不同封育季節放牧干擾對青海云杉種群結構和動態的影響
2012-08-20 08:01:58劉建泉羅永壽呂海元
草業科學 2012年6期
劉建泉,羅永壽,呂海元
(1.甘肅祁連山國家級自然保護區管理局,甘肅 張掖734000;2.甘肅祁連山國家級自然保護區哈溪保護站,甘肅 武威733000)
林區放牧既是森林資源多種利用的途徑之一,又因其影響幼苗正常生長而引發林牧矛盾,不合理的放牧已成為森林群落自我恢復的重要障礙之一。祁連山區是一個林、牧混雜的區域,傳統的牧業活動引發的生態保護與區域經濟社會發展的矛盾日趨突出,特別是林區尖銳的林牧矛盾已引起社會各界的普遍關注[1-2],并采取了生態移民、封山禁牧、退耕還林、天然林保護、公益林保護、石羊河和黑河流域綜合治理、舍飼養殖等重大的政策、工程和技術措施,保護祁連山生態環境,特別是保護祁連山水源涵養林。青海云杉(Picea crassifolia)是祁連山水源涵養林的主要組成樹種,合理的種群結構是其健康和持續利用的基礎。目前關于自然狀態下青海云杉種群的分布格局、種群數量動態和群落特征及動態、種間關系和聯結相對清晰[3-6],而不同封育季節放牧干擾對青海云杉種群的研究主要集中于空間格局[7]。本研究用種群年齡結構、種群生命表和生存函數分析不同放牧季節青海云杉種群的年齡結構、數量動態特點,以期為青海云杉林的保護和恢復提供理論依據。
1 研究區概況
哈溪林區位于祁連山北坡的東祁連山中部,102°01′~102°51′E,37°16′~37°45′N,海拔2 400~4 800m,氣候屬大陸性高寒半干旱氣候,年平均氣溫為1.8℃;年平均相對濕度50%~75%,年降水量350~450mm,多集中在6-9月;年蒸發量1 234.21mm,無霜期110d左右;發育有山生柳(Salix oritrepha)、鬼 箭 錦 雞 兒 (Caragana jubata)、杜鵑(Phododendron spp.)