PES1蛋白與雄激素受體的相互作用及定位
2014-10-27 09:04:42關鑫王洪遠任偉杜佩云秘曉林程龍葉棋濃
生物技術通訊 2014年3期
關鑫,王洪遠,任偉,杜佩云,秘曉林,程龍,葉棋濃
1.軍事醫學科學院 生物工程研究所,北京 100850;2.大連醫科大學 腫瘤干細胞研究院,遼寧 大連 116000;3.安徽醫科大學 附屬安慶市立醫院口腔科,安徽 安慶 246600;4.福建農林大學 生命科學學院,福建 福州 350002
雄激素受體(androgen receptor,AR)屬于核受體超家族,是一種配體依賴的轉錄因子[1],包含4個結構域,即N端的調控結構域(NTD)、DNA結合結構域(DBD)、鉸鏈區(H)及配體結合結構域(LBD)[2]。NTD結構域又稱為轉錄激活域,它包含一個重要的激活區域 AF1(activation function 1),當人工刪減LBD區域后,AF1可通過配體不依賴的方式發揮作用。LBD結構域包含一個配體依賴的激活區域AF2,其突變或缺失會明顯降低AR的轉錄活性[3]。當AR與雄激素結合后,會發生構象變化而與熱激蛋白HSP90分離。此時AR將以二聚體的形式入核,并與基因中的特定序列——雄激素應答元件(andro?gen response element,ARE)結合,進而激活靶基因的轉錄[4]。
AR在大多數組織中均有不同程度的表達。無論在正常發育中還是病理狀態下,AR都發揮重要的作用。目前,AR已成為前列腺癌的治療靶標[5]。此外,有研究表明AR在很多乳腺癌患者中高表達,且可能成為治療ER-/HER2+乳腺癌患者的新靶標[6-9]。
人PES1蛋白由588個氨基酸殘基(aa)組成,除包含多個核定位信號和一個小泛素化樣修飾蛋白(small ubiquitin-like modifier,SUMO)位點外,在311~415 aa區域存在典型的 BRCT 結構域[10-12]。PES1主要定位于核中,具有DNA結合能力。低等真核生物、鼠或人PES1在正常胚胎發育、核糖體生物發生、DNA復制、染色體穩定性和細胞周期調控中起重要作用[13-16]。
PES1在許多腫瘤,如乳腺癌、膠質瘤、前列腺癌及胃癌等中表達水平上調。我們發現,在雌激素受體(estrogen receptor,ER)陽性的乳腺癌中,PES1通過調節ERα和ERβ的平衡來影響乳腺癌的發生和發展[17]。也有研究表明,在ER陰性的乳腺癌細胞中,PES1仍能促進細胞生長,發揮癌基因的作用,提示在乳腺癌中,PES1還有不依賴于ER的作用方式[18],但這種作用方式尚不明確。AR和ER有著相似的結構,AR能夠調節ERα陽性和ERα陰性乳腺癌細胞的生長,因此我們檢測了PES1是否能夠與AR相互作用,調節AR的表達水平。我們在實驗中發現,PES1能夠與AR相互作用,PES1不調節AR的表達水平,這為研究PES1對乳腺癌中AR功能的調節提供了重要線索。
1 材料與方法
1.1 材料
