水稻病程相關基因OsPR1b啟動子的表達特性研究
2014-11-29 04:15:56虞飛博周潔王栩鳴楊勇余初浪程曄嚴成其陳劍平
生物技術通訊 2014年4期
關鍵詞:水稻
虞飛博 ,周潔,王栩鳴,楊勇,余初浪,程曄,嚴成其,陳劍平
1.浙江師范大學 生化學院,浙江 金華 321000;2.浙江省農業科學院 省部共建國家重點實驗室培育基地,農業部植物保護與生物技術重點實驗室,浙江省植物病毒重點實驗室,浙江 杭州 310021
水稻(Oryza sativa)是世界上最重要的農作物之一,為全球半數以上人口特別是發展中國家提供糧食和蛋白質的來源。然而,由于農作物單一的種植方式使它的生長過程更容易受到外界病原菌的影響,而干旱和鹽漬是影響世界水稻產量的兩個主要的非生物因素。
水稻在受病原菌感染、物理傷害和防衛相關信號等刺激后,會誘導自身的抗病防衛體系,從而激活表達下游相應的病程相關(pathogenesis-related,PR)基因。PR基因的誘導表達可以增強植株的抗病性,在水稻病害防御相關的研究中常被作為特征分子標記而廣泛使用[1]。同時,PR基因在衰老、逆境及生長發育過程中也發揮著重要作用。由于PR 蛋白是在植物受到脅迫后誘導合成的一類蛋白,它的表達關鍵在于轉錄水平[2]。在擬南芥和煙草中,關于PR基因的誘導表達調控機制已有較多報道[2-5],對于它們啟動子的活性及其順式作用元件和轉錄因子的作用也分析得較為詳盡。而水稻作為單子葉植物研究的模式生物,了解PR基因的表達調控及功能,對研究植物的抗病性具有重要的現實意義。
PR1基因是PR基因家族的重要組成部分,它的表達經常被作為系統獲得性抗性(systemic acquired resistance,SAR)建立的標志。最早發現的水稻PR1家族成員OsPR1a和OsPR1b被報道受稻瘟病菌及環境脅迫和一些化學物質的廣泛誘導[6],但有關它們在水稻各個組織中的具體表達模式和誘導特性的信息仍然非常有限。啟動子的起始往往間接反映基因的表達情況,因此,我們通過構建OsPR1bp::GUS表達載體和Real-time PCR,分析了OsPR1b的活性和表達特征,旨在進一步探索水稻SAR 誘導過程和OsPR1b基因表達的內在關系。
1 材料與方法
1.1 材料
水稻品種日本晴(O.sativaL.ssp.japonicacv.Nipponbare)和石狩白毛(O.sativaL.ssp.japonicacv.Ishikari-shiroge,I-S)由本實驗室提供;大腸桿菌DH5α、根瘤農桿菌EHA105和水稻白葉枯菌(Xanthomonas oryzaepv.oryzae,Xoo)菲律賓生理小種P10(PXO124)均為本實驗室保存;雙元載體Pnos-HPT-T35S-GUS-Tnos 由本實驗室在pCAMBIA0380載體基礎上改造獲得;pMD19-T載體、Pyrobest DNA聚合酶購自TaKaRa 公司;總DNA 提取試劑盒DNeasy Plant Mini Kit 購 自Qiagen 公 司;TRIzol 試劑購自Invitrogen 公司;iScript 逆轉錄試劑盒和Sso-Fast EvaGreen Supermix 試劑盒均購自Bio-Rad 公司;DNA 片段及質粒回收試劑盒購自Promega 公司;BamHⅠ和KpnⅠ內切酶購自NEB 公司;T4DNA 快速連接酶購自Fermentas 公司;X-gluc(5-溴-4-氯-3-吲哚基-β-D-葡萄糖醛酸)購自PhytoTechnology Laboratories 公司;甲基茉莉酸(methyl jasmonate,JA)購自TCI公司;乙酰水楊酸(salicylic acid,SA)購自BBI公司;激動素(kinetin,KT)購自Sigma公司;3-吲哚乙酸(3-indole-acetic acid,IAA)、赤霉素(gibberellic acid,GA)、脫落酸(abscisic acid,ABA)和乙烯利(ethephon,ETH)均購自生工公司。