、金露梅(Dasiphphora fruticosa)等灌叢植被,祁連圓柏(Sabina przewalskii)林、青海云杉林等喬木植被和苔草(Carexspp.)、針茅(Stipaspp.)、珠芽蓼(Polygonum viviparum)草甸和草原植被以及高山灌叢草甸土、森林灰褐土、山地灰鈣土、山地栗鈣土等。青海云杉林是該林區代表性群落,主要分布于陰坡、半陰坡和半陽坡,是群落中唯一的建群種和優勢種,群落結構簡單,一般為喬木層、灌木層、草本層和苔蘚層4層,喬木層的郁閉度0.4~0.8,草本層蓋度60%以上。哈溪林區夏季牧場放牧一般從6月開始,11月結束,轉入冬季牧場至翌年5月,夏季牧場在冬季封育,冬季牧場在夏季封育。
2 材料與方法
2.1 調查方法 分別在哈溪天然林區選擇林分狀況和載畜量基本一致、代表冬季放牧(雀兒溝)和夏季放牧(寺陽洼)的草類-青海云杉林,用GPS定位,設置樣地進行群落調查,每個樣地20m×20m,并劃分成16個5m×5m的小樣方,共32個小樣方。對每一小樣方的植物進行調查,對喬木進行每木檢尺,記錄樹高、胸徑等指標及樹高<1.5m的幼苗、幼樹的樹高、地徑、株數,對灌草層進行植被調查,并記錄整個樣地生境條件。
2.2 年齡結構分析方法 種群年齡結構分析是揭示種群生存現狀和更新策略的重要途徑之一[8]。根據以往的調查數據和解析木資料,青海云杉的年齡(A)與胸徑(HD)存在線性關系[5,9]。
年齡10~40年:
