松嫩平原星星草果后營養期無性系結構及其生長分析
2012-08-20 08:01:58張春艷張春雨陳玉交李海燕楊允菲
草業科學 2012年6期
張春艷,張春雨,陳玉交,李海燕,楊允菲
(東北師范大學草地科學研究所 植被生態科學教育部重點實驗室,吉林 長春130024)
多年生植物的果后營養期是冬眠構件形成和養分向地下儲藏的重要階段[1]。對于禾本科植物來說,果后營養期形成的分株不一定都是冬眠構件,只有那些不拔節的苗期分株因生長點位于地表以下,才能進入冬眠,下一年返青繼續生長[2-3]。叢生型植物的每一個分蘗叢都是界限分明的獨立無性系,了解其各生長階段構件結構及無性系各數量性狀之間的關系具有理論和實踐意義。
星星草(Puccinellia tenuiflora)是多年生叢生型禾草,具有耐鹽堿性強、飼用價值高等特點,是治理天然鹽堿化草地和利用價值最大的牧草之一,在松嫩平原堿化草甸廣泛分布,常形成大面積單優種群落[4]。星星草分蘗力強,春季返春早,果后營養期長,營養豐富,其飼用價值是鹽生植被中較高的優良牧草[5]。有關星星草的形態解剖的結構特征[6]、生理 生 化 機 制[7-9]、生 物 生 態 學 特 性[1,10-12]、生 產 特性[13-16]、種群數 量 特 征[2,17]等 研 究 已 有 報 道。 但 對于星星草無性系水平的定量研究僅有較少報道[18]。本研究以實地調查為依據,從無性系水平定量分析了松嫩平原果后營養期星星草各構件的數量和生物量的結構,以及各組分數量性狀與無性系大小之間的關系,為叢生無性系植物種群生態學的深入研究累積資料,為松嫩平原堿化草地的利用與管理提供科學依據。
1 研究地區與研究方法
