c-Fos不同結構域的真核表達及功能鑒定
2013-10-29 09:36:08付潔王瑜程龍徐小潔張浩宋海峰葉棋濃
生物技術通訊 2013年1期
付潔 ,王瑜 ,程龍 ,徐小潔 ,張浩 ,宋海峰 ,葉棋濃
軍事醫學科學院 a.生物工程研究所;b.放射與輻射醫學研究所;北京 100850
激活蛋白1(activator protein 1,AP-1)是一類由即刻早期應答基因編碼的核轉錄因子,在哺乳動物中,AP-1主要由 Fos家族(c-Fos、FosB、Fra-1和Fra-2)和Jun家族(c-Jun、JunB和JunD)組成。AP-1蛋白以同源(Jun-Jun)或異源二聚體(Fos-Jun)的形式發揮轉錄激活作用。基礎條件下,細胞中的AP-1以Jun同源二聚體為主,其表達水平和活性均很低;當細胞受到生理和病理因素刺激時,如細胞因子、生長因子、應激信號、感染、致癌劑等的刺激,細胞中的AP-1將以c-Fos和c-Jun異源二聚體為主,其表達水平和活性均迅速升高,激活下游信號轉導通路,在多種腫瘤的生長、侵襲和轉移中發揮重要作用[1-2]。AP-1作為對外界信號發生反應的轉錄因子而且位于信號級聯反應的末端,使外界信號和細胞功能之間建立起一定的聯系,因此已作為抗腫瘤藥物開發的重要靶標之一。
調節AP-1活性的方式目前主要有以下幾個層面:二聚體的組成成分和量的不同,會在腫瘤發生發展的不同階段起到致癌或抑癌的效果[3-4];轉錄水平或翻譯后水平的磷酸化修飾等調節方式[5-6];蛋白質間相互作用,如與癌蛋白(RAS)或輔助蛋白(糖皮質激素受體GR或維甲酸受體RAR)等的相互作用也是調節AP-1轉錄活性的重要方式之一[7]。盡管已發現一些與AP-1相互作用的蛋白質,但只能部分解釋腫瘤發生發展機制,且許多與AP-1相互作用的蛋白質的臨床意義不清楚。由于AP-1在腫瘤發生發展中起極其重要的作用,因此與AP-1相互作用的新因子還在不斷被發現。哺乳動物Fos家族中以c-Fos最重要,Fos家族在調節AP-1活性和腫瘤發生發展過程等方面具有重要作用[8-10]。
我們擬構建c-Fos蛋白的不同結構域片段的真核表達載體,并在293T細胞中表達,利用免疫共沉淀技術確定構建的結構域的正確性,為進一步研究與c-Fos相互作用的蛋白及其區域定位奠定基礎。
1 材料與方法
1.1 材料
人HEK293T細胞株由本實驗室保存(用含10%熱滅活小牛血清的DMEM培養基,培養于37℃、5%CO2細胞培養箱);大腸桿菌DH5α、FLAG-pcDNA3.0載體由本實驗室保存;LipofectAMINE2000轉染試劑、DMEM培養基購自Invitrogen公司;小牛血清購自杭州四季青公司;限制性內切酶、DNA連接酶、PrimeSTAR HS DNA聚合酶購自TaKaRa公司;質粒提取試劑盒、凝膠回收試劑盒、PCR回收試劑盒、顯色液均購自康維世紀公司;預染蛋白marker購自Fermentas公司;瓊脂糖珠偶聯的抗FLAG抗體購自Sigma公司;抗FLAG和抗HA標簽抗體、蛋白酶抑制劑cocktail均購自Sigma公司;擴增基因片段的PCR引物由上海生工生物有限公司合成。
1.2 c-Fos基因結構域劃分及引物設計