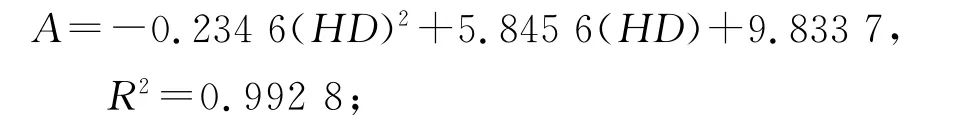
年齡40~150年:
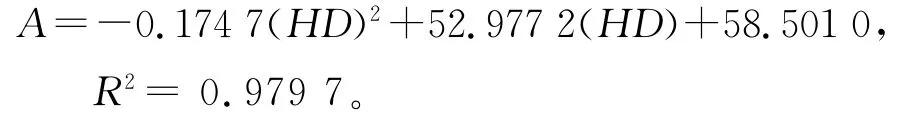
故用徑階代替年齡的方法,分析種群動態。
根據青海云杉胸徑、樹高的生長過程,按15年為1個齡級將樣地調查的青海云杉按照年齡與胸徑的關系劃分為8個齡級(表1)進行分析。
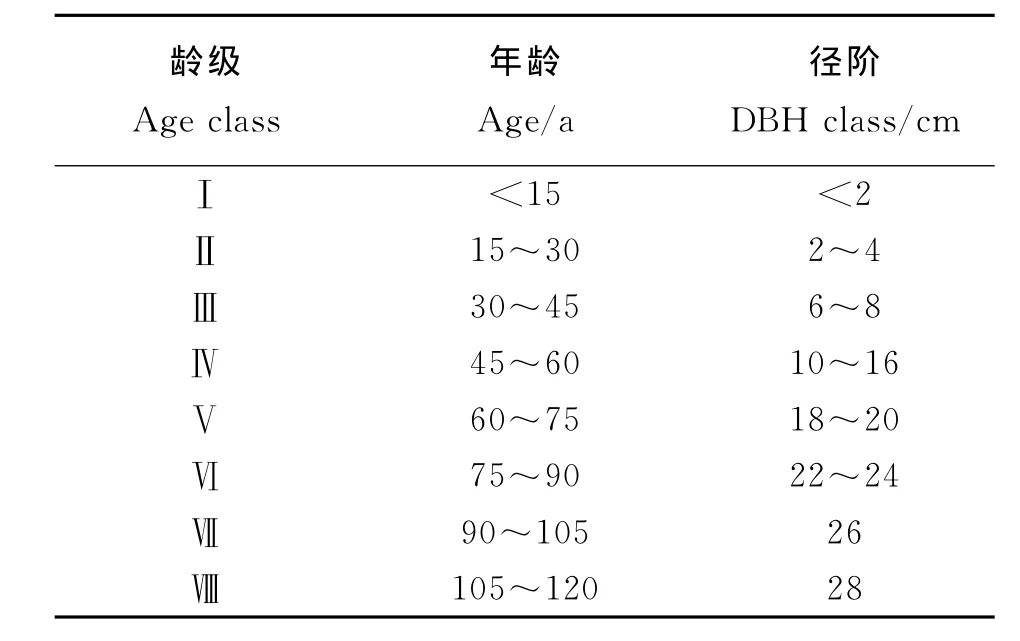
表1 青海云杉林齡級劃分Table 1 Age classes of Picea crassifolia forest
2.3 生命表數據的處理與編制 統計各齡級內的青海云杉株數,按照靜態生命表的編制方法[9-16],分別用哈溪林區不同放牧季節的林內青海云杉種群各齡級株數編制靜態生命表。調查樣地屬青海云杉異齡林,不完全滿足:1)種群數量是靜態的,即密度不變;2)年齡組合是穩定的,即種群的年齡結構與時間無關,各年齡的比例不變;3)個體的遷移是平衡的3個假設[15]。由于靜態生命表用特定時間段調查的種群所有個體編制而成,利用了異齡林種群年齡結構在空間上的差異,研究特定時間斷面上的種群動態,難免存在系統抽樣誤差,產生與數學假設技術不符的現象,但仍能提供有用的生態學記錄[9,11,16]。因此,勻滑(Smooth out)技術在編制靜態生命表時經常用到[9,11,13,16-17]。
以各生命表的齡級為橫坐標,齡級的標準化存活數的自然對數為縱坐標,繪制不同放牧季節的林內青海云杉種群的存活曲線。