霍亂弧菌中環腺苷酸受體蛋白對高絲氨酸脫氫酶基因的調控作用
2014-11-29 04:16:00蔡紅艷陳保立梁未麗王多春李杰段國賢闞飆
生物技術通訊 2014年4期
蔡紅艷 ,陳保立,梁未麗,王多春,李杰,段國賢,闞飆
1.河北聯合大學 基礎醫學院,河北 唐山 063000;2.山東省疾病預防控制中心,山東省傳染病預防控制重點實驗室,山東 濟南 250014;3.中國疾病預防控制中心 傳染病預防控制所,傳染病預防控制國家重點實驗室,北京 102206
細菌中的環腺苷酸受體蛋白(cAMP receptor protein,CRP)是一種全局性調控因子,最早在大腸桿菌分解代謝產物阻遏過程中被鑒定出[1]。CRP 與cAMP 形成cAMP-CRP 聚合體后與啟動子上游保守序列TGTGA-(N6)-TCACA 結合,從而調控基因的轉錄[1-2]。在大腸桿菌中CRP 的調控作用已經得到較深入的研究,已發現至少有378 個基因受到CRP的調控[3],轉錄組分析數據表明CRP 對碳源核心代謝通路上的酶及轉運載體的轉錄有調控作用[4]。霍亂弧菌CRP 的基因和蛋白序列與大腸桿菌的CRP分別有81%和95%的一致性[5-6],這提示CRP 在霍亂弧菌與大腸桿菌中有著相似的作用。CRP在霍亂弧菌中功能的研究主要集中在其對毒力基因的調控上,如CRP 調控霍亂毒素和菌素共調節菌毛基因的轉錄[7]、調控血凝素/蛋白酶的產生[8]。另外,CRP 還影響密度感應系統及霍亂弧菌在腸道中生存所必需基因的轉錄[9]。
我們曾利用表型芯片檢測、轉錄水平分析、調控蛋白結合DNA 調控區的實驗方式,分析了霍亂弧菌中CRP 對不同碳源利用的調節作用,找到24 種被CRP 調控利用的碳源,其中11 種未在細菌中見到受CRP 調控的報道[10]。其中,發現參與L-天冬氨酸代謝的VC0391(編碼天冬氨酸激酶,aspartokinase isozymes,AK)和VC2364(編碼高絲氨酸脫氫酶,homoserine dehydrogenase,HDH)基因受CRP 的調控。天冬氨酸是一種α-氨基酸,其異構物L-天冬氨酸是20種蛋白氨基酸之一。天冬氨酸代謝通路廣泛存在于植物及微生物中,涉及的步驟及參與的酶均比較保守。在大腸桿菌中,天冬氨酸在AK的作用下生成磷酸化天門氨酰。共有3 類AK,其中AKⅠ和AKⅡ是雙功能酶,酶的活性分別受到蘇氨酸和甲硫氨酸的反饋抑制;AKⅢ為單功能酶,受到賴氨酸的反饋抑制。磷酸化天門氨酰在天冬氨酸-β-半醛脫氫酶(aspartate-β-semialdehyde dehydrogenase,ASADH)的作用下生成天冬氨酸-β-半醛,而后在HDH 的作用下生成高絲氨酸,進而生成甲硫氨酸、蘇氨酸和異亮氨酸,或者在二氫吡啶二羧酸合成酶(dihydrodipicolinate synthase)的作用下生成四氫吡啶二羧酸,進而生成賴氨酸。
我們的前期研究證實,參與L-天冬氨酸代謝的VC0391基因啟動子區與CRP 直接結合,雖看到CRP對編碼HDH 的VC2364 基因的調控效果,但未發現VC2364 基因調控區典型的CRP 結合位點及CRP 的直接結合作用[10]。在本研究中,我們進一步分析了CRP對VC2364基因的調控作用,發現將霍亂弧菌中VC2364 基因啟動子區非典型的CRP 結合序列該造為典型序列后,增強了受CRP 調控的作用,顯示原VC2364基因序列能促使CRP的弱調控,提示霍亂弧菌中可能存在的受CRP精細調節的作用。
1 材料和方法
1.1 材料
菌株與質粒見表1。大腸桿菌與霍亂弧菌在LB培養基中于37℃培養,培養基中加入合適的抗生素。大腸桿菌培養所加氯霉素(Cm)終濃度為10 mg/mL;霍亂弧菌所加Cm 終濃度為2 mg/mL,多黏菌素B終濃度為0.1 U/mL。
PCR 產物純化試劑盒、質粒提取試劑盒、瓊脂糖凝膠回收試劑盒購自Omega 公司;DNA 提取試劑盒、PCR 擴增用2×Pfumaster Mix 購自天根公司;限制性內切酶、T4DNA 連接酶購自TaKaRa 公司;酶聯檢測儀Infinite M200 Pro 購自Tecan 公司;細胞培養板購自Corning 公司;PCR 儀、電泳儀、電泳槽、凝膠成像儀購自Bio-Rad 公司;生物安全柜Forma ClassⅡ,A2,購自Thermo公司。
