯性模型,不存在上位性效應。

表1 不同組合每穗粒數和千粒質量在2個試點的平均值Table 1 Averages for number of kernels per spike and thousand-grain weight in different crosses at two sites

表2 2個試點每穗粒數和千粒質量的方差分析Table 2 ANOVA for kernel number per spike and thousand-grain weight at two sites
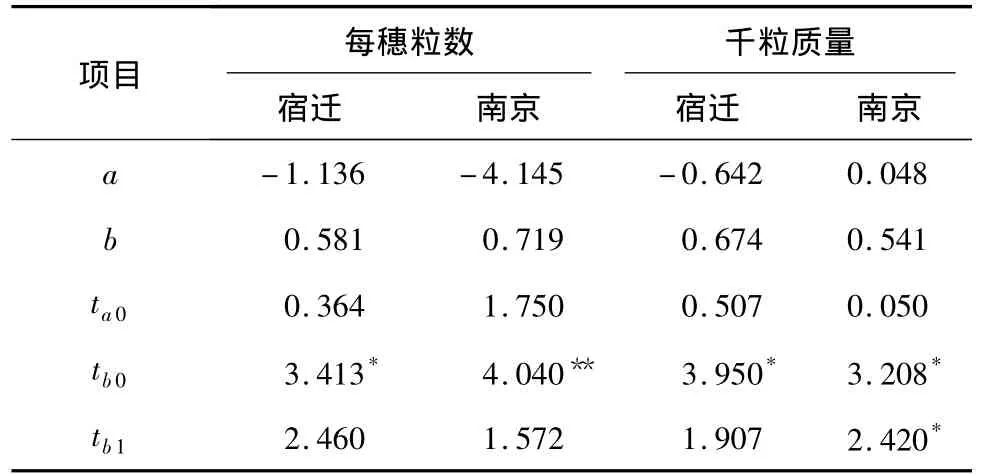
表3 每穗粒數和千粒質量系列協方差(Wr)對方差(Vr)的回歸分析Table 3 Regression analyses of covariance(Wr)on variance(Vr)for kernel number per spike and thousand-grain weight at two sites
2.1.3 親本顯隱性基因的分布及其作用方向分析 由圖1可知,在每穗粒數的遺傳上,各親本具有不等的顯隱性等位基因。其中淮麥33、煙2801距原點最近,含有最多的顯性基因;徐麥25、矮抗58距原點最遠,表明含隱性基因最多。由圖2可知,淮麥20具有較多控制千粒質量遺傳的顯性基因,而其他6個親本在2個試點的試驗中所含顯隱性基因比例不完全一致。由表5可知,每穗粒數和千粒質量的親本均值(Yr)與Wr+Vr的相關系數均呈負相關,表明每穗粒數和千粒質量的遺傳表現為高值受顯性基因控制,低值受隱性基因控制,顯性等位基因對每穗粒數和千粒質量起增效作用,隱性基因則起減效作用。

表4 千粒質量Wr+Vr和Wr-Vr的方差分析(南京點)Table 4 ANOVA for Wr+Vrand Wr-Vrof thousand-grain weight in Nanjing experiment
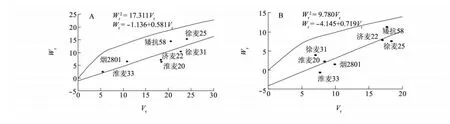
圖1 宿遷(A)、南京(B)試點每穗粒數Wr依Vr的回歸直線Fig.1 Regression of Wron Vrfor kernel number per spike in Suqian(A)and Nanjing(B)
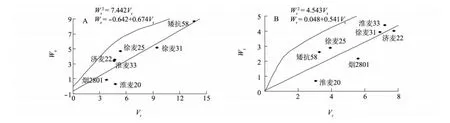
圖2 宿遷(A)、南京(B)試點千粒質量Wr依Vr的回歸直線Fig.2 Regression of Wron Vrfor thousand-grain weight in Suqian(A)and Nanjing(B)
2.1.4 遺傳參數的估算 表5結果表明,每穗粒數和千粒質量的加性效應方差D和顯性效應方差H1、H2均達極顯著水平,說明每穗粒數和千粒質量的遺傳受加性效應和顯性效應共同作用;由于H1遠大于D,表明在每穗粒數和千粒質量的遺傳中,基因的顯性作用較加性作用更重要。每穗粒數的平均顯性度分別為1.715(宿遷點)和2.047(南京點),千粒質量的平均顯性度分別為1.576(宿遷點)和1.787(南京點),顯性程度均為超顯性。每穗粒數和千粒質量的H2/4H1值均<0.25,表明親本中每穗粒數和千粒質量的增、減效等位基因分布不對稱;由顯性和隱性基因之比[(4DH1)1/2+F]/[(4DH1)1/2-F]均為<1可知,控制親本的隱性基因頻率大于顯性基因頻率。每穗粒數和千粒質量在2個試點的平均狹義遺傳力分別為54.00%和63.17%。
2.2 每穗粒數和千粒質量的雜種優勢
由表6可見,盡管每穗粒數和千粒質量在2試點間都出現了中親、超高親和超低親優勢為負值的一些組合,但兩性狀的超低親優勢、平均中親優勢和超高親優勢均為正值,這表明每穗粒數和千粒質量主要表現正向雜種優勢,且表現為趨高親遺傳。每穗粒數和千粒質量的3種雜種優勢變異幅度都較大,并在組合間、地點間以及基因型與地點互作間均存在極顯著差異(表7)。因此,每穗粒數和千粒質量的雜種優勢表現受基因型、環境和基因型與環境互作等因素的共同影響。