1.1 試驗地概況 試驗地設在松嫩平原南部的吉林省長嶺種馬場、東北師范大學草地生態研究站(44°38′N,123°41′E)。該地區位于松嫩平原南部沖積平原,地勢平坦,海拔137.8~144.8m,屬于典型的大陸性季風氣候。年平均氣溫4.9℃,≥10℃的年積溫2 545~3 374℃·d,無霜期136~163d。6-8月平均氣溫超過20℃,7月氣溫最高;年降水量313~581mm,主要集中在6-9月,約占全年降水量的60%以上,年蒸發量一般為年降水量的2~3倍[19-21]。在松嫩平原鹽堿化草甸植被中,星星草在低洼地段經常形成單一優勢種群落,也容易成為獐毛(Aeluropus sinensis)、黃 蒿 (Artemisia scoparia)、虎尾草(Chloris virgata)、堿蒿(A.anethifolia)等其他鹽堿群落的伴生種。
1.2 研究方法 在松嫩平原,星星草種群于6月上、中旬開花,6月末籽實成熟。本研究在東北師范大學草地生態研究站的天然鹽堿化草甸的星星草群落進行,于2009年7月中旬,在星星草果后營養期,采用整個分蘗叢挖掘的方法,隨機挖取30個大小不同的星星草分蘗叢。帶回室內逐一測定無性系的叢徑后,將營養株從分蘗節處掰下,按拔節的分蘗株和未拔節的分蘗苗分別計數,并放置于80℃下烘至質量不變后用1/1 000電子天平稱量。
1.3 數據分析 統計分析各樣地所測定的數量指標,用最大值和最小值反映觀測樣本的實際大小范圍,用平均數代表樣本各性狀的整體水平,用標準差(SD)反映樣本的絕對變異度,用變異系數(CV)反映樣本的相對變異度[22]。選取直線函數、冪函數、指數函數和對數函數的4種函數模型回歸分析各性狀間的關系,對各回歸模型的相關系數作顯著性檢驗,比較模型的相關系數,并選用相關程度較高的函數方程作為定量化描述模型。
2 結果與分析
2.1 構件的數量特征及結構分析 星星草無性系叢徑的整體水平為(2.65±1.06)cm,叢徑的變異系數最小,除分蘗苗生物量外,其他各項指標的變異系數大約為叢徑的1.5~1.6倍,而以分蘗苗生物量的變異系數最大,為叢徑的1.9倍,表明星星草正處于果后營養期的生長階段,生物量隨著生長而變化;分蘗株數量與分蘗苗數量的變異系數二者差異相對較小;總生物量與分蘗株生物量二者的變異度基本處于同一水平,而明顯低于分蘗苗的變異度(表1)。