1.2 啟動子的克隆與載體構建
根 據 NCBI 中的水 稻OsPR1b(LOC_Os01g28450.1)基因組序列,利用Oligo 軟件設計引物,用于擴增基因啟始密碼子ATG 前約2.5 kb 的啟動子序列,并在上、下游引物中分別引入BamHⅠ和KpnⅠ酶切位點,便于和GUS報告基因表達載體連接。引物由上海生工生物工程公司合成,序列見表1。按照DNeasy Plant Mini 試劑盒說明書提取水稻日本晴總DNA,并以此DNA 為模板、OsPR1bp-F/R為引物進行PCR 擴增,體系參照Pyrobest DNA 聚合酶說明書。擴增條件為94℃ 5 min,然后以94℃30 s、58℃30 s、72℃2.5 min 行33 個循環。擴增結束后,加入rTaq DNA 聚合酶,繼續在72℃反應10 min 加“A”,獲得的啟動子片段大小為2593 bp。用T4DNA 連接酶于25℃連接30 min,將回收純化的啟動子片段連入pMD19-T 載體中。PCR 鑒定正確后送鉑尚生物技術(上海)有限公司測序,測序結果與GenBank 中的日本晴序列一致。將連有啟動子片段的pMD19-T 載體用BamHⅠ和KpnⅠ酶切,回收片段后,連入經同樣酶切的GUS表達載體中。
1.3 農桿菌介導的水稻轉化
將構建得到的OsPR1bp::GUS表達載體通過電擊轉入農桿菌EHA105 中,參考Hiei 等[7]的轉基因方法,再將農桿菌轉入石狩白毛成熟胚誘導的愈傷組織中,獲得相應的OsPR1bp::GUS轉基因植株。
1.4 GUS組織化學染色
GUS 組織化學染色法參照Jefferson 等[8]報道的方法進行。轉入OsPR1bp::GUS的愈傷組織經首輪篩選21 d 后,隨機挑選抗性愈傷進行染色,其他植物組織取自T2代轉基因植株。用Nikon SMZ1000立體顯微鏡拍攝,拍攝前將含有葉綠素的樣品置于100%酒精中進行脫色處理。染色液成分包括1 mmol/L X-gluc[溶于0.1 mol/L 磷酸緩沖液(pH7.0)中]、10 mmol/L Na2EDTA、1 mmol/L 鐵氰化鉀、1 mmol/L 亞鐵氰化鉀、20%甲醇和0.5% Triton X-100。
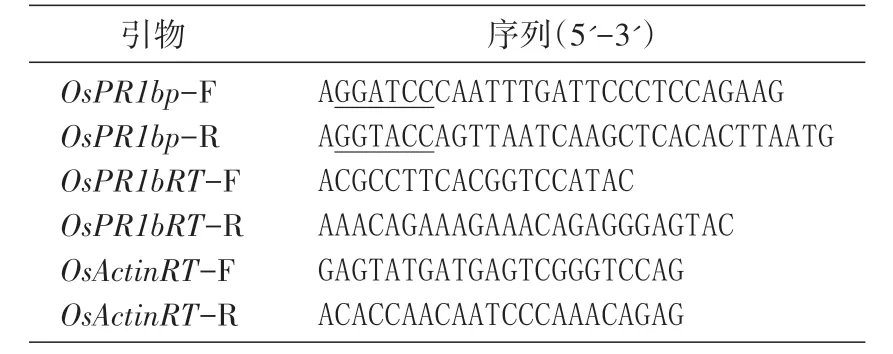
表1 引物及序列
1.5 試驗水稻的處理
水稻石狩白毛種植于浙江省農科院病毒學與生物技術研究所溫室內,采用國際水稻所(IRRI)水稻營養液配方進行培養。水稻胚性愈傷用含2 mg/L 2,4-D 的NB 培養基誘導獲得。取2 周齡根、莖、葉(第四葉),抽穗期帶花幼穗和劍葉,1 月齡胚性愈傷組織進行RNA 提取和定量PCR,分析OsPR1b基因的組織表達特性。
轉基因水稻石狩白毛種子溶液培養于溫室內(生長條件:白天30℃,夜晚25℃,相對濕度60%~80%,光周期12 h/d),待長至四葉一心期時,分別移至含0.1 mmol/L 激素、100 mmol/L NaCl 及10% 聚乙二醇(PEG)的溶液中,以無菌去離子水為對照,處理根部24 h 后分別采集葉片,制備樣品,用于分析OsPR1b基因啟動子的非生物學應答反應。