人胚腎293T細胞由本實驗室保存;乳腺癌ZR75-1敲低PES1穩定克隆細胞系由本實驗室構建;大腸桿菌DH5α感受態細胞購自Invitrogen公司;DMEM培養基和胎牛血清購自GIBCO公司;轉染試劑LipofectAMINE 2000購自Invitrogen公司;質粒Flag-PES1、Flag-PES1(1-110)、Flag-PES1(111-220)、Flag-PES1(221-322)、Flag-PES1(311-588)、Flag-PES1(415-588)均由本實驗室構建;質粒MYC-AR、MYC-AR(1-568)、MYC-AR(484-651)、MYC-AR(484-918)由周建光研究員實驗室贈送;Flag-HRP、MYC-HRP抗體購自Sigma公司;兔抗人GAPDH抗體和辣根過氧化物酶偶聯的IgG均購自Santa Cruz公司。
1.2 細胞培養及轉染
293T細胞和ZR75-1細胞用含10%胎牛血清、50 U/mL青霉素、100 U/mL鏈霉素的DMEM培養基,在CO2含量為5%的37℃孵箱中培養,細胞長滿時用胰酶消化傳代。質粒轉染時嚴格按照說明書中轉染試劑與質粒的量進行操作,轉染4~6 h后更換新鮮的培養基,轉染24 h后收細胞。
1.3 Western印跡及免疫共沉淀
1.3.1 Western印跡 收細胞后加入SDS加樣緩沖液,煮沸15 min,離心后取上清液進行SDS-PAGE,電轉移至硝酸纖維素膜,用5%脫脂奶粉于4℃封閉過夜,加入用5%脫脂奶粉1∶5000稀釋的MYC抗體或1∶10 000稀釋的GAPDH抗體,室溫輕搖1 h,用TBST洗膜3次,每次7 min,加入用5%脫脂奶粉稀釋的辣根過氧化物酶偶聯的羊抗兔IgG,室溫輕搖1 h,用TBST洗膜3次,每次7 min,用化學發光法顯色5 min,壓片顯影。
1.3.2 免疫共沉淀 收集細胞后加入裂解液于冰上裂解30 min,用注射器反復吹打至染色質斷裂,離心去上清,加入偶聯抗體的瓊脂糖珠,4℃結合4 h至過夜,用裂解液洗滌沉淀物3次,每次10 min,在洗滌后的瓊脂糖珠中加入SDS上樣緩沖液進行Western印跡分析,用另一種抗體檢測2種蛋白質之間是否存在相互作用。
2 結果
2.1 PES1蛋白與AR的相互作用
將帶有FLAG標簽的PES1表達質粒和帶有MYC標簽的AR表達質粒共轉染293T細胞,24 h后收細胞,用抗FLAG瓊脂糖珠進行免疫共沉淀,用FLAG和MYC抗體進行Western印跡檢測。結果表明,帶有MYC標簽的AR只在含有FLAG PES1的免疫共沉淀產物中被檢測出來(圖1),即AR能夠特異地與PES1蛋白結合。
2.2 PES1與AR的相互作用定位
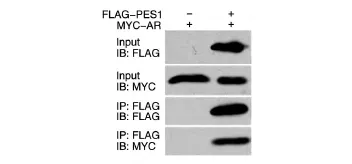
圖1 免疫共沉淀檢測PES1與AR的相互作用
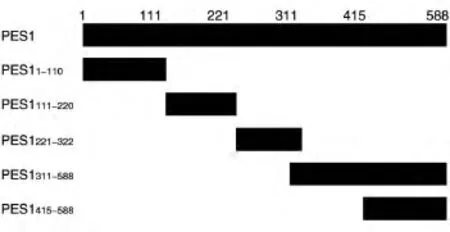
圖2 帶FLAG標簽的PES1截短突變體示意圖