存活曲線是借助于存活 個體 數 量 來 描 述 特 定 年 齡 死 亡 率[9,11-12],Deevey(1947)把存活曲線分成3種類型:Ⅰ型為凸型,表示青海云杉種群的大多數個體均能實現其平均的生理壽命,在達到平均壽命時,幾乎同時死亡;Ⅱ型為對角線型;Ⅲ型為凹型,表示青海云杉幼苗幼樹死亡率高,以后的 死 亡 率 低 而 穩 定[8-9,11-12,16]。本 研 究 根 據存活量的對數值和齡級作圖,用直線、對數、指數、冪函數和多項式5種曲線進行擬合,選擇相關系數最大的曲線方程,并進行檢驗。
2.4 生存分析 為了更好地分析不同放牧季節對青海云杉種群結構的影響,本研究用4個種群生存函數,即種群生存率函數(St)、累計死亡率函數(Ft)、死亡密度函數(ft)和危險率函數(λt)進行分析,這4個函數用下列公式[4,11,15,18,25,27]計算:
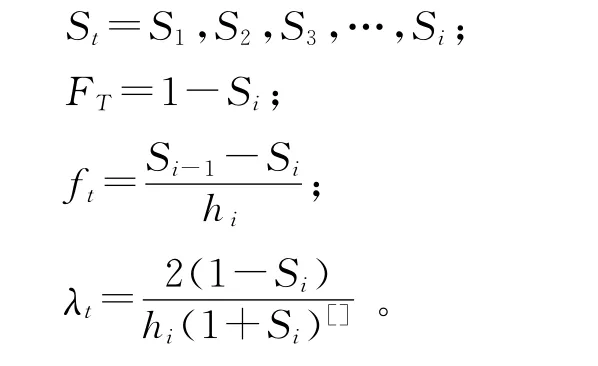
式中,hi為年齡寬度,St為存活率。
3 結果與分析
3.1 年齡結構分析 雀兒溝青海云杉處于中齡林階段,種群中幼苗幼樹的數量不足,不到寺陽洼的1/2,從Ⅰ齡級到Ⅱ齡級幼樹的存活率達67.5%,從Ⅳ齡級到Ⅴ齡級的存活率達21.4%,種群的存活率只有2.9%。寺陽洼青海云杉處于成熟林階段,種群中幼苗幼樹的數量豐富,從Ⅰ齡級到Ⅱ齡級幼樹的存活率只有16.5%,Ⅲ齡級后的存活率平緩下降,種群的存活率僅有0.8%。兩個樣地中,青海云杉種群均為增長型種群。不同放牧季節青海云杉種群的年齡結構在兩個地點存在差異(圖1)。
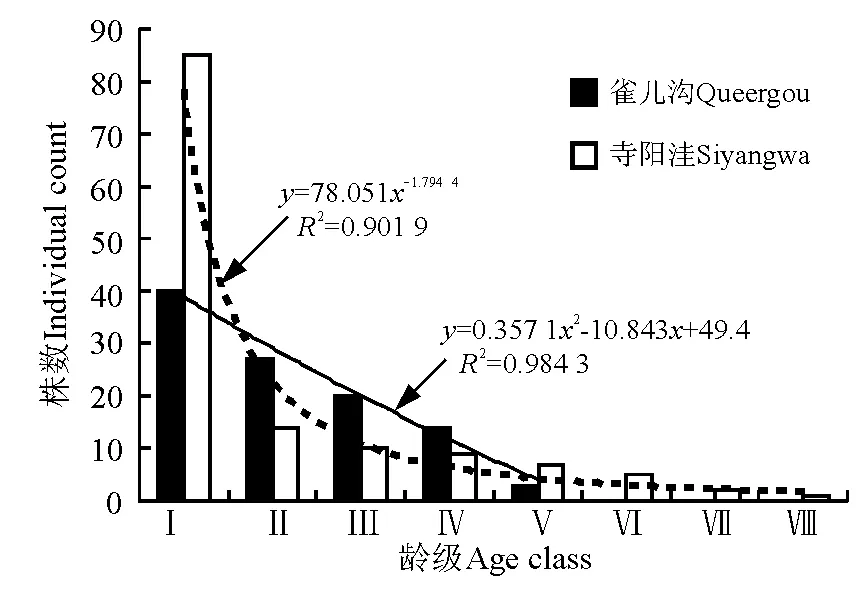
圖1 雀兒溝和寺陽洼樣地青海云杉種群的年齡結構Fig.1 Age structure of Picea crassifolia populations in Queergou and Siyangwa plots
