黃瓜把長和瓜長的遺傳分析
2012-08-08 12:23:08秦智偉王桂玲孫夢陽
東北農業大學學報 2012年1期
趙 鵬,秦智偉,王桂玲,魯 明,孫夢陽
(1.東北農業大學生命科學學院,哈爾濱 150030;2.東北農業大學園藝學院,哈爾濱 150030)
黃瓜(Cucumis sativusL.)又稱胡瓜、王瓜、青瓜,原產于印度喜馬拉雅山脈南麓的熱帶潮濕地區,在植物分類學上隸屬于葫蘆科(Cucurbitaceae)黃瓜屬(Cucumis)[1],為一年生草本蔓生攀緣植物,廣泛分布于我國各地,是與大眾生活密切相關的重要蔬菜作物之一。黃瓜的把長和瓜長是構成黃瓜果實外觀品質的重要性狀。近年來,隨著人民生活水平的不斷提高,消費者對黃瓜品質的要求越來越高,其中外觀品質是引起消費者購買欲望的直接因素,如色澤、口感、把長、瓜長、黃色線、彎曲度、果肉顏色及刺瘤稀密等等。因此,黃瓜的把長和瓜長直接影響著黃瓜果實的經濟價值和商品價值。
對許多數量性狀的研究發現,控制數量性狀的多個基因在效應上是不等的,效應大的表現為主基因,效應小的表現為多基因。蓋鈞鎰、王建康、章元明等在前人的研究理論基礎上將混合分布理論與數量遺傳學有機結合起來,提出了一套完整的分析植物數量性狀主基因與多基因存在和效應的方法,其方法的建立突破了原有數量遺傳學的理論假設,對分離世代的分析提供了植物數量性狀主基因與多基因兩方面的量化分析[2-6]。目前,這種方法已經應用到多種作物遺傳機制的研究當中。在黃瓜果實數量性狀的遺傳分析方面,現在已經進行了較多的研究,如果實彎曲性[7]、果皮葉綠素含量[8]、果肉顏色[9]、黃色線與瓜長比[10]以及單瓜重、瓜條直徑、把長、瓜長[11-15]等,這對我國的黃瓜品種改良和遺傳育種研究都起到了重要的指導作用。本研究以把長和瓜長差異較大的兩個黃瓜品種,利用植物數量性狀主基因+多基因混合遺傳模型分離分析法,對黃瓜F6代RIL群體的把長和瓜長進行了遺傳分析,為黃瓜外觀品質的改良提供了理論基礎,以加快我國黃瓜品質育種的進程,縮短育種周期,具有理論價值和實踐意義。