培養條件同上,取3 周齡水稻幼苗。將培養好的水稻白葉枯菌致病菌株P10(PXO124)調至D600nm值為0.6~1.0,利用剪切接種法接種水稻幼苗,以無菌去離子水剪切為對照,接種24 h 后分別采集葉片,進行樣品制備,用于分析OsPR1b基因啟動子的生物學應答反應。以上所取的每個材料均有3 個不同的生物學重復。
1.6 RNA的提取及逆轉錄反應
樣品用液氮速凍充分研磨后,加入1~2 mL TRIzol,按照TRIzol 試劑盒說明書提取RNA,待沉淀適度干燥后,用20~40μL 無RNase 的水溶解RNA。隨后采用iScript cDNA Synthesis 試劑盒進行逆轉錄反應,合成第一鏈cDNA。
1.7 熒光定量PCR檢測
Real-time PCR 反應體系按SsoFast EvaGreen Supermix 試劑盒說明書配制。擴增條件為95℃ 5 min,以95℃10 s、58℃10 s、72℃20 s 行45 個循環,最后72℃ 10 min。在40℃~95℃溫度范圍內,95℃10 min,40℃10 min,再緩慢提高至95℃,反應結束冷卻至40℃繪制熔解曲線。Real-time PCR所用儀器為LightCycler 480 real-time PCR 儀(Roche公司)。采用比較Ct值的方法進行相對定量,以管家基因OsActin作為內標校正模板量,計算公式為2-ΔΔCt。Real-time PCR 引物見表1。
2 結果
2.1 啟動子的序列分析
以已知的OsPR1b基因(LOC_Os01g28450.1)5'端上游3 kb的區域作為啟動子搜索對象,在PLACE網站(http://www.dna.affrc.go.jp/PLACE/signalscan.html)進行搜索和掃描,對其中潛在的一些順式作用元件進行分析(圖1)。結果表明,OsPR1b啟動子序列除了包含核心啟動元件及CAAT 盒之外[9],還包含W box、TGA element、A box、-624 box、AuxRE 等一些被報道的順式調控元件(表2)。表明OsPR1b基因可能受多重信號分子的調控。
2.2 啟動子表達載體的構建
用BamH Ⅰ和KpnⅠ分別雙酶切Pnos-HPTT35S-GUS-Tnos 載體和重組質粒pMD19-T-Os-PR1bp(圖2B),再經連接、轉化和PCR驗證后獲得相應的啟動子表達分析載體OsPR1bp::GUS(圖2A)。
2.3 轉基因植株的獲得
通過根癌農桿菌介導的遺傳轉化、轉化子篩選和轉基因植株的再生,共獲得了68 個T0代株系,經PCR 分子鑒定及GUS 組織染色分析,得到58 個轉基因陽性株系,其中24 個株系GUS 組織染色呈陽性;隨機挑選部分具有GUS 活性的株系進行繁種,得到T2代株系后進行系統的GUS組織化學分析。
2.4 OsPR1b 基因啟動子在器官或組織中的特異性表達
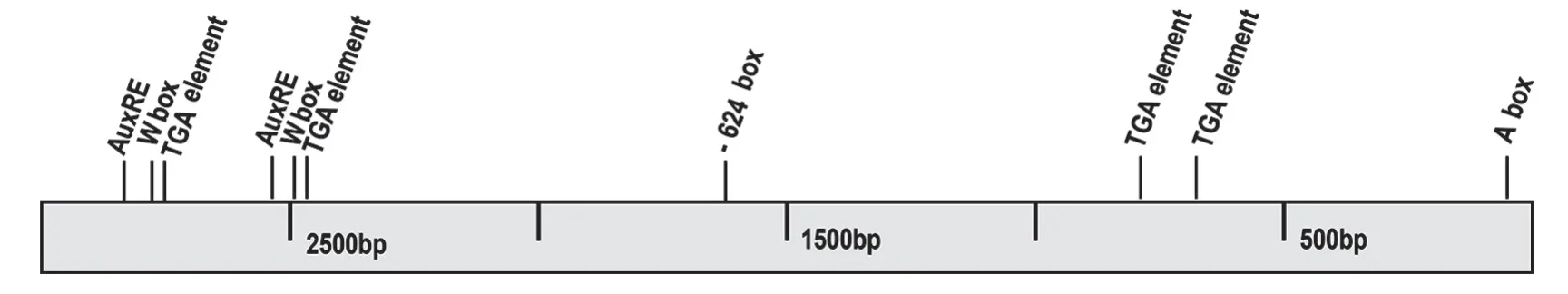
圖1 OsPR1b啟動子中所預測的部分脅迫響應元件位置

表2 OsPR1b啟動子中的順式作用元件詳情