表6 每穗粒數和千粒質量的雜種優勢Table 6 Heterosis of kernel number per spike and thousand-grain weight

表7 每穗粒數和千粒質量超高親優勢聯合方差分析Table 7 Pooled analysis of variance of super-parent heterosis for kernel number per spike and thousand-grain weight
3 討論
3.1 關于小麥穗粒數和千粒質量的遺傳特點
不少學者對小麥穗粒數和千粒質量的遺傳規律、基因作用方式等進行了研究。姚金保等[31]早期的研究結果表明,小麥每穗粒數和千粒質量遺傳控制由加性和非加性效應共同起作用,但以加性為主。Inamullah等[8]的研究結果也得到相似的結論。本試驗結果表明,小麥每穗粒數和千粒質量受加性效應和顯性效應2種遺傳效應的共同作用,顯性作用相對較大,加性作用相對較小,這與張利華等[11]、Hussain 等[12]、Akram 等[13]的研究結果相符。總之對小麥每穗粒數和千粒質量的遺傳研究中,關于加性和顯性共同起作用這一觀點是基本一致的,但在加性與顯性的相對重要性方面尚有分歧。這些分歧可能與試驗環境、供試材料的不同有關。
3.2 關于環境對小麥穗粒數和千粒質量的影響
小麥產量性狀是復雜的數量性狀,除受控于基因型外,還在一定程度上受環境因子以及基因型與環境因子互作等的影響[32-33]。本研究中,有關試驗結果在2個地點間存在一定差異,宿遷點的每穗粒數和千粒質量分別比南京點高2.25粒和1.20 g。這種差異主要是由于種植地點的氣候條件(溫度、光照、降水)、土壤以及栽培措施的不同所引起。Reynolds等[34]認為每穗粒數的減少與二棱期至開花的時間縮短有關。Ishag等[35]研究認為,千粒質量與灌漿期間的平均氣溫呈負相關,且溫度每上升1℃,千粒質量下降4 mg。Borner等[36]檢測到2個與每穗粒數相關的QTL以及3個與千粒質量相關的QTL;Heidari等[37]檢測到2個控制每穗粒數的主效QTL和6個控制千粒質量的主效QTL,具有基因型-環境互作效應。因此,在小麥高產育種實踐中,應在不同生態區對育種材料進行多點鑒定試驗,篩選在不同環境條件下表現均穩定的優良基因型。在推廣高產小麥品種時,還需要采用配套栽培技術,按照品種的適應范圍進行區域化種植和生產。
3.3 關于小麥穗粒數和千粒質量的遺傳改良
對產量構成因素雜種優勢的分析結果表明,每穗粒數和千粒質量均表現正向雜種優勢,且表現為趨高親遺傳,而且優勢變幅都較大,這與Ilker等[29]對每穗粒數和Inamullah等[23]對千粒質量的研究結果相似。因此,通過正確選擇雙親,利用其正向優勢來協調提高穗粒數和千粒質量進而篩選出產量潛力更高的小麥品種是可能的。本研究結果表明,小麥每穗粒數和千粒質量的遺傳符合加性-顯性模型,基因的顯性作用較加性作用重要。高每穗粒數和千粒質量表現高值為顯性,低值為隱性。有關研究結果對小麥產量構成因素的遺傳改良可能有一定參考價值。在育種早代,由于高每穗粒數和千粒質量基因型含顯性基因較多,且處于雜合分離狀態,因此必須通過連續多代的定向選擇才能獲得理想的材料。在本研究中,淮麥33和煙2801是含高穗粒數顯性基因較多的2個親本,淮麥20是含高千粒質量顯性基因較多的親本,這3個親本其他農藝性狀和抗病性表現也較好,在小麥高產育種中,應進一步利用這3個優良親本。
[1] 吳宏亞,蔣正寧,王搖玲,等.小麥籽粒硬度及其對面粉加工品質影響的研究進展[J].江蘇農業學報,2014,30(2):437-441.