同時,星星草無性系構件是由分蘗株和分蘗苗組成,分蘗株數量占總體構件數量的53.3%,是分蘗苗的1.14倍;分蘗株生物量居優勢地位,占總體生物量的90.6%,是分蘗苗生物量的9.58倍(表1)。由此表明,在果后營養期,分蘗株是無性系的主要組成部分。
2.2 構件數量與叢徑的關系 對于無性系植物來說,衡量無性系大小的數量指標之一是叢徑。經統計分析,星星草分蘗株和分蘗苗數量與叢徑之間均呈極顯著直線正相關關系(圖1)。從圖1直線方程斜率值可以反映出,在叢徑0.6~4.6cm范圍內,理論上叢徑每增加1cm,將增加11.607個分蘗株和10.565個分蘗苗,兩者增長速率大體相同,反映了分蘗株與分蘗苗的數量增長具有同步生長規律。而總構件數與叢徑之間呈極顯著冪函數關系,增長的冪值為1.156 7,由此反映了總構件數與不同組分增長的方式不同,并表明總構件數生長要快于叢徑的增長。
2.3 構件生物量與叢徑的關系 星星草無性系分蘗株生物量、分蘗苗生物量、總生物量與其叢徑之間也有較好的正相關關系(圖2)。經回歸分析分蘗株生物量與叢徑呈極顯著直線相關關系,分蘗苗生物量與叢徑呈極顯著對數函數關系,總生物量與叢徑呈極顯著冪函數關系。由此表明三者的增長方式不同,但都與叢徑呈極顯著的正相關關系,反映了叢徑的大小制約著無性系的物質生產。
2.4 不同構件生物量與其數量的關系 經相關性分析,星星草無性系分蘗株生物量與分蘗株數量、總生物量與總構件數量呈極顯著直線函數關系,分蘗苗生物量與分蘗苗數量呈極顯著冪函數關系(圖3)。就兩個直線方程的斜率值比較,分蘗株生物量生長速率是總生物量的1.8倍,由此間接地反映了分蘗苗生物量增長率遠小于分蘗苗數量的增長率。

表1 星星草無性系構件的數量特征Table 1 Quantitative characters on modules of Puccinellia tenuiflora clone(n=30)