1 材料與方法
1.1 材料
本試驗材料由東北農業大學園藝學院瓜類課題組提供,包括溫室黃瓜品種拉迪特(Z9)和東農黃瓜品種129衍生的150個黃瓜F6代重組自交家系(RIL)及其親本。自交及繁育工作在東北農業大學園藝站和香坊實驗實習基地完成。
1.2 方法
1.2.1 田間試驗
2010年7月下旬在東北農業大學園藝站內播種育苗,8月初定植于東北農業大學香坊實驗實習基地大棚內,采用隨機區組設計,親本P1、P2各種植20株,F6代種植150株,行距60 cm,株距35 cm,3次重復,栽培管理同一般生產水平。
1.2.2 性狀調查
記錄各單株第一朵雌花的開花日期,選取節位在8~12節之間、商品性良好的成熟商品瓜,于同一日采摘,當日記錄把長和瓜長,每個單株取樣2~3次,計算家系重復平均值。調查方法見表1。

表1 把長與瓜長的調查方法Table 1 Survey method of carpopodium length and fruit length
1.2.3 統計分析
采用Excel 2007對親本及F6代把長和瓜長的表型數據進行描述性統計、正態分布檢驗和遺傳相關分析,并繪制次數分布圖。
1.2.4 遺傳分析
采用蓋鈞鎰等主基因+多基因混合遺傳模型對親本及把長和瓜長的家系重復平均值數據進行分離分析[2]。通過極大似然法和IECM(Iterated expectation and conditional maximization)算法計算出混合分布中各個成分分布參數,然后根據最大概率熵AIC準則選擇AIC值相對低的一個或幾個候選模型,并對其進行一組適合性測驗,共有5個統計量,包括Ul2、U22、U32、nW2和Dn,其中Ul2、U22、U32為均勻性檢驗,nW2為Smirnov檢驗,Dn為Kolmogorov檢驗。