特定的啟動子起始過程常常決定了某個基因是否應當表達或表達量的多少。通過分析OsPR1b啟動子在各個器官或組織中驅動GUS基因的表達情況,可以在一定程度上反映它所控制的OsPR1b基因的時空表達特性。為此,我們對轉OsPR1bp::GUS抗性愈傷組織、T1代轉基因植株的種子及T2代植株的不同部位進行了GUS 組織化學分析,同時,利用Real-time PCR 對野生型愈傷組織和水稻不同時期各組織(14 d 根、莖、葉;開花期劍葉、小花)中OsPR1b基因的表達量進行了分析(圖3)。結果表明,GUS基因在各組織中的表達差異很大,大部分組織均呈現低水平的GUS 活性,葉片中的GUS 染色較為明顯,在5 日齡的幼苗中主要集中在葉尖處,葉緣及節間上也有部分活性(圖3,a~e),在2 周齡的植株葉片及開花期的劍葉中則可以觀察到較為明顯GUS 染色,這些葉片上的染色模式基本上表現為不規則斑點狀分布(圖3,f~i)。另外,在抗性愈傷及萌發的種胚中也存在GUS基因表達的跡象(圖3,l、m)。而在其他部位,如莖、根、花中均未觀察到明顯的GUS活性(圖3,a、j、k)。Real-time PCR 的結果也顯示,OsPR1b在葉片中的表達量遠遠高于其他組織(圖3n)。這些結果證明,OsPR1b基因主要在葉片部位表達,從而發揮其抗病的生物學功能。
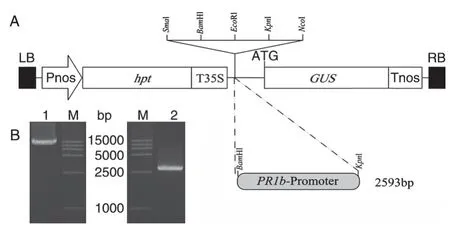
圖2 OsPR1bp::GUS表達載體的構建(A)及質粒酶切電泳圖(B)
2.5 OsPR1bp的誘導表達特性
為了了解所分離的啟動子片段對不同信號分子的響應變化,我們對2 周齡水稻苗的根部進行了各種處理,并利用Real-time PCR 技術對水稻第四葉和根部OsPR1b基因的表達變化進行分析(圖4)。結果顯示,在直接接觸激素的根部,MeJA和KT 可誘導OsPR1b表達增強,誘導倍數分別為17.91±2.27和4.13±2.14;鹽脅迫處理也可顯著提高基因的表達水平,達到了33.09±2.01 倍;其他信號分子,除了H2O2的處理沒有明顯差異外,基因的表達量均出現了不同程度的下調。在水稻的第四葉,KT和ABA可以強烈誘導OsPR1b的表達改變,基因的表達量分別提高了196.8±5.19和62.85±9.12 倍;鹽脅迫和H2O2處理時則分別提高了4.81±1.68和3.00±0.47 倍;其他處理情況下,除了GA 處理抑制外,均出現了不同程度的上調趨勢,其中,抗病信號分子SA和MeJA 的上調變化并不顯著。不論是在根中還是葉片,經過不同激素和非生物脅迫的處理,OsPR1b基因的表達差異變化不大,只有少數幾種激素和鹽脅迫的處理較為顯著,這暗示著OsPR1b基因的表達主要受少數信號途徑的調控。
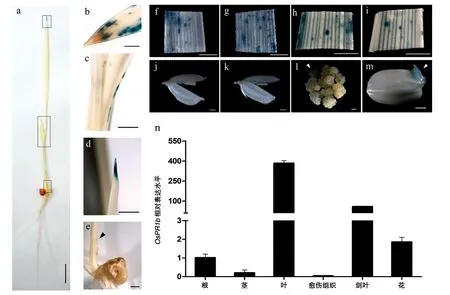
圖3 OsPR1bp::GUS轉基因水稻的GUS染色和OsPR1b基因在健康組織中的表達情況

圖4 激素和非生物脅迫處理24 h后OsPR1b的誘導表達特征
為了闡明水稻致病菌與OsPR1b基因的互作關系,以及抗病信號分子SA和MeJA 在此過程中的作用,我們利用水稻白葉枯菌毒性菌株P10(PXO124)對2 周齡水稻苗第四葉和第五葉進行接種,同時用SA和MeJA 溶液處理其根部。共同處理24 h 后,分別取第四葉和第五葉進行相對表達量的分析(圖5),結果發現,與接水對照相比,單獨接種Xoo后Os-PR1b基因的表達量變化并不顯著;而SA 共同處理組則表現為微弱的上調趨勢,其中第四葉的變化較為明顯;有意思的是,當同時進行接種Xoo和MeJA處理后,OsPR1b基因的表達量受到了明顯的誘導,與對照相比,第四葉和第五葉中分別提高了約17 倍和8倍。這表明MeJA很可能在植物對抗Xoo侵染的過程中起著某種重要的作用。

圖5 水稻白葉枯菌PXO124菌株和SA、MeJA共同處理24 h后OsPR1b基因表達水平的變化
3 討論