以本實驗室保存的FLAG-c-Fos(GenBank登錄號為NM_005252)質粒為模板,根據文獻[11]將c-Fos全長分為c-FosB、c-FosD和c-FosE等3個結構域片段(圖1),用PCR技術設計引物(表1),將c-Fos分為3段結構域克隆到FLAG-pcDNA3.0載體中,酶切位點采用BamHⅠ和EcoRⅠ。因為構建的表達載體在3'端含有FLAG標簽編碼序列,因此將各結構域擴增片段后的終止密碼子在引物中去掉,使目的蛋白與標簽融合表達。
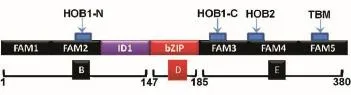
圖1 c-Fos基因結構域劃分模式圖
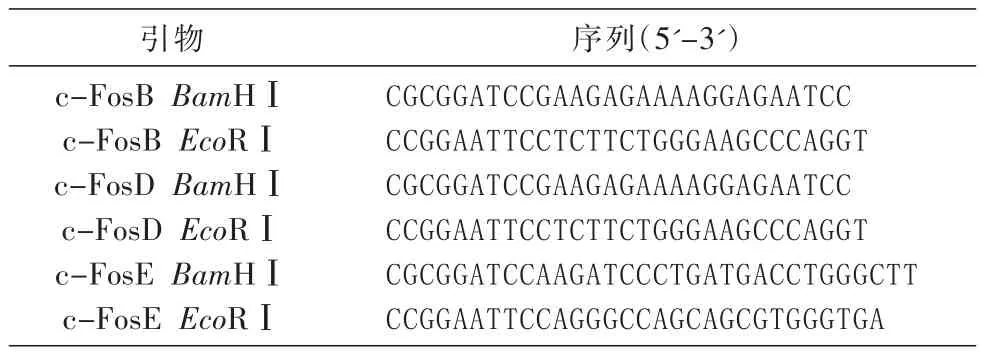
表1 引物及序列
1.3 PCR程序和體系
參照PrimeSTAR HS DNA聚合酶說明書進行。在0.2 mL離心管中依次加入1 μL質粒模板、引物各0.5 μL(儲存濃度 10 μmol/L)、5×PS緩沖液10 μL、dNTP 4 μL、0.5 μL PrimeSTAR 酶 ,用ddH2O將體系補足至50 μL。將上述混合液輕輕混勻,短暫離心。PCR反應程序:95℃ 10 min,然后以95℃ 20 s、55℃ 20 s、72℃ 1 min 進行 30 個循環,72℃ 7 min。取9 μL PCR產物,加入10×上樣緩沖液1 μL,混勻,上樣,進行1.5%瓊脂糖凝膠電泳檢測,紫外凝膠成像儀驗證條帶是否正確。
1.4 FLAG-c-Fos不同結構域片段真核表達載體的構建
瓊脂糖凝膠回收PCR產物,與FLAG-pcDNA3.0載體質粒分別用BamHⅠ/EcoRⅠ雙酶切,將酶切后的目的基因與載體DNA分別用1.5%和1%瓊脂糖凝膠電泳回收,用T4DNA連接酶于16℃連接過夜。連接產物轉化大腸桿菌DH5α,涂板后37℃溫箱中倒置培養至長出克隆菌落。挑取克隆菌落放入5 mL含氨芐西林的LB培養基中,37℃、220 r/min振蕩過夜培養,用質粒小量純化試劑盒少量提取質粒。表達載體質粒經雙酶切鑒定無誤后,將樣品送上海生工公司測序。
1.5 表達載體質粒的提取
將測序結果序列比對后,樣品接入10 mL含有氨芐西林的LB培養基中,37℃、220 r/min振蕩過夜培養,用Promega公司的質粒提取試劑盒提取質粒,紫外分光光度法測定濃度和純度,D260nm/D280nm為1.8~1.9的質粒保存在-20℃待用。
1.6 細胞轉染和收集
293T細胞用含10%胎牛血清、2 mmol/L谷酰胺、1×105U/L青霉素和100 mg/L鏈霉素的DMEM培養液培養。將105細胞鋪24孔板,次日細胞融合率約為70%時用LipofectAMINE2000轉染試劑將1.5 μg FLAG-c-FosB、FLAG-c-FosD 和 FLAG-c-FosE質粒轉染,轉染后24~36 h以1000 r/min離心收集細胞,棄上清,用PBS洗滌一次,置冰上,加入1×SDS上樣緩沖液,充分裂解細胞后行SDS-PAGE,Western印跡檢測片段表達正確與否。
進行免疫共沉淀的轉染實驗,細胞培養條件同上,將106細胞鋪35 mm直徑的小皿,次日細胞融合率約為70%時用LipofectAMINE2000轉染試劑將5 μgHA-c-Jun和 5 μgFLAG-c-FosB、FLAG-c-FosD、FLAG-c-FosE分別共轉染,轉染后24~36 h,以1000 r/min離心收集細胞,棄上清,用PBS洗滌一次,置冰上。