3.2 種群靜態生命表 基于青海云杉林生長過程,對各齡級株數通過勻滑技術處理獲取,編制了不同放牧季節青海云杉種群特定時間的生命表(表2),其中雀兒溝青海云杉種群最大編表年齡為Ⅴ齡級,約75年,種群死亡量Ⅰ、Ⅳ齡級最高,死亡率在Ⅳ齡級最高,可能和自然稀疏有關;區間壽命、總壽命、期望壽命隨齡級的增加而降低;在Ⅱ齡級以后,消失率隨齡級的增加而增大,存活率Ⅱ齡級最大,以后隨齡級的增加而減小。寺陽洼青海云杉種群最大編表年齡為Ⅷ齡級,約120年,種群死亡量Ⅰ齡級最高,其次為Ⅱ、Ⅵ齡級;死亡率在Ⅰ和Ⅵ齡級最高,Ⅲ齡級最低,區間壽命、總壽命隨齡級的增加而降低,除Ⅰ齡級外,期望壽命隨齡級的增加而降低;消失率在Ⅰ、Ⅵ齡級最大;存活率Ⅲ齡級最大。
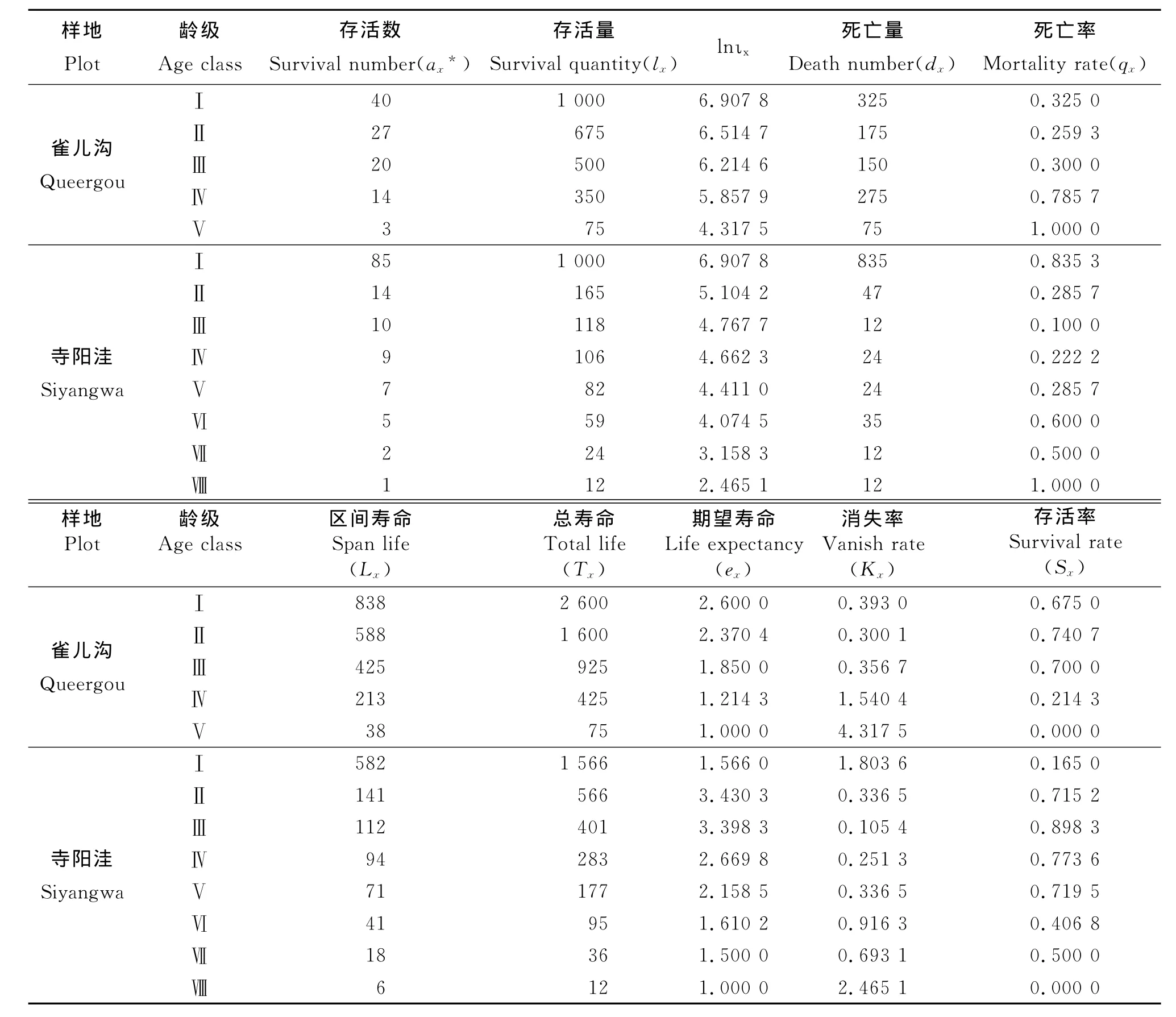
表2 雀兒溝和寺陽洼樣地青海云杉種群靜態生命表Table 2 Static life table of Picea crassifolia population in Queergou and Siyangwa plots
3.3 存活曲線分析 根據存活量的對數值和齡級作圖,得到不同放牧季節青海云杉種群的存活曲線(圖2)。擬合結果表明,冬季放牧(雀兒溝)的青海云杉林內,種群存活曲線接近DeeveyⅠ型,曲線符合拋物線方程模型:
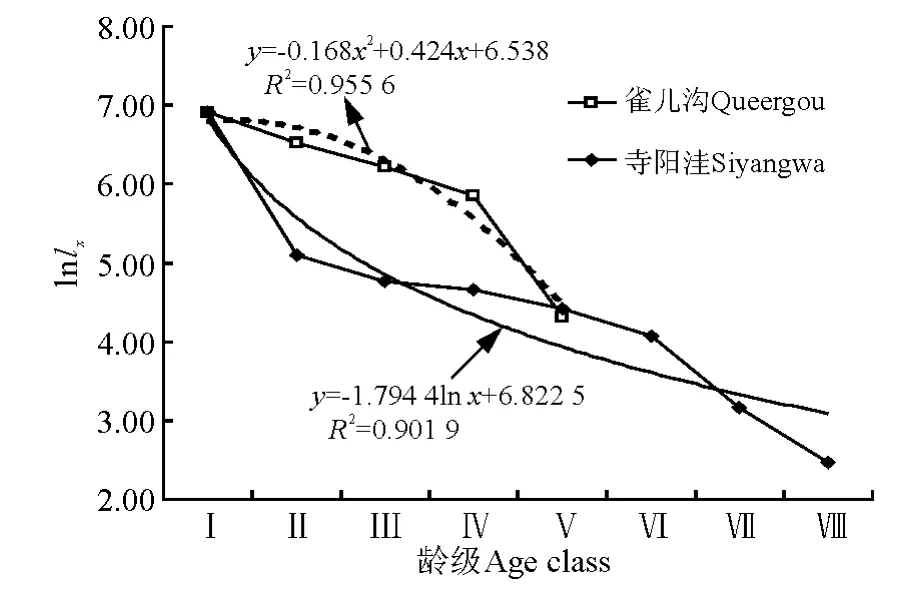
圖2 雀兒溝和寺陽洼樣地青海云杉種群存活曲線Fig.2 Survival curve of Picea crassifolia populations in Queergou and Siyangwa plots