作為一種分子標記基因,OsPR1b的表達在健康植物的葉片中幾乎是沉默的[6],在OsPR1bp轉基因水稻中,我們也僅在葉片中觀察到了明顯的GUS活性,而在其他所有組織中的表達十分微弱,甚至不能被檢測到。Real-time PCR 結果也顯示,在同一組織如根中,OsPR1b基因的Ct值和管家基因Actin相比較,有10個循環以上的差異。OsPR1bp在幼葉中的表達主要分布在葉尖和葉緣等一些細胞分裂較為旺盛的部位,這表明OsPR1bp很可能參與了水稻幼苗的生長發育過程。同樣,在萌動的種胚內也出現了GUS活性,事實上,通過進一步的掃描啟動子序列,我們發現在基因的上游存在數個與種子萌發相關的POLASIG1 元件(核心序列為AATAAA)[15]。到了水稻秧苗期和開花期,葉片上的GUS 染色呈現斑點狀的隨機分布,我們推測這種表達模式可能和OsPR1b的抗病性有關。
茉莉酸(JA)和茉莉酸甲酯是脂源性激素分子,調節生長發育的許多方面,調控對非生物及生物脅迫的應答反應。用JA 處理水稻后會誘導許多PR基因如OsPR1b 基因的表達[16]。然而,PR 的誘導表達屬于較晚期的事件[17],我們的實驗表明接種Xoo或JA 單獨處理根部24 h 后,葉片中OsPR1b的表達水平沒有出現明顯的上調變化,但在水稻受到Xoo侵染時,外源施加JA 可以顯著增強OsPR1b基因的表達量,從而增強植株的抗病性,這與已報道的外源施加JA 確實能提高水稻植株抗白葉枯病能力的實驗結果相符[18]。
通常,水稻葉片中存在高水平的自由態SA[19],內源性SA 水平的提高對于水稻防御反應的影響似乎并不顯著[20],PR基因在SA 缺陷的NahG 水稻中的表達也并沒有因為SA 的缺乏而顯著減弱[21],而我們的實驗結果也顯示,在Xoo侵染后24 h,SA 的外源施加也不能引起OsPR1b表達量的顯著增加。然而,SA信號通路的一個重要調節者NPR1,被證明在水稻抗白葉枯病的過程中起重要作用,它能夠和許多TGA家族轉錄因子相互作用,將其在水稻中超表達后能夠極大地提高水稻對Xoo的抗性,同時也激活了一系列防御相關基因[22]。在擬南芥中,TGA3 能夠誘導細胞分裂素(CTK)的轉錄激活因子ARR2 結合于PR1啟動子上,從而激活CTK 響應的防御基因的轉錄[23]。在OsPR1b基因啟動子上我們也發現了數個TGA結合的順式作用元件,Real-time PCR 結果也證明KT 確實能夠顯著增強OsPR1b的表達水平,所以我們推測在水稻中KT 對OsPR1b基因的誘導變化很可能也是通過調節SA-NPR1 的信號路徑,最終影響水稻對病原菌的免疫反應。
值得注意的是,在根中鹽脅迫的誘導非常明顯,2個模擬干旱的非生物因子NaCl和PEG 均能不同程度地提高葉片中OsPR1b的表達水平。ABA 調節著植物生長和發育的許多生理過程,它在水稻對鹽、干旱和寒冷等非生物脅迫的耐受性方面所起的作用已得到廣泛研究[24],近期的研究也表明,干旱條件下水稻植株會產生MeJA,進而刺激產生ABA[25],而我們的研究結果也證明根部施加ABA 確實能夠顯著增強OsPR1b在葉片的表達。因此,我們推斷干旱條件誘導OsPR1b基因的表達很可能是通過刺激水稻植株產生ABA來實現的。
GA 通常被認為是水稻內部免疫反應的負控調節者[26],據報道調節GA 水平的OsGA20ox3轉基因植株可改變水稻對Xoo及稻瘟病的抗性,PR1的誘導表達水平及SA和JA 的積累水平也確實受到了GA水平的影響[27]。這和我們的實驗結果相符,即GA 有可能通過抑制如OsPR1b基因的表達而增加水稻的感病性。
