嗜熱芽孢桿菌噬菌體的分離及特征研究
2013-10-29 09:36:10晏愛芬余麗林連兵
生物技術通訊 2013年1期
晏愛芬,余麗,林連兵
1.保山學院 資源環境學院,云南 保山 678000;2.昆明理工大學 生物工程技術研究中心,云南 昆明 650224
噬菌體是一類寄生在古生菌和真細菌中的病毒,是體積微小、結構簡單、性質特殊的生命形態。隨著對噬菌體在生物學和醫學領域中地位的認識不斷加深,高溫噬菌體引起了越來越多科研工作者的關注,因為它們不僅可以作為一種模式系統,用于研究地熱區生物的分子生物學及生物化學特性,而且還深刻影響著生物地球化學和生態系統的進化過程,包括遺傳轉換、營養循環、生物種群結構、生物分類、生物進化[1]等。對騰沖熱海嗜熱芽孢桿菌噬菌體的研究,不僅可以了解它們在騰沖熱海中的分布及特征,還有助于豐富我國嗜熱噬菌體資源。
本文報道了分離自騰沖熱海熱泉的一株嗜熱芽孢桿菌噬菌體,我們對其進行了電鏡形態觀察、生理特征初步研究。
1 材料和方法
1.1 材料
噬菌體分離所用的宿主菌嗜熱芽孢桿菌為本實驗室保存;從云南省騰沖縣熱海熱泉采集水樣及沉積物,室溫保存于無菌的離心管中,采樣點溫度為50~75℃,pH值為7.0~8.0。
高溫菌培養基DSM88[2],調節pH值為7.5;固體培養基的瓊脂濃度為2%,半固體培養基的瓊脂濃度為1%。
1.2 噬菌體的分離及純化
采用雙層平板法分離嗜熱噬菌體[3]。取20 mL熱泉樣品接種到200 mL DSM88液體培養基中,55℃、200 r/min振蕩過夜,所得培養液經0.22 μm滅菌濾膜過濾除菌后,取100 μL濾液與350 μL培養至對數生長期早期的嗜熱芽孢桿菌混合,室溫靜置10 min后與3 mL半固體培養基混勻,均勻傾倒在DSM88固體平板上,即形成雙層平板;將雙層平板置于密封袋中,55℃培養過夜;肉眼觀察到噬菌斑后,用牙簽挑取單個噬菌斑,溶于DSM88液體培養基中,作為下次感染的病毒儲液感染宿主細胞,經3次純化獲得純噬菌體。
1.3 噬菌體的形態觀察
取純化后的噬菌體(效價為108pfu/mL)10 μL,用一定量的2%醋酸雙氧鈾染色,吸附于230目銅網(噴涂碳膜)上,干燥后用JEOL-TEM1400透射電子顯微鏡觀察。
1.4 最佳感染復數的測定
感染復數(multiplicity of infection,MOI)是指初始感染時加入噬菌體的數量與宿主菌數量的比值,也稱感染倍數。參照Lu等[4]的方法,略有改動。培養宿主菌NHH4至對數前期,用分光光度計測定此時的 D600nm值約為 0.2,相當于 1×108CFU/mL;按照MOI分別為0.01、0.1、1、10和100的比例,加入噬菌體純培養液和宿主菌,加入DSM88培養基使各管總體積相同;在55℃搖床中160 r/min振蕩培養8 h,13 000×g離心10 min,收集上清測定噬菌體滴度;各點均作2份復管培養,取平均值,同時以不加噬菌體的宿主菌和不加宿主菌的噬菌體為對照,以產生最高噬菌體滴度的MOI為最佳感染復數。
1.5 物理因素對噬菌體活性的影響
1.5.1 噬菌體的熱穩定性分析[5]取滴度為2.1×108pfu/mL的噬菌體懸液1 mL,分別置于50、55、60、65℃水浴鍋中,每隔10 min取一次樣,共取5次,測定各溫度梯度下不同時間的噬菌體滴度 。
1.5.2 噬菌體pH穩定性分析[5]取pH值分別為4、5、6、7、7.5、8、9、10、11的DSM88培養基0.99 mL,加入1.5 mL滅菌離心管中,加入0.01 mL滴度為2.1×108pfu/mL的噬菌體懸液,室溫靜置1 h,用DSM88培養基稀釋至1/10,取20 μL與350 μL對數期宿主細胞混合,室溫靜置15 min后測定滴度。
2 結果
2.1 噬菌體的分離和純化
把不同采樣點的富集樣品過濾除菌得到的病毒儲液,與培養至對數期的不同菌株吸附,采用雙層平板法檢測是否有噬菌斑形成。結果NHH4菌株的菌苔可見清新、透明的噬菌斑,直徑2~4 mm(圖1)。反復挑取3次純化噬菌體,在合適的液體培養條件下分離到1株噬菌體能快速裂解宿主菌,使培養液變得澄清,噬菌體滴度較高,一般可達108pfu/mL以上;電鏡觀察,可見形態均一的噬菌體顆粒。
2.2 噬菌體電鏡形態
用滅菌牙簽把雙層平板上的噬菌斑挑到液體培養基中培養24 h,對噬菌體培養液超速離心濃縮,于電鏡下觀察,噬菌體顆粒為二十面體型,直徑為80~100 nm,無尾,未見刺突,衣殼較薄,截面呈六邊形,邊長60~80 nm(圖2)。根據其形態,將感染嗜熱芽孢桿菌NHH4的噬菌體命名為嗜熱芽孢桿菌二十面體噬菌體(thermophilicBacillusicosahedral phage,TBIP1)。