[2] 蔣小忠,封超年,郭文善.磷肥種類對弱筋小麥籽粒產量和蛋白質含量的影響[J].江蘇農業科學,2013,41(12):63-66.
[3] 蔡金華,陳愛大,李東升.不同施氮條件下鎮麥168籽粒產量與構成因素的相關性研究[J].江蘇農業科學,2014,42(10):73-75.
[4] HE Z H,RAJARAM S,XIN Z Y,et al.A history of wheat breeding in China[M].Mexico DF:CIMMYT,2001.
[5] 田紀春,王延訓.超級小麥的育種策略與實踐[J].作物雜志,2005(4):67-68.
[6] TAHIR M S,ALAM K,CHOWDHRY M A,et al.Genetic analy-sis of some important economic traits in bread wheat(Triticum aestivum L.)[J].Pakistan Journal of Agricultural Sciences,1995,32:172-177.
[7] KHAN A S,KHAN M K R,KHAN T M.Genetic analysis of plant height,grain yield and other traits in wheat(Triticum aestivum L.)[J].International Journal of Agriculture&Biology,2000,2:129-132.
[8] INAMULLAH,AHMAD H,MOHAMMAD F,et al.Diallel analysis of the inheritance pattern of agronomic traits of bread wheat[J].Pakistan Journal of Botany,2006,38(4):1169-1175.
[9] NAZEER W,ALI Z,ALI A,et al.Genetic behavior for some polygenic yield contributing traits in wheat(Triticum aestivum L.)[J].Journal of Agricultural Research,2010,48(3):267-276.
[10] CHEN D,ZHANG J P,WANG J S,et al.Inheritance and availability of high grain number per spike in two wheat germplasm lines[J].Journal of Integrative Agriculture,2012,11(9):1409-1416.
[11]張利華,許梅芬.小麥收獲指數和其它幾個農藝性狀的基因效應分析[J].核農學報,1997,11(3):135-140.
[12] HUSSAIN F,SIAL R A,ASHRAF M.Genetic studies for yield and yield related traits in wheat under leaf rust attack[J].International Journal of Agriculture&Biology,2008,10:531-535.
[13] AKRAM Z,AJMAL S U,SHABBIR G,et al.Inheritance mechanism of some yield components in bread wheat Journal[J].Pakistan Journal of Agricultural Research,2009,22:1-8.
[14] RABBANI G,MAHMOOD A,SHABBIR G,et al.Gene action in some yield attributes of bread wheat under two water regimes[J].Pakistan Journal of Botany,2011,43(2):1141-1156.
[15] RAHMAN M A,SIDDQUIE N A,ALAM M R,et al.Genetic analysis of some yield contributing and quality characters in spring wheat(Triticum aestivum)[J].Asian Journal of Plant Sciences,2003,2(3):277-282.
[16]姚金保,任麗娟,張平平,等.小麥產量構成因素的雙列雜交分析[J].核農學報,2011,25(4):633-638.