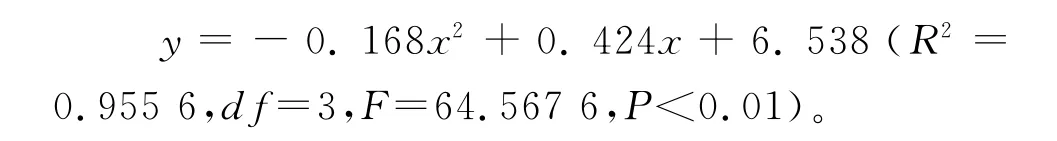
前期存活數呈緩慢下降趨勢,即前期死亡率較低,環境篩的選擇強度較小,有67.5%的幼苗能穿過此篩進入Ⅱ齡級,Ⅳ齡級以后死亡率增大;夏季放牧的青海云杉林內,種群存活曲線接近DeeveyⅡ型,曲線符合冪函數方程模型和對數函數方程模型:
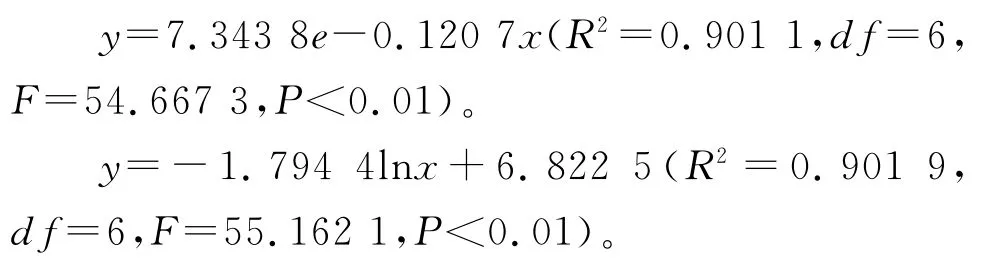
前期存活數呈下降趨勢,即前期死亡率較高,環境篩的選擇強度較大,只有16.5%的幼苗能穿過此篩進入Ⅱ齡級,Ⅱ齡級以后死亡率降低,至Ⅵ齡級死亡率再次增加。
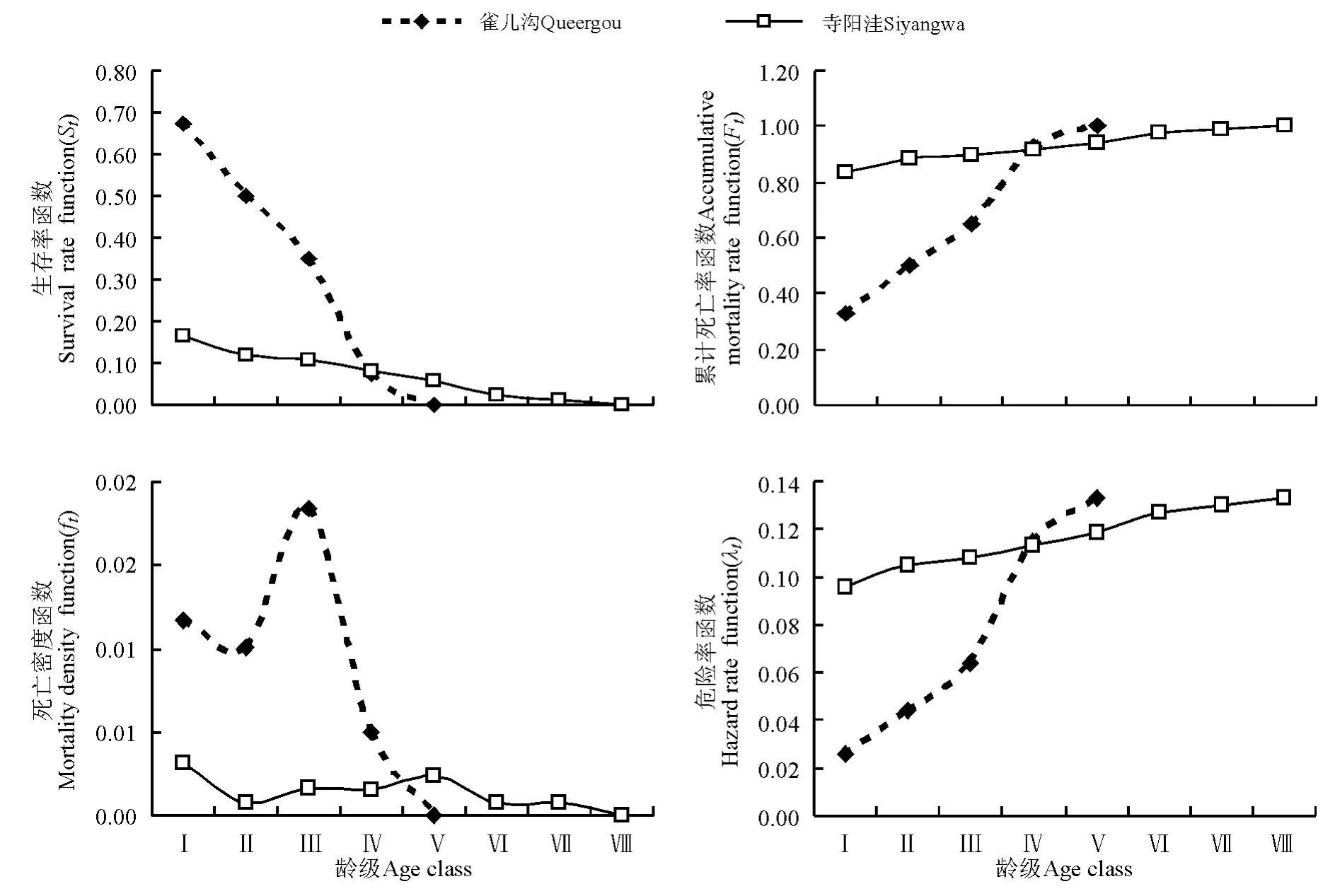
圖3 雀兒溝和寺陽洼樣地青海云杉種群生存函數Fig.3 Survival curve of Picea crassifolia populations in Queergou and Siyangwa plots
3.4 生存函數分析 根據4個生存函數的估算值,繪制生存率函數曲線(St)、累計死亡率函數曲線(Ft)、死亡密度函數曲線(ft)和危險率函數曲線(λt)(圖3)。雀兒溝和寺陽洼樣地青海云杉種群的生存曲線隨齡級增加都呈單調下降趨勢,生存率在Ⅰ齡級最大;在Ⅰ~Ⅲ齡級,相同齡級的種群,雀兒溝的生存率大于寺陽洼,種群存活到Ⅲ齡級的概率為35.0%,存活到Ⅳ齡級的概率則降低至7.5%,主要是種群種內和種間競爭加劇的結果;Ⅳ齡級的種群,寺陽洼的生存率大于雀兒溝,且Ⅰ~Ⅷ齡級的種群一直保持相對穩定的生存率。累計死亡率曲線和危險率曲線隨齡級增加都呈增加趨勢,雀兒溝增加快,而寺陽洼較為平緩。雀兒溝青海云杉種群的死亡密度函數在Ⅲ齡級達最大,為1.83%,寺陽洼青海云杉種群的死亡密度函數在Ⅴ齡級達最大,為0.23%。寺陽洼青海云杉種群的4個函數變幅較小,而雀兒溝的變幅較大。