總之,根據我們現階段的實驗結果和前人的研究,我們推測,OsPR1b基因可通過JA、KT和ABA 等激素水平的調控整合到了水稻生物和非生物信號傳導的網絡,從而對Xoo侵染或鹽/干旱脅迫做出響應,同時SA-NPR1 的信號通路可能在其中發揮重要作用。當然,我們也應該認識到,PR基因作為一種較為敏感的基因,它的表達還受到多種因素如生理因素的影響[28]。目前,我們已構建和積累了部分突變體材料,希望通過研究OsPR1b基因在這些突變體中的表達變化,以進一步明確OsPR1b基因在水稻中的表達調控機制。
[1]Gnanamanickam S,Priyadarisini V B,Narayanan N,et al.An overview of bacterial blight disease of rice and strategies for its management[J].Curr Sci,1999,77(11):1435-1444.
[2]Datta S K,Muthukrishnan S.Pathogenesis-related proteins in plants[M].CRC press,1999.
[3]Niki T,Mitsuhara I,Seo S,et al.Antagonistic effect of salicylic acid and jasmonic acid on the expression of pathogenesis-related(PR) protein genes in wounded mature tobacco leaves[J].Plant Cell Physiol,1998,39(5):500-507.
[4]Van Loon L,Pierpoint W,Boller T,et al.Recommendations for naming plant pathogenesis-related proteins[J].Plant Mol Biol Rep,1994,12(3):245-264.
[5]Cao H,Bowling S A,Gordon A S,et al.Characterization of an Arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance[J].Plant Cell Online,1994,6(11):1583-1592.
[6]Agrawal G K,Rakwal R,Jwa N-S,et al.Signalling molecules and blast pathogen attack activates rice OsPR1a and Os-PR1b genes:a model illustrating components participating during defence/stress response[J].Plant Physiol Biochem,2001,39(12):1095-1103.
[7]Hiei Y,Komari T.Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed[J].Nat Protocols,2008,3(5):824-834.
[8]Jefferson R A,Kavanagh T A,Bevan M W.GUS fusions:beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants[J].EMBO J,1987,6(13):3901.