1.7 共轉染蛋白分子的免疫沉淀
用0.5 mL預冷的細胞裂解液[1 mol/L Tris-HCl(pH8.0)2 mL、5 mol/L NaCl 2.5 mL、NP-40 0.25 mL、0.5 mol/L EDTA 1 mL、蛋白酶抑制劑0.1 mL、1 mol/L DTT 0.1 mL,用 ddH2O 補至 100 mL]冰浴30 min后超聲波裂解細胞,4℃、12 000 r/min離心,取適量上清與等體積2×SDS蛋白加樣緩沖液混勻作為input,-70℃保存備用于免疫印跡分析。將其余上清轉移到新的無菌管中,加入瓊脂糖珠偶聯的抗FLAG抗體,于4℃在旋轉混合器上孵育結合4 h后用裂解液洗滌瓊脂糖珠4次,每次4℃、3000 r/min離心3 min,洗滌完畢棄上清,向沉淀中加入30 μL 2×SDS蛋白加樣緩沖液,-70℃保存備用于免疫印跡分析。
1.8 免疫沉淀標本的免疫印跡
將上述收集的免疫沉淀標本煮沸10 min變性,經10%SDS-PAGE分離,采用半干轉印儀,以15 V恒壓轉印50 min,將凝膠中的蛋白質轉移至PVDF膜上,以含30 g/L脫脂奶粉的TTBS[100 mmol/L Tris-HCl(pH7.5)、9 g/L NaCl,0.1%Tween-220]室溫封閉1 h,加1∶5000稀釋的HA-HRP或FLAGHRP抗體室溫孵育1 h,用TTBS洗滌后,用A/B液于X線膠片上曝光檢測。
2 結果
2.1 c-Fos不同結構域的構建
c-Fos不同結構域片段的構建如圖1,PCR擴增后獲得c-FosB、c-FosD和c-FosE片段,分別為441、237、531 bp(圖 2A),經酶切、連接、轉化和挑取菌落,對重組質粒進行雙酶切鑒定(圖2B),將酶切正確的質粒送奧科生物公司測序,證實重組質粒構建正確(數據未示)。
2.2 c-Fos不同結構域的表達
將c-Fos不同結構域片段的表達載體轉染293T細胞后,收集細胞進行Western印跡。c-FosB(147殘基)、c-FosD(79殘基)、c-FosE(176殘基)的理論相對分子質量分別為 18×103、10×103、22×103,因連接有FLAG標簽,表達產物比理論值略大(圖3)。
2.3 HA-c-Jun和FLAG-c-Fos不同結構域片段的免疫共沉淀實驗
免疫共沉淀方法檢測到FLAG瓊脂糖珠能將FLAG-c-FosB、FLAG-c-FosD和FLAG-c-FosE從細胞裂解液中沉降下來,并且在FLAG-c-FosD組可以特異性地檢測到HA-c-Jun的存在(圖4),說明本實驗構建的FLAG-c-FosD可特異性地與HA-c-Jun結合,而FLAG-c-FosB、FLAG-c-FosE不與HA-c-Jun結合,這與文獻報道的c-Fos和c-Jun形成異源二聚體依賴于bZIP區[16-17]相符。
3 討論