4 討論與結論
種群的齡級結構、生命表、存活曲線、生存函數是種群發育過程中種群的生物學特性與環境因素共同作用的外在表現,冬季放牧和夏季放牧對青海云杉種群產生了深刻的影響,特別是干擾了幼苗、幼樹的生存環境,冬季放牧的影響尤為嚴重。
盡管冬季放牧和夏季放牧的青海云杉種群屬增長型種群,但種群年齡結構存在明顯的幼樹比例偏小的缺陷,造成幼樹有較強的穿過環境篩能力的假象。自然狀態下,草類-青海云杉林內青海云杉種群的存活曲線符合DeeveyⅢ型[4],而冬季牧場的青海云杉林內,種群存活曲線接近DeeveyⅠ型。牲畜踐踏,破壞了地表覆蓋物,造成土壤干旱,種子難以發芽,幼苗受到放牧的嚴重干擾;同時,由于冬季林下牧場飼草缺乏,牲畜踐踏和啃食,直接造成幼苗幼樹死亡,林內更新受到嚴重影響。夏季放牧的青海云杉林內,種群存活曲線接近DeeveyⅡ型。由于地表覆蓋物的保護和林下飼草逐漸增加,牲畜踐踏和啃食對種子萌發和幼樹生長的干擾弱于冬季放牧。因此,兩種放牧季節對草類-青海云杉林都產生了影響,其中冬季放牧嚴重破壞了種子萌發和幼苗生長環境,對青海云杉林的保護和恢復十分不利。
冬季牧場的青海云杉種群個體數量明顯低于夏季牧場,4個生存函數反映出種群退化的風險大于夏季放牧的青海云杉種群;夏季放牧的青海云杉種群保持相對穩定的生存率和死亡率,反映出幼樹的儲備不足,種群結構存在一定的缺陷,同樣存在退化的風險。
[1] 汪有奎,楊全生,李世霞,等.石羊河流域上游祁連山植被恢復與重建對策[J].中國科技成果,2008(9):18-19.