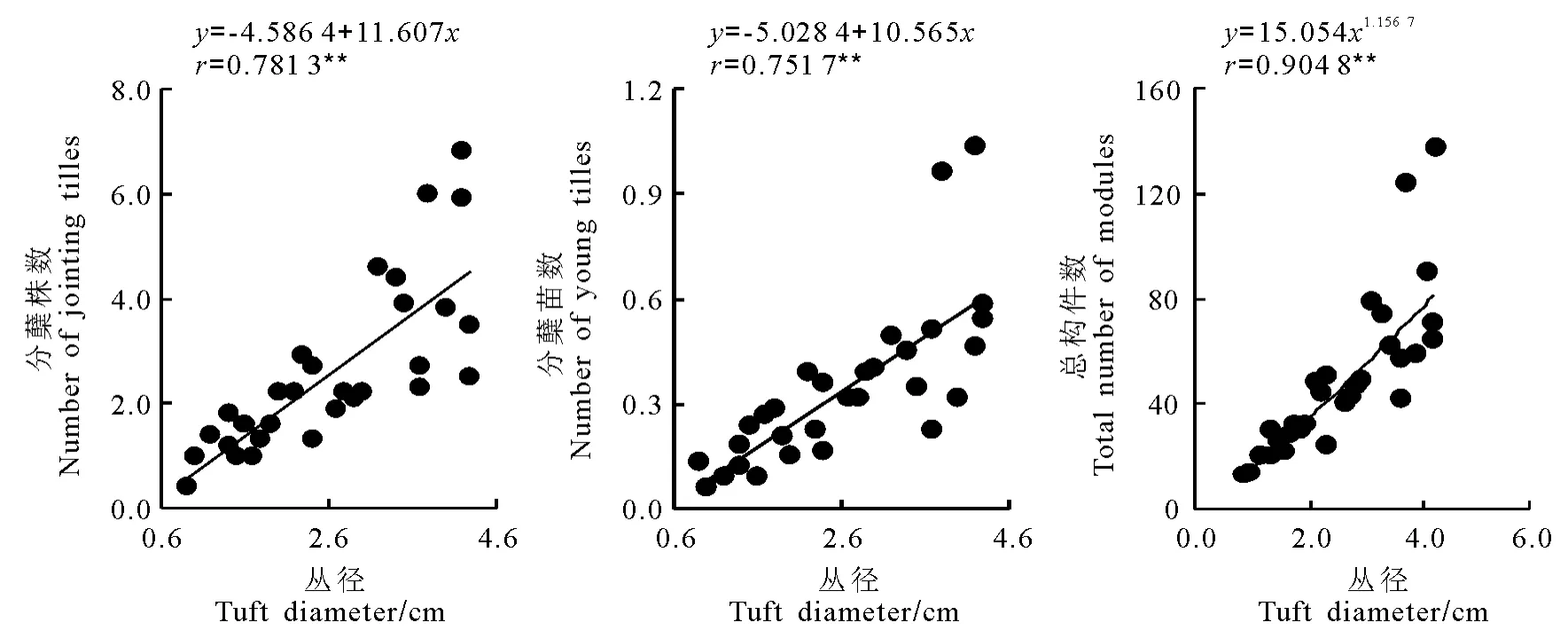
圖1 星星草無性系不同構件的數量與叢徑的觀測值及擬合曲線(n=30)Fig.1 Observed data and the simulated curves on relationships between the number of different modules and the tuft diameter in Puccinellia tenuiflora clone(n=30)
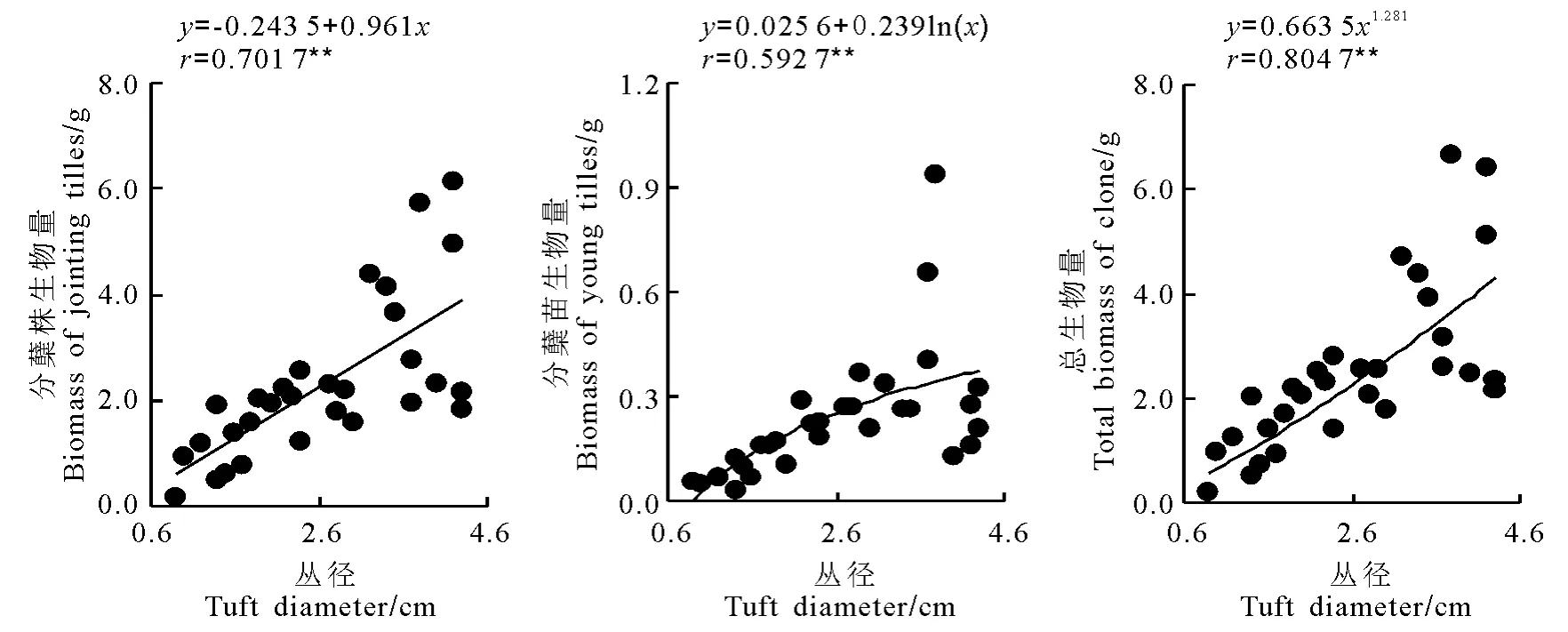
圖2 星星草無性系不同構件的生物量與叢徑的觀測值及擬合曲線(n=30)Fig.2 Observed data and the simulated curves on relationships between biomass of different modules and the tuft diameter in Puccinellia tenuiflora clone(n=30)
3 討論與結論
天然鹽堿化草甸群落中,星星草的分蘗叢大小參差不齊,主要是這些無性系并非同一時間定居的同生群,致使星星草無性系構件的數量性狀均變異較大(表1)。一般情況下,種子萌發與定居越早,其無性系生長的時間就越長,營養繁殖的構件越多,叢徑就越大,反之亦然。此外,由于植物固著生長,種子萌發與定居微地形的不同,將引起微環境的差異。即任何群落內在土壤質地的均勻性、土壤小動物的干擾等方面總是存在著不同程度的差異,即使是同時播種與相同管理下的栽培草地,其無性系構件的數量性狀也存在較大的生態可塑性[20,23-24]。