2.3 噬菌體TBIP1的最佳MOI測定
加入噬菌體和宿主菌培養8 h后,宿主菌被充分裂解,培養液變得澄清,計數各測定管中的噬菌體滴度,結果見表1。當MOI為1、0.1和0.01時,噬菌體TBIP1感染其宿主菌NHH4產生的子代噬菌體的滴度較高;當MOI為1.0時,子代噬菌體滴度為1.09×109PFU/mL,在所有MOI中,該產出率最高。因此,確定噬菌體TBIP1感染其宿主嗜熱芽孢桿菌NHH4的最佳MOI為1.0。

圖1 噬菌體在平板上形成的噬菌斑形態

圖2 二十面體噬菌體TBIP1的形態
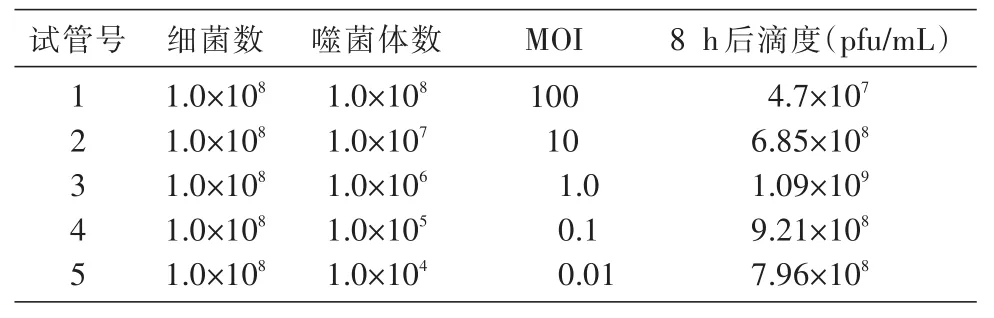
表1 噬菌體TBIP1的感染復數測定
2.4 物理因素對噬菌體TBIP1活性的影響
2.4.1 溫度對噬菌體活性的影響 宿主菌NHH4在45~65℃可被TBIP1感染,以培養溫度為55℃時獲得的噬菌體滴度最高,培養液中噬菌體滴度可達2.8×108pfu/mL。熱穩定性分析(圖3)表明,TBIP1在55℃最穩定,隨著溫度升高,其存活率降低,65℃處理20 min后噬菌體失活。
2.4.2 pH值對噬菌體活性的影響 pH值的改變對噬菌體TBIP1滴度的影響較大,從圖4可以看出,在pH為5~9時,噬菌體均保持45%以上的活性,尤以pH7.5時滴度最高;隨著pH值的升高,噬菌體TBIP1的活性快速降低,pH11時噬菌體完全失去感染宿主的能力;當pH值降到4時滴度為零。相比之下,TBIP1在較為溫和的pH值環境下有較好的穩定性。
3 討論
高溫噬菌體生活在一類特殊的環境中,對溫度具有很強的耐受力和適應性,它們通過侵染嗜熱細菌而生存繁殖,具有獨特的分子生物學和生物化學特性。噬菌體TBIP1是分離自從騰沖熱海溫泉中得到的嗜熱芽孢桿菌NHH4的一株新的烈性噬菌體。TBIP1在DSM88培養基中能夠很好地感染宿主菌NHH4并形成清晰的噬菌斑,感染宿主菌的溫度范圍為45~65℃,最適感染溫度為55℃;感染宿主菌的pH值范圍為5.0~10.0,最適感染pH值為7.5。根據對噬菌體TBIP1的生物學特征分析,認為噬菌體TBIP1比較典型。
騰沖熱海作為我國著名的火山地熱區,具有不同水化學特征的溫泉類型,與之對應的漫長的微生物生態系統的演化,孕育出了現代騰沖熱海豐富的高溫微生物資源。已從騰沖熱海中分離到硫化葉菌噬菌體[6]、棲熱菌噬菌體[5],可見騰沖熱海的嗜熱菌及其噬菌體都具有生物多樣性。騰沖熱海作為我國重要的高溫菌及其嗜熱噬菌體資源寶庫,值得進一步研究。
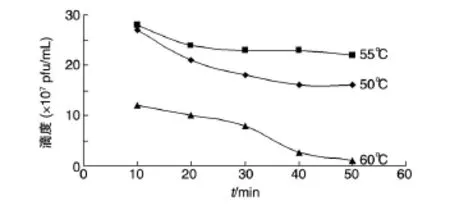
圖3 溫度對噬菌體TBIP1活性的影響
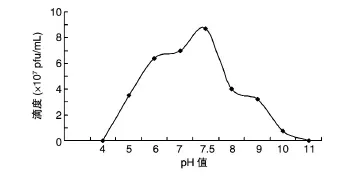
圖4 不同pH值對噬菌體TBIP1活性的影響
[1]Fuhrman J A.Marine viruses and their biogeochemical and ecological effects[J].Natue,1999,399:541-548.
[2]Chen C,Lin L,Qian P,et al.Meiothermus rosaceus sp.nov.isolated from Tengchong hotspring in Yunnan,China[J].FEMS Microbiol Lett,2002,216:263-268.
[3]Martha R J,Clokie A M K.Bacteriophages:methods and pro?tocols,Volume 1:Isolation,characterization,and interactions[M].Humana Press,2008:28.
[4]Lu Z,Breidt F Jr,Fleming H P,et al.Isolation and charac?terization of a Lactobacillus plantarum bacteriophage,ΦJL21,from a cucumber fermentation[J].Int J Food Microbiol,2003,84(2):225-235.
[5]洪偉,韓劍,戴欣,等.騰沖熱海一株棲熱菌裂解性噬菌體的分離及其特征[J].微生物學報,2010,50(3):322-327.
[6]Xiang X,Chen L,Huang X,et al.Sulfolobus tenchongensis spindle-shaped rius STSV1:virus-host interactions and genom?ic features[J].Virology,2005,79:8677-8686.