為了確定PES1與AR相互作用的區域,首先構建了帶有FLAG標簽的PES1不同區域的表達載體(圖2),將這些載體和MYC-AR質粒共轉染293T細胞,用上述方法進行免疫共沉淀之后用Western印跡檢測。結果表明,PES11-110、PES1111-220、PES1221-322、和PES1311-588均能與AR相互結合,而PES1415-588不能結合(圖3)。此外,將FLAG-PES1質粒與帶MYC標簽的AR不同區域的表達質粒共轉染293T細胞進行免疫共沉淀后發現,PES1與AR484-918能夠結合,而與AR1-568、AR484-651不結合(圖 4),說明 PES1結合在AR651-918區域。
2.3 PES1不調節AR水平
我們以前的研究發現PES1能和ER結合,并調節ER的蛋白表達水平。因此,我們檢測了PES1是否能夠調節AR的蛋白表達水平。在ZR75-1細胞中轉染PES1 siRNA,48 h后收集細胞,用Western印跡檢測PES1和AR的表達水平,發現敲低PES1后AR的蛋白水平沒有發生變化(圖5)。
3 討論
在本研究中,我們發現PES1能夠與AR相互作用,PES1的多個區域能與AR結合,且主要結合在AR的AF2結構域,PES1不能調節AR的表達水平。
PES1的截短突變體中,除PES1415-588外的區域均能與AR結合,提示PES11-110、PES1111-220、PES1221-322和PES1311-588內均含有與AR直接或間接結合的區域。PES1能夠與AR的AF2區域結合,而AF2區域是配體依賴的轉錄激活域,提示PES1可能作為AR的共調節因子來調節AR的轉錄活性和功能,這需要進一步通過實驗驗證。我們以前的研究發現PES1能與ERα、ERβ相互作用,通過抑制泛素化連接酶CHIP與ERα的結合升高ERα的表達水平,通過促進CHIP與ERβ的結合下調ERβ的表達水平。CHIP也是AR的泛素化連接酶,能夠介導AR通過泛素-蛋白酶體途徑降解。因此,我們檢測了PES1是否能夠調節AR的表達水平,證實PES1不能調節AR的蛋白表達水平,提示PES1可能不能調節CHIP介導的AR蛋白降解。
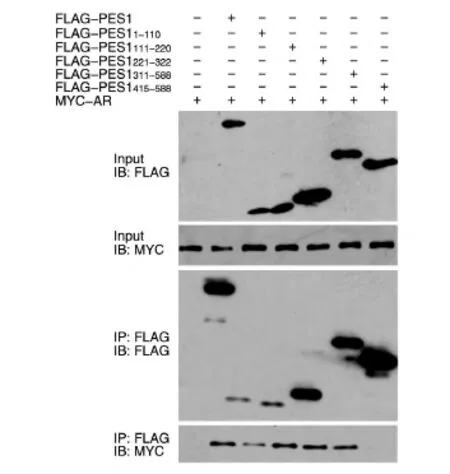
圖3 免疫共沉淀檢測PES1突變體與AR的相互作用
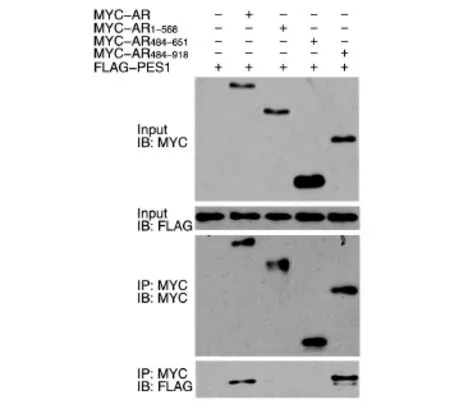
圖4 免疫共沉淀檢測PES1與AR突變體的相互作用
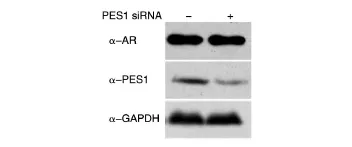
圖5 Western印跡檢測ZR75-1細胞中PES1對AR的影響
乳腺癌發生發展的分子機制復雜,眾多調節因子參與了這一過程。有研究表明,AR在ERα陽性細胞中通過抑制ERα的活性而抑制細胞生長,在ERα陰性細胞中則促進細胞生長。目前關于AR在乳腺癌中的功能及其調節因子和機制的研究較少,本實驗發現了一個新的AR相互作用蛋白PES1,PES1可能通過調節AR的活性調節乳腺癌細胞的增殖。
[1]Mangelsdorf D J,Thummel C,Beato M,et al.The nuclear re?ceptor superfamily:the second decade[J].Cell,1995,83(6):835-839.