不同生活史階段的個體組成植物種群,而種群的生存與發展,以及種群的調節體現在不同生活史階段個體的數量和生物量的配置[20,25]。在整個生長季的不同生育期,植物種群組成的生活史格局必然發生變化。在松嫩平原天然草地,星星草果后營養期達3個月(7-9月)之久,其中,果后營養期前期形成的分蘗大多數可進入拔節生長,而后期則大部分以芽的階段冬眠[1,26]。本研究只是星星草果后營養期前期的構件結構特征,其中,后期隨著新構件的不斷補充,其結構的變化過程有待進一步研究。
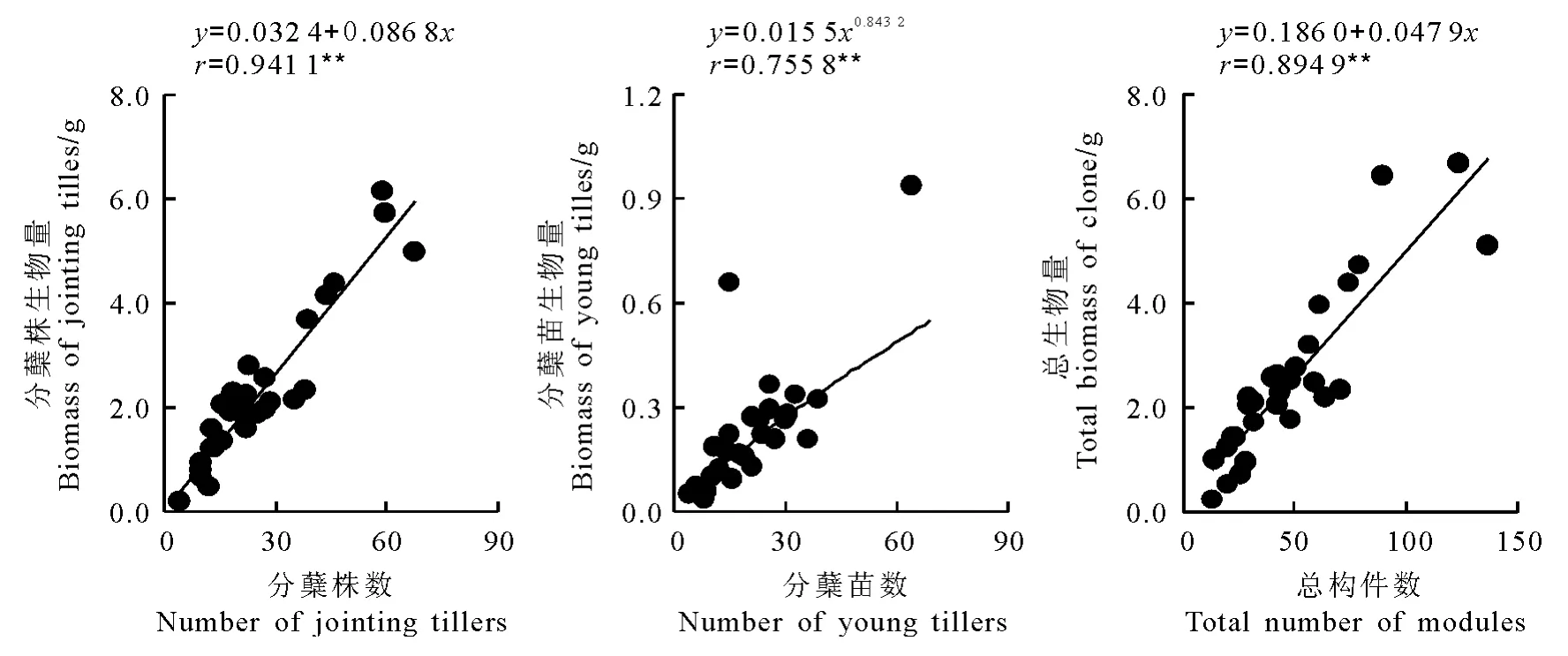
圖3 星星草無性系不同構件的生物量與數量的觀測值及擬合曲線(n=30)Fig.3 Observed data and the simulated curves on relationship between the biomass and the number of different modules in Puccinellia tenuiflora clone(n=30)
伴隨著構件的形成和生長,無性系在不斷增大。每一個邊界明顯的叢生型禾草的無性系(分蘗叢)均可視為一個小種群,其構件增加和空間擴展均極為有序。在松嫩平原天然草地星星草果后營養期前期,星星草無性系各構件的數量與叢徑之間,各構件的生物量與叢徑之間,各構件的生物量與其數量之間均具有極顯著(P<0.01)的相關關系,大體呈線性、對數和冪函數規律變化,這與植物個體的生長及生產分配有著相同的規律[14,22]。但是隨著生長季的進程,這些數量性狀之間的規律是否發生變化有待進一步研究。
[1] 楊允菲,祝玲.松嫩平原十五種多年生禾草種群營養繁殖體冬眠特性的分析[J].草業科學,1994,11(2):26-31.
[2] 張麗輝,趙驥民,楊允菲.放牧和割草利用對星星草種群冬眠構件的影響[J].草業科學,2006,23(4):8-11.
[3] 宋金枝,楊允菲.松嫩平原堿化草甸朝鮮堿茅無性系冬眠構件的結構及生長分析[J].生態學雜志,2006,25(7):743-746.