1.2 CRP結合位點預測與搭橋PCR
分別用Virtual Footprint[11]和Regulatory Sequence Analysis Tools(RSAT)[12]軟件預測可能的CRP 結合位點。為了將VC2364 基因的啟動子序列中與CRP 結合位點保守序列不一致的3 個堿基替換成保守序列,我們在預測堿基位點上下游500 bp 同源臂處設計引物P1、P2、P3、P4,通過P2、P3 的設計將不一致的序列置換為CRP 保守序列。在引物P2、P3 的5'端各取10 個堿基反向互補,將P2 反向互補序列加在P3 的5'端,P3 反向互補序列加在P2 的5'端,并在P1、P4 引物的5'端加上酶切位點與保護堿基。以C7258 基因組DNA 為模板,分別以引物P1、P2和P3、P4 擴增得到同源臂,將擴增得到的2 個PCR 產物片段按1∶1 的比例混合后純化回收,以純化回收的PCR 產物為模板,用引物P1、P4 進行搭橋PCR 擴增,得到上下游同源臂搭橋序列片段。引物序列見表2。
1.3 質粒的構建
未改造的PCR 產物、搭橋PCR 產物、pBBRlux 質粒分別用PvuⅠ和SacⅠ雙酶切后切膠回收,酶切產物用T4DNA 連接酶連接1 h,取5μL 轉入100μL大腸桿菌DH5α感受態細胞中,冰浴30 min,42℃熱激90 s,加入1 mL液體LB,37℃搖床振搖40 min后離心,取100μL液體涂布于含Cm的平板培養基上,待長出菌落后,挑取50 個菌落用pBBRlux-PvUⅠ、pBBRLux-BamHⅠ引物進行PCR擴增、測序,挑取陽性菌落,將構建成功的質粒命名為pvc2364-lux、pvc2364s-lux。

表1 研究使用的菌株與質粒

表2 質粒構建所用引物及序列
1.4 接合轉移
將質粒pvc2364-lux、pvc2364s-lux 分別轉 入SM10 供體菌、受體菌(C7258、WL7258),各取0.6 mL 菌液混合,5000 r/min 離心5 min,棄上清,用1 mL LB 懸菌后再次離心,棄上清,用80μL LB 懸菌,將菌液點到高壓滅菌并平貼于LB 平板上的0.45μm 的微孔水系濾膜上,菌液吸收后于37℃溫箱中培養4~6 h,用LB 沖洗濾膜,收集洗液,離心后沉淀重懸于適量LB,涂布于質粒所帶抗性多黏菌素B和霍亂弧菌所帶抗性Cm 的培養基平板,37℃培養過夜。
1.5 熒光值的測定
將含有pvc2364-lux,pvc2364s-lux 質粒的霍亂弧菌活化過夜,接種于含Cm和多黏菌素B 的5 mL液體培養基中,第2 d 按1/1000 的比例轉接到2 mL無菌LB 中,37℃振蕩培養,到一定時間分別取200μL 菌液轉移到96 孔板中,于Tecan Infinite M200 Pro酶標儀中檢測熒光值,同時在該儀器中測定菌濁度(D600nm值),以時間點為橫坐標、熒光值/D600nm為縱坐標,得到光值曲線。
2 結果
2.1 CRP結合位點預測
分別用Virtual Footprint和RSAT 軟件預測基因VC2364 上游是否有CRP 可能的結合位點。Virtual Footprint預測結果顯示VC2364基因上游431~452處有可能的CRP 結合位點,與保守序列有7 個位點一致;RSAT 未預測到VC2364 上游有CRP 可能的結合位點。
2.2 質粒構建
用搭橋PCR 的方法,將VC2364 基因啟動子上游序列中與CRP 結合位點保守序列不一致的3 個堿基替換成保守序列,未改造的序列經PCR 擴增后與pBBRlux 連接并導入感受態大腸桿菌DH5α,挑取PCR 擴增陽性的菌落,提取質粒進行酶切驗證,選取酶切條帶大小合適的質粒測序,結果顯示構建的報告質粒正確。圖1為改造前與改造后的序列對比。
2.3 報告質粒在不同菌株中熒光值的測定
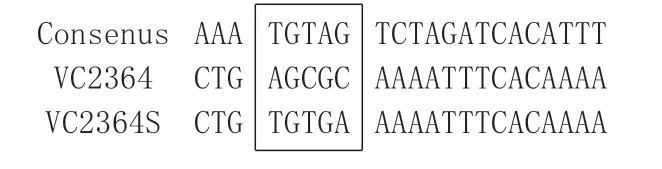
圖1 基因VC2364啟動子CRP結合位點堿基的改造
質粒pBBRlux 利用luxCDABE 基因作為報告基因,其熒光表達需要借助插入的啟動子。將VC2364基因啟動子上游可能的CRP結合位點原始序列和改造序列構建到報告質粒pBBRlux 中,檢測報告質粒pBBRlux 在C7258 與WL7258 菌株中的熒光值,發現改造的質粒在C7258 株中的熒光值高于未改造的,在WL7258 株中的差異更明顯。另外,序列改造與未改造的質粒在CRP野生株C7258中的熒光值均高于CRP突變株WL7258中。見圖2。