蛋白質間相互作用存在于機體每個細胞的生命活動過程中,包括復制、轉錄、翻譯、細胞周期調控和信號轉導等,特定蛋白質相互作用發生改變可以導致疾病的發生,因此,基于信號轉導通路的關鍵蛋白進行與其相互作用蛋白的篩選,是新藥研發的重要內容之一。在AP-1信號途徑中,AP-1與其他蛋白質的相互作用是AP-1發揮功能活性的重要方式之一。這些與AP-1相互作用的蛋白不但能改變AP-1活性,而且其本身基因結構和/或表達水平在病理狀態下也可能發生改變,因此這些相互作用蛋白也是藥物開發的潛在靶標。
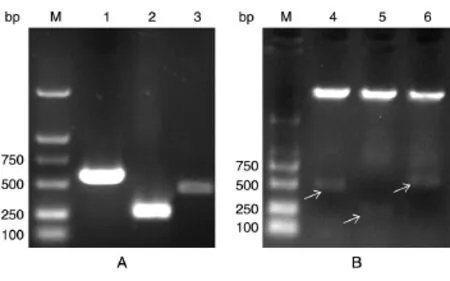
圖2 c-Fos不同片段結構域的構建鑒定
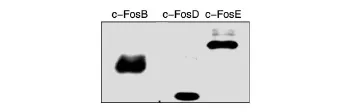
圖3 利用FLAG標簽抗體對c-Fos不同結構域片段進行表達鑒定

圖4 FLAG標簽 c-Fos不同結構域片段與HA-c-Jun的免疫共沉淀
AP-1通過蛋白質間相互作用來發揮其功能至少有以下幾種方式:①通過AP-1不同成員之間的相互作用調節AP-1轉錄活性。細胞中AP-1的組成和各組分的相對比例決定了AP-1的功能[3-4]。②通過AP-1與其他蛋白質的相互作用調節AP-1轉錄活性。如c-Jun或c-Fos可與一些Maf蛋白(v-Maf和c-Maf)相互作用形成異源二聚體;c-Fos還能與MafB、MafF、MafG和MafK相互作用,c-Jun則不能。通過這些相互作用,AP-1的功能大大增強[12-13]。一些調節蛋白也可通過與AP-1的相互作用抑制AP-1的功能。LaminA/C蛋白通過與c-Fos的相互作用改變c-Fos的亞細胞定位、抑制AP-1的DNA結合活性、抑制AP-1靶基因的轉錄激活,并導致細胞周期改變[14]。③AP-1通過與其他轉錄因子的相互作用調節AP-1或其他轉錄因子的轉錄活性[15]。如屬于轉錄因子的雌激素受體(ER)可與AP-1相互作用并調節AP-1轉錄活性[6],AP-1與轉錄因子Smad結合后可調節Smad介導的轉化生長因子β(TGF-β)信號通路活性。因此,構建AP-1家族主要蛋白的不同結構域片段,對于研究其相互作用蛋白并進一步功能定位,具有重要意義。
哺乳動物細胞中,AP-1家族的主要成員為Fos和Jun家族,其中,c-Fos是Fos家族的最重要成員。根據現有文獻報道[16-17],c-Fos有多個結構域可以激活或調節轉錄,其中C端結構域有3個Fos激活分子(Fos activation modlues,FAM),其中2個FAM區含HOB1和HOB2基序,第三個FAM區含TBP結合基序(TBP binding motif,TBM),TBM的TBP結合活性可以被含有HOB1和HOB2的激活結構域放大;C-fos的N端含有一個抑制性結構域(inhibitory do?main,ID1),可作用于含有HOB1基序的FAM區,使之沉默;C-Fos含有堿性亮氨酸拉鏈(bZIP)的DNA結合結構域。我們希望獲得可以和c-Fos相互作用的蛋白,因此并未對c-Fos的N端和C端的結構域進行細分,而只分為3個結構域。本研究中免疫共沉淀結果也表明,bZIP與c-Jun結合,從而佐證了c-Fos不同結構域構建的正確性。
[1]Verde P,Casalino L,Talotta F,et al.Deciphering AP-1 func?tion in tumorigenesis[J].Cell Cycle,2007,6(21):2633-2639.