[17] AHMAD F,KHAN S,AHMAD S Q,et al.Genetic analysis of some quantitative traits in bread wheat across environments[J].African Journal of Agricultural Research,2011,6(3):686-692.
[18]李斯深,陳茂學,王洪剛.利用重組自交系(RILs)群體進行質量-數量性狀的遺傳分析—遺傳模型和小麥產量性狀遺傳[J].作物學報,2001,27(6):896-904.
[19] RASUL I,KHAN A S,ALI Z.Estimation of heterosis for yield and some yield components in bread wheat[J].International Journal of Agriculture&Biology,2002,4:214-216.
[20]冀天會,張燦軍,楊子光,等.水旱生態型冬小麥品種雜種優勢分析[J].麥類作物學,2003,23(4):19-22.
[21] BAO Y G,WANG S,WANG X Q,et al.Heterosis and combining ability for major yield traits of a new germplasm Shannong 0095 derived from Thinopyrum intermedium[J].Agricultural Sciences in China,2009,8(6):753-760.
[22]王玉斌,逯臘虎,武計萍,等.冬小麥主要農藝性狀的雜種優勢和配合力分析[J].山西農業科學,2014,42(7):653-656.
[23] INAMULLAH,AHMAD H,MOHAMMAD F,et al.Evaluation of the heterotic and heterobeltiotic potential of wheat genotypes for improved yield[J].Pakistan Journal of Botany,2006,38(4):1159-1167.
[24] AL-FALAHY M A H.Estimate of genetic parameters of grain yield and some agronomic traits in durum wheat using diallel crosses[J].Journal of Agricultural Science and Technology,2013,3:26-34.
[25]許永財,相吉山.春小麥主要農藝性狀雜種優勢與配合力研究[J]. 青海大學學報:自然科學版,2007,25(2):4-9.
[26]趙 鵬,彭惠茹,倪中福,等.穗分枝小麥與普通小麥雜交組合主要性狀的雜種優勢與配合力分析[J].麥類作物學報,2009,29(2):212-216.
[27]史秀秀,畢曉靜,馬守才,等.黃淮麥區雜交小麥親本的雜種優勢和配合力分析[J].麥類作物學報,2013,33(6):1111-1118.
[28] ABDULLAH G M,KHAN A S,ALI Z.Heterosis study of certain important traits in wheat[J].International Journal of Agriculture& Biology,2002,4(3):326-328.
[29] ILKER E,TONK F A,TOSUN M.Heterosis for yield and its components in bread wheat crosses among powdery mildew resistant and susceptible genotypes[J].Pakistan Journal of Botany,2010,42(1):513-522.
[30]劉來福,毛盛賢,黃遠樟.作物數量遺傳[M].北京:農業出版社,1984:125-149.
[31]姚金保,王書文,姚國才,等.小麥產量構成因素的遺傳分析[J].上海農業學報,2004,20(1):45-48.
[32]常 磊,柴守璽,楊德龍,等.我國旱地春小麥產量及主要農藝指標的變異分析[J].應用生態學報,2010,21(11):2821-2829.
[33] SANCHEZ-GARCIAA M,ALVAROA F,MARTIN-SANCHEZA J A,et al.Breeding effects on the genotype×environment interaction for yield of bread wheat grown in Spain during the 20th century[J].Field Crops Research,2012,126:79-86.
[34] REYNOLDS M P,CALDERINI D F,CONDON A G,et al.Physiological basis of yield gains in wheat associated with the LR19 translocation from Agropyron elongatum[J].Euphytica,2001,119:139-144.
[35] ISHAG H M,MOHAMED B A.Phasic development of spring wheat and stability of yield and its components in hot environments[J].Field Crops Research,1996,46:169-176.
[36] BORNER A,SCHUMANN E,FURSTE A,et al.Mapping of quantitative trait loci determining agronomic important characters in hexaploid wheat(Triticum aestivum L.)[J].Theoretical and Applied Genetics,2002,105:921-936.
[37] HEIDARI B,SAYED-TABATABAEI B E,SAEIDI G,et al.Mapping QTL for grain yield,yield components,and spike features in a doubled haploid population of bread wheat[J].Genome,2011,54:517-527.
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