[2]Brinkmann A O,Faber P W,Van Rooij H C,et al.The hu?man androgen receptor:domain structure,genomic organiza?tion and regulation of expression[J].J Steroid Biochem,1989,34(1-6):307-310.
[3]Bevan C L,Hoare S,Claessens F,et al.The AF1 and AF2 domains of the androgen receptor interact with distinct re?gions of SRC1[J].Mol Cell Biol,1999,19(12):8383-8392.
[4]Dehm S M,Tindall D J.Molecular regulation of androgen ac?tion in prostate cancer[J].J Cell Biochem,2006,99(2):333-344.
[5]Keren I K,Susan H,Alex G,et al.A genome-wide RNA in?terference screen identifies new regulators of androgen recep?tor function in prostate cancer cells[J].Genome Res,2013,23(4):581-591.
[6]Moe R E,Anderson B O.Androgens and androgen receptors:aclinically neglected sectorin breastcancerbiology[J].J Surg Oncol,2007,95(6):437-439.
[7]Higgins M J,Wolff A C.The androgen receptor in breast can?cer:learning from the past[J].Breast Cancer Res Treat,2010,124(3):619-621.
[8]Park S,Koo J S,Kim M S,et al.Androgen receptor expres?sion is significantly associated with better outcomes in estro?gen receptor-positive breast cancers[J].Ann Oncol,2011,22(8):1755-1762.
[9]Ni M,Chen Y,Lim E,et al.Targeting androgen receptor in estrogen receptor-negative breast cancer[J].Cancer Cell,2011,20(1):119-131.
[10]Du Y C,Stillman B.Yphlp,an ORC-interacting protein:po?tential links between cell proliferation control,DNA replica?tion,and ribosome biogenesis[J].Cell,2002,109(7):835-848.
[11]Holzel M,Grimm T,Rohrmoser M,et al.The BRCT domain of mammalian PES1 is crucial for nucleolar localization and rRNA processing[J].Nucleic Acids Res,2007,35(3):789-800.
[12]Kinoshita Y,Jarell A D,Flaman J M,et al.Pescadillo,a novel cell cycleregulatory protein abnormally expressed in ma?lignant cells[J].J Biol Chem,2001,276(9):6656-6665.
[13]Lerch-Gaggl A,Haque J,Li J X,et al.Pescadillo is essen?tial for nucleolar assembly,ribosome biogenesis,and mammali?an cell proli-feration[J].J Biol Chem,2002,277(47):45347-45355.
[14]Prisco M,Maiorana A,Guerzoni C,et al.Role of pescadillo and upstream binding factor in the proliferation and differenti?ation of murine myeloid Cells[J].Mol Cell Biol,2004,24(12):5421-5433.
[15]HolzelM M,RohrmoserM,SchleeT,etal.Mammalian WDR12 is a novel member of the Pes1-Bop1 complex and is required for ribosome biogenesis and cell proliferation[J].J Cell Biol,2005,170(3):367-378.
[16]Zhang H,Fang Y,Huang C,et al.Human pescadillo induces large-scale chromatin unfolding[J].Sci China C Life Sci,2005,48(3):270-276.
[17]Cheng L,Li J P,et al.PES1 promotes breast cancer by dif?ferentially regulating ERα and ERβ[J].J Clin Invest,2012,122(8):2857-2870.
[18]Li J,Yu L,Zhang H Z,et al.Down-regulation of pescadillo inhibits proliferation and tumorigenicity of breast cancer cells[J].Cancer Sci,2009,100(12):2255-2260.
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中國生殖健康(2019年2期)2019-08-23 08:11:42
人大建設(2019年12期)2019-05-21 02:55:32
中國生殖健康(2019年6期)2019-01-06 09:20:12
祝您健康(2018年5期)2018-05-16 17:10:16
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
電測與儀表(2015年5期)2015-04-09 11:30:52
民生周刊(2012年10期)2012-10-14 09:06:46