[2] 劉建泉,車克鈞,馬世貴,等.祁連山森林增量提質的重要性及其對策[J].中國林學會:西北生態環境論壇—西北地區生態環境建設研討會論文專輯[C].北京:中國林業出版社,2001:219-223.
[3] 劉建泉.祁連山保護區青海云杉種群分布格局的研究[J].西北林學院學報,2004,19(2):152-155.
[4] 丁國民,劉興明,倪自銀,等.甘肅祁連山青海云杉種群數量動態的初步研究[J].植物資源與環境學報,2005,14(4):36-41.
[5] 劉建泉,丁國民,郝虎,等.青海云杉群落特征和動態的研究[J].西北林學院學報,2008,23(1):14-17.
[6] 劉建泉.祁連山林區青海云杉群落種間關系和聯結分析[J].西北林學院學報,2009,24(2):18-22.
[7] 劉建泉,楊全生,汪有奎,等.不同封育季節放牧干擾對青海云杉種群空間格局的影響[J].南京林業大學學報(自然科學版),2010,34(6):53-36.
[8] 宋于洋,趙自玉,楊振安,等.天山云杉種群數量動態研究[J].南京林業大學學報(自然科學版),2009,33(1):64-68.
[9] 江洪.云杉種群生態學[M].北京:中國林業出版社,1992:8-26.
[10] 沈澤昊,方精云,劉增力,等.貢嘎山海螺溝林線附近峨眉冷杉種群的結構與動態[J].植物學報,2001,43(12):1288-1293.
[11] 洪偉,王新功,吳承禎,等.瀕危植物南方紅豆杉種群生命表及譜分析[J].應用生態學報,2004,15(6):1109-1112.
[12] 肖宜安,何平,李曉紅,等.瀕危植物長柄雙花木自然種群數 量 動 態 [J].植物 生 態 學 報,2004,28(2):252-257.
[13] 程偉,吳寧,羅鵬.岷江上游林線附近岷江冷杉種群的生存分析[J].植物生態學報,2005,29(3):349-353
[14] 陳國科,彭華.不同人為干擾條件下毒藥樹種群數量特征 的 比 較 [J].植 物 生 態 學 報,2006,30(3):426-431.
[15] 張橋英,張運春,羅鵬,等.白馬雪山陽坡林線方枝柏種群的生態特征[J].植物生態學報,2007,31(5):857-864.
[16] 張志祥,劉鵬,蔡妙珍,等.九龍山珍稀瀕危植物南方鐵杉種群數量動態[J].植物生態學報,2008,32(5):1146-1156.
[17] 張憲強,郭衛華,楊繼紅,等.刺槐(Robinia pseudoacacia)無性系種群結構與生長動態的研究[J].山東大學學報(理學版),2006,41(2):135-139.
[18] 何亞平,費世民,蔣俊明,等.不同齡級劃分方法對種群存活分析的影響——以水災跡地油松和華山松種群分 析 為 例 [J].植 物 生 態 學 報,2008,32(2):448-455.