3 討論
本研究論證了細菌中CRP調控單元中一個CRP結合位點序列的特殊例子。我們在分析CRP對霍亂弧菌碳源代謝的影響中,發現了多種碳源的代謝受到CRP 調控,參與L-天冬氨酸代謝的VC2364 基因在轉錄水平上受CRP 的調控,但其啟動子區未發現典型的CRP結合位點序列。CRP與啟動子區保守序列TGTGA-(N6)-TCACA 結合從而調控基因的轉錄,軟件預測發現在VC2364基因啟動子區上游有可能的CRP 結合位點AGCGC-(N6)-TCACA,因此這是一個不典型的CRP 結合序列,與典型序列相比有7個堿基序列一致。但從前期研究結果來看,這7個堿基序列很可能已促進了CRP的結合和調控。為了闡明CRP 是否直接調控VC2364 的轉錄,我們利用報告質粒比較了其啟動子在CRP野生株和缺失株中的轉錄表達,發現在野生株中VC2364啟動子序列啟動的表達是受CRP 調節的,因為其在CRP 野生株中的表達高于在CRP缺失株中。這是對我們前期研究結果的確認。我們也看到,在缺失株中也有啟動子序列啟動的表達,這說明該啟動子同時受到其他因素的調控。
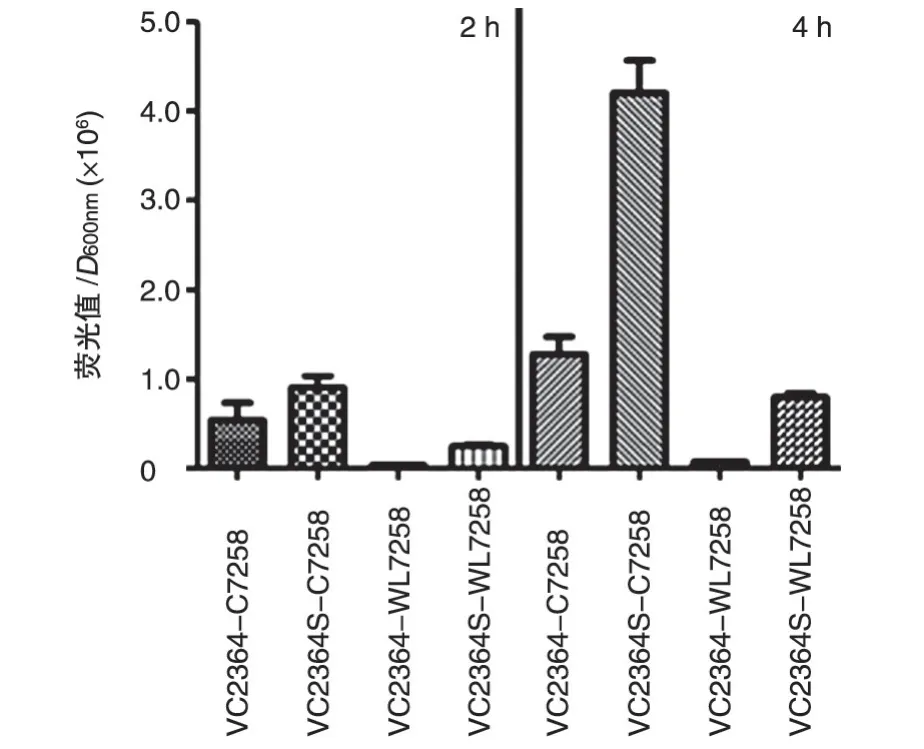
圖2 報告質粒在CRP野生株C7258及其突變株WL7258中熒光值的差異
CRP 識別調控區的典型結合位點序列是TGTGA-(N6)-TCACA,在VC2364 基因前面僅有7 個堿基與經典結合位點一致,為此我們改構了VC2364啟動子區的該區域序列,使其形成典型的CRP 結合位點。實驗測定證實,改造為典型CRP結合序列后,啟動子受CRP 的作用大為增強,改造序列在CRP 缺失株中的熒光變化不大,但在野生株中明顯增強,而且改造的啟動子在CRP 野生株中,其促進表達的熒光也超過了VC2364的野生型啟動子的作用。因此,這些結果提示,VC2364啟動子區非典型的CRP結合序列AGCGC-(N6)-TCACA 也是能被CRP 作用的,但改造為典型序列后受到CRP 更強的作用,激發了更強的轉錄作用。在細菌中,CRP被別構效應物cAMP激活并與之結合形成二聚體復合物,該復合物結合于靶啟動子上游特異的DNA 位點TGTGA-(N6)-TCACA 后與RNA聚合酶直接作用,增強后者在啟動子上的結合能力,從而起始轉錄。我們推測,VC2364啟動子區的這個非典型CRP結合序列,依然能夠使CRP二聚體形成較微弱的結合并發揮一定的激活作用。
在大部分植物和微生物中,天冬氨酸代謝是合成賴氨酸、蘇氨酸、異亮氨酸、甲硫氨酸所必需的,另外該途徑還產生許多重要的中間產物,如在細胞壁合成過程中起交聯作用的二氨基庚二酸及革蘭陽性菌芽胞的重要組成成分吡啶二羧酸[13]。在霍亂弧菌中,天冬氨酸代謝涉及的高絲氨酸脫氫酶基因VC2364啟動子CRP結合位點可能發生了突變,形成了非典型結合序列,弱化了被CRP 調控的作用。為什么沒有出現典型的CRP結合序列?這種序列變化弱化了CRP的調控作用,但這種突變卻被保留,其中的機制目前還不清楚,是否是適應某類微環境而形成的適應性改變,還需要進一步的實驗分析。
[1]Kolb A,Busby S,Buc H,et al.Transcriptional regulation by cAMP and its receptor protein[J].Annu Rev Biochem,1993,62:749-795.
[2]Busby S,Ebright R H,Transcription activation by catabolite activator protein(CAP)[J].J Mol Biol,1999,293(2):199-213.
[3]Shimada T,Fujita N,Yamamoto K,et al.Novel roles of cAMP receptor protein(CRP) in regulation of transport and metabolism of carbon sources[J].Plos One,2011,6(6):e20081.