[2]Malnou C E,Brockly F,Favard C,et al.Heterodimerization with different Jun proteins controls c-Fos intranuclear dynam?ics and distribution[J].J Biol Chem,2010,285(9):6552-6562.
[3]Bakiri L,Matsuo K,Wisniewska M,et al.Promoter specifici?tyand biologicalactivityoftethered AP-1 dimers[J].Mol Cell Biol,2002,22(13):4952-4964.
[4]van Dam H,Castellazzi M.Distinct roles of Jun:Fos and Jun:ATF dimersin oncogenesis[J].Oncogene,2001,20(19):2453-2464.
[5]O'Donnell A,Yang S H,Sharrocks A D.MAP kinase-mediat?ed c-fos regulation relies on a histone acetylation relay switch[J].Mol Cell,2008,29(6):780-785.
[6]Portal M M,Ferrero G O,Caputto B L.N-terminal c-Fos ty?rosine phosphorylation regulates c-Fos/ER association and c-Fos-dependentphospholipid synthesisactivation[J]. Onco?gene,2007,26(24):3551-3558.
[7]Herrlich P.Cross-talk between glucocorticoid receptorand AP-1[J].Oncogene,2001,20(19):2465-2475.
[8]Milde-Langosch K.TheFosfamilyoftranscription factors and their role in tumourigenesis[J].Eur J Cancer,2005,41(16):2449-2461.
[9]Lu C,Shen Q,DuPré E,et al.c-Fos is critical for MCF-7 breast cancer cell growth[J].Oncogene,2005,24(43):6516-6524.
[10]Durchdewald M,Angel P,Hess J.The transcription factor Fos:a Janus-type regulator in health and disease[J].Histol Histopathol,2009,24(11):1451-1461.
[11]John M,Leppik R,Busch SJ,et al.DNA binding of Jun and FosbZip domains:homodimersand heterodimersinducea DNA conformational change in solution[J].Nucleic Acids Res,1996,24(22):4487-4494.
[12]Chinenov Y,Kerppola T K.Close encounters of many kinds:Fos-Jun interactions that mediate transcription regulatory spec?ificity[J].Oncogene,2001,20(19):2438-2452.
[13]Eferl R,Wagner E F.AP-1:a double-edged sword in tumori?genesis[J].Nat Rev Cancer,2003,3(11):859-868.
[14]Ivorra C,Kubicek M,González J M,et al.A mechanism of AP-1 suppression through interaction of c-Fos with lamin A/C[J].Genes Dev,2006,20(3):307-320.
[15]Lopez-Bergami P,Lau E,Ronai Z.Emerging roles of ATF2 and the dynamic AP1 network in cancer[J].Nat Rev Cancer,2010,10(1):65-76.
[16]Brown H J,Sutherland J A,Cook A,et al.An inhibitor do?main in c-Fosregulatesactivation domainscontaining the HOB1 motif[J].EMBO J,1995,14(1):124-31.
[17]Metz R,Bannister A J,Sutherland J A,et al.c-Fos-induced activation of a TATA-Box-containing promoter involves direct contactwith TATA-box-binding protein[J].MolCellBiol,1994,14(9):6021-6029.