[4] 鄭慧瑩,李建東.松嫩平原的草地植被及其利用保護[M].北京:科學出版社,1993:40-44.
[5] 蘇加楷,張文淑,李敏.星星草、牧草高產栽培[M].北京:金盾出版社,1993:115-116.
[6] 陸靜梅,李建東,景德章,等.星星草Puccinellia tenuiflora(Turcz.)Scribn.et Merr.解剖研究[J].東北師大學報(自然科學版),1994(1):63-66.
[7] 閻秀峰,孫國榮,李敬蘭,等.羊草和星星草光合蒸騰日變化的比較研究[J].植物研究,1994,14(3):287-291.
[8] 王蘋,李建東,歐勇玲.松嫩平原鹽堿化草地星星草的適應性及耐鹽生理特性的研究[J].草地學報,1997,5(2):80-84.
[9] 肖瑋,孫國榮,閻秀峰,等.松嫩鹽堿草地星星草種群地上生物量的季節動態[J].哈爾濱師范大學自然科學學報,1995,11(1):81-83.
[10] 楊允菲,祝玲,李建東.松嫩平原堿化草甸星星草種群營養繁殖及有性生殖的數量特征[J].應用生態學報,1995,6(2):166-171.
[11] 張春華,楊允菲,李建東.兩種堿茅種群果后營養繁殖的定量分析[J].中國草地,1996(1):31-35.
[12] 竇聲云,周學麗,莫玉花.Na2CO3脅迫對老芒麥和星星草種子萌發的影響[J].草業科學,2010,27(9):124-127.
[13] 楊允菲,李建東.松嫩平原堿化草甸三種多年生鹽堿禾草群落生產特征的分析[J].草業科學,1994,11(3):34-35.
[14] 楊允菲,李建東.松嫩平原全葉馬蘭種群不同年齡級分株的生長策略[J].應用生態學報,2003,14(12):2171-2175.
[15] 張春和,李建東.星星草群落地上生產結構、現存量季節動態和凈初級生產力的研究[J].草業學報,1995,4(1):36-43.
[16] 郭孝,張莉.朝鮮堿茅與星星草經濟性狀的研究[J].河南農業科學,1996(11):34-36.
[17] 張麗輝,趙驥民,李帥.松嫩平原不同生境星星草無性系種群的數量特征分析[J].長春師范學院學報(自然科學版),2007,26(4):58-61.
[18] 張麗輝,楊允菲.松嫩平原星星草無性系生殖株的數量特征[J].中國科學學報,2006,3(7):78-81.
[19] 李建東,鄭慧瑩.松嫩平原鹽生植物與鹽堿化草地的恢復[M].北京:科學出版社,1997:1-5.
[20] 楊允菲,張寶田,李建東.松嫩平原人工草地野大麥無性系冬眠構件的結構及形成規律[J].生態學報,2004,24(2):268-273.
[21] 楊允菲,李建東.東北草原羊草種群單穗數量性狀的生態可塑性[J].生態學報,2001,21(5):752-758.
[22] 楊允菲,李建東.松嫩平原不同生境蘆葦種群分株的生物量分配與生長分析[J].應用生態學報,2003,14(1):30-34.
[23] 楊允菲,張寶田.松嫩平原貝加爾針茅無性系構件的結構及生長規律[J].應用生態學報,2005,16(12):2311-2315.
[24] 張經經,楊允菲,陳永強.松嫩平原扁稈藨草種群生殖株構件表型可塑性及生長分析[J].草業科學,2011,28(11):1957-1961.
[25] 楊允菲,張寶田.松嫩平原人工草地野大麥無性系構件的生物量結構及生產規律[J].應用生態學報,2004,15(8):1378-1382.
[26] 李海燕,李建東,徐振國,等.內蒙古圖牧吉自然保護區羊草種群營養繁殖特性的比較[J].草業學報,2011,20(5):19-25.