[4]Gosset G,Zhang Z,Nayyar S,et al.Transcriptome analysis of Crp-dependent catabolite control of gene expression in Escherichia coli[J].J Bacteriol,2004,186(11):3516-3524.
[5]Zhan L,Han Y,Yang L,et al.The cyclic AMP receptor protein,CRP,is required for both virulence and expression of the minimal CRP regulon in Yersinia pestis biovar microtus[J].Infect Immun,2008,76(11):5028-5037.
[6]Espert S M,Elsinghorst E A,Munson G P.The tib adherence locus of enterotoxigenic Escherichia coli is regulated by cyclic AMP receptor protein[J].J Bacteriol,2011,193(6):1369-1376.
[7]Skorupski K,Taylor R K.Cyclic AMP and its receptor protein negatively regulate the coordinate expression of cholera toxin and toxin-coregulated pilus in Vibrio cholerae[J].Proc Natl Acad Sci USA,1997,94(1):265-270.
[8]Silva A J,Benitez J A.Transcriptional regulation of Vibrio cholerae hemagglutinin/protease by the cyclic AMP receptor protein and RpoS[J].J Bacteriol,2004,186(19):6374-6382.
[9]Liang W,Pascual-Montano A,Silva A J,et al.The cyclic AMP receptor protein modulates quorum sensing,motility and multiple genes that affect intestinal colonization in Vibrio cholerae[J].Microbiology,2007,153(Pt 9):2964-2975.
[10]Chen B,Liang W,Wu R,et al.Phenotype microarray screening of carbon sources used by Vibrio cholerae identifies genes regulated by the cAMP receptor protein[J].Can J Microbiol,2013,59(7):472-478.
[11]Munch R,Hiller K,Grote A,et al.Virtual footprint and PRODORIC:an integrative framework for regulon prediction in prokaryotes[J].Bioinformatics,2005,21(22):4187-4189.
[12]Turatsinze J V,Thomas-Chollier M,Defrance M,et al.Using RSAT to scan genome sequences for transcription factor binding sites and cis-regulatory modules[J].Nat Protoc,2008,3(10):1578-1588.
[13]Viola R E.The central enzymes of the aspartate family of amino acid biosynthesis[J].Acc Chem Res,2001,34(5):339-349.