[9]Yamaguchi-Shinozaki K,Shinozaki K.Arabidopsis DNA encoding two desiccation-responsive rd29 genes[J].Plant Physiol,1993,101(3):1119.
[10]Eulgem T,Somssich I E.Networks of WRKY transcription factors in defense signaling[J].Curr Opin Plant Biol,2007,10(4):366-371.
[11]Lebel E,Heifetz P,Thorne L,et al.Functional analysis of regulatory sequences controlling PR? 1 gene expression in Arabidopsis[J].Plant J,1998,16(2):223-233.
[12]Niu X,Renshaw-Gegg L,Miller L,et al.Bipartite determinants of DNA-binding specificity of plant basic leucine zipper proteins[J].Plant Mol Biol,1999,41(1):1-13.
[13]Hennig J,Dewey R E,Cutt J R,et al.Pathogen,salicylic acid and developmental dependent expression of a β-1,3-glucanase/GUS gene fusion in transgenic tobacco plants[J].Plant J,1993,4(3):481-493.
[14]Ulmasov T,Hagen G,Guilfoyle T J.ARF1,a transcription factor that binds to auxin response elements[J].Science,1997,276(5320):1865-1868.
[15]O'Neill S D,Kumagai M H,Majumdar A,et al.The α-amylase genes in Oryza sativa:characterization of cDNA clones and mRNA expression during seed germination[J].Mol Gen Genet,1990,221(2):235-244.
[16]Browse J.Jasmonate passes muster:a receptor and targets for the defense hormone[J].Ann Rev Plant Biol,2009,60:183-205.
[17]Ku J.Concepts and direction of induced systemic resistance in plants and its application[J].Eur J Plant Pathol,2001,107(1):7-12.
[18]Mei C,Qi M,Sheng G,et al.Inducible overexpression of a rice allene oxide synthase gene increases the endogenous jasmonic acid level,PR gene expression,and host resistance to fungal infection[J].Mol Plant Microbe In,2006,19(10):1127-1137.
[19]Silverman P,Seskar M,Kanter D,et al.Salicylic acid in rice(biosynthesis,conjugation,and possible role)[J].Plant Physiol,1995,108(2):633-639.
[20]Yuan Y,Zhong S,Li Q,et al.Functional analysis of rice NPR1 like genes reveals that OsNPR1/NH1 is the rice orthologue conferring disease resistance with enhanced herbivore susceptibility[J].Plant Biotechnol J,2007,5(2):313-324.
[21]Yang Y,Qi M,Mei C.Endogenous salicylic acid protects rice plants from oxidative damage caused by aging as well as biotic and abiotic stress[J].Plant J,2004,40(6):909-919.
[22]Chern M,Fitzgerald H A,Canlas P E,et al.Overexpression of a rice NPR1 homolog leads to constitutive activation of defense response and hypersensitivity to light[J].Mol Plant Microbe In,2005,18(6):511-520.
[23]Choi J,Huh S U,Kojima M,et al.The cytokinin-activated transcription factor ARR2 promotes plant immunity via TGA3/NPR1-dependent salicylic acid signaling in Arabidopsis[J].Dev Cell,2010,19(2):284-295.
[24]Cutler S R,Rodriguez P L,Finkelstein R R,et al.Abscisic acid:emergence of a core signaling network[J].Ann Rev Plant Biol,2010,61:651-679.
[25]Kim E H,Kim Y S,Park S-H,et al.Methyl jasmonate reduces grain yield by mediating stress signals to alter spikelet development in rice[J].Plant Physiol,2009,149(4):1751-1760.
[26]Yang D L,Yang Y,He Z.Roles of plant hormones and their interplay in rice immunity[J].Mol Plant,2013,6(3):675-685.
[27]Yang D-L,Li Q,Deng Y-W,et al.Altered disease development in the eui mutants and Eui overexpressors indicates that gibberellins negatively regulate rice basal disease resistance[J].Mol Plant,2008,1(3):528-537.
[28]Davoine C,Le Deunff E,Ledger N,et al.Specific and constitutive expression of oxalate oxidase during the ageing of leaf sheaths of ryegrass stubble[J].Plant Cell Environ,2001,24(10):1033-1043.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