選擇統計量達到顯著水平且個數較少的模型作為黃瓜把長和瓜長的最適遺傳模型,根據最適遺傳模型的成分分布參數,用最小乘二法估算相應的遺傳參數,包括主基因與多基因的加性效應、上位性效應、遺傳方差和遺傳率等。采用南京農業大學提供的遺傳分析軟件進行遺傳分析。
2 結果與分析
2.1 統計分析
采用Excel 2007對親本及F6代各個單株的把長和瓜長表型數據進行描述性統計后,結果表明,母本“拉迪特Z9”和父本“東農129”的把長和瓜長都存在著明顯差異(見表2)。從F6代把長與瓜長的次數分布圖可以看出,兩個性狀均呈正態分布(見圖1、2),偏度值分別為1.28和0.24,說明把長和瓜長都是數量遺傳,而這兩個性狀的偏度值均大于0,呈現左偏分布,表明主基因控制黃瓜把長和瓜長的表現,需做進一步的遺傳分析。此外,遺傳相關分析表明,把長與瓜長的相關值為0.538**,呈正相關極顯著水平。
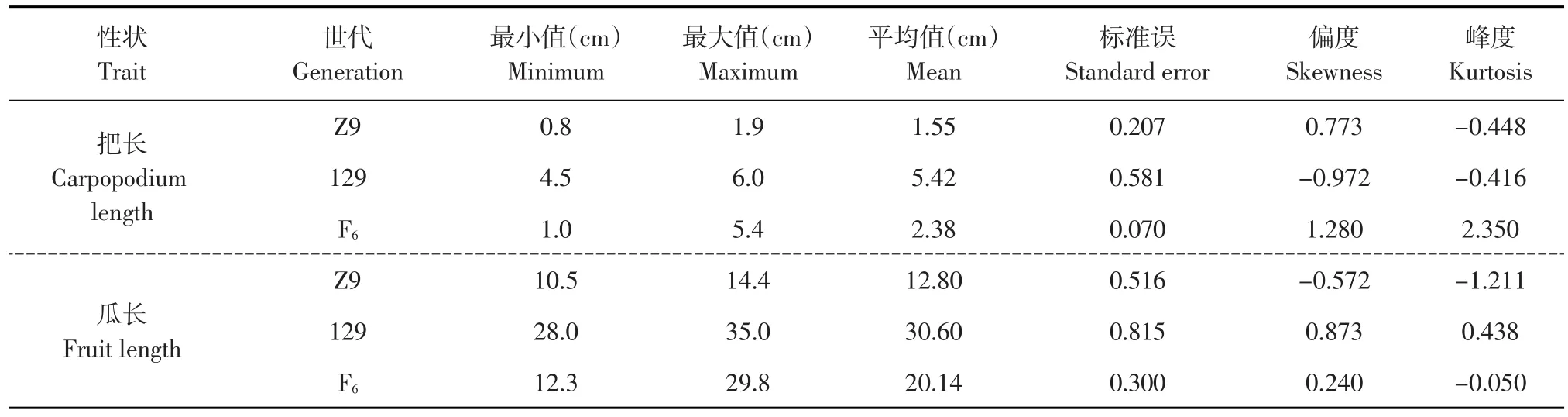
表2 黃瓜把長與瓜長的統計分析Table 2 Statistic analysis of carpopodium length and fruit length in cucumber
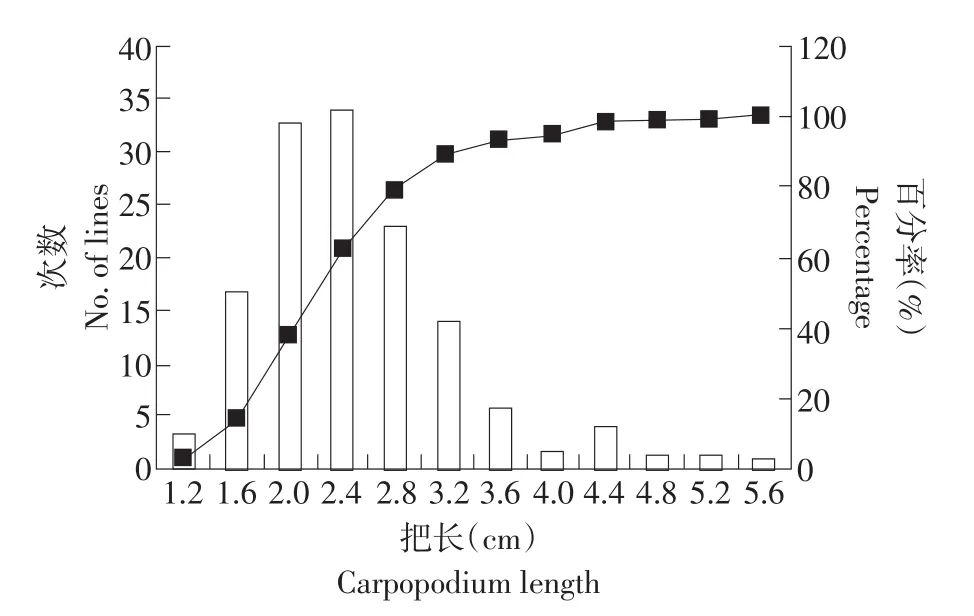
圖1 黃瓜把長的次數分布Fig.1 Frequency distribution of carpopodium length in cucumber
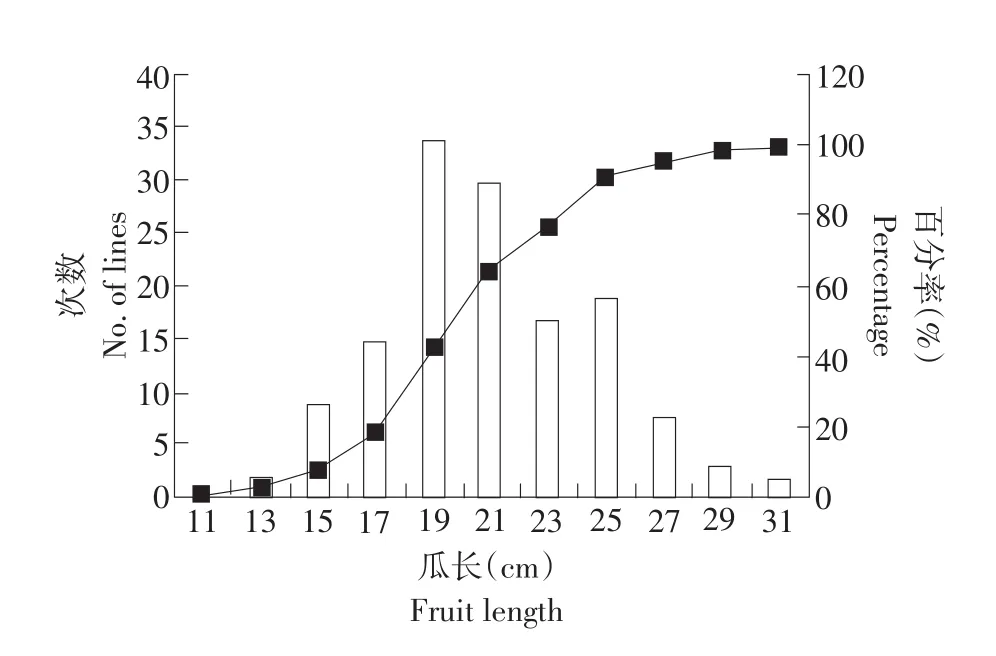
圖2 黃瓜瓜長的次數分布Fig.2 Frequency distribution of fruit length in cucumber
2.2 遺傳模型的選擇
利用主基因+多基因混合遺傳模型分離分析方法對把長和瓜長的表型數據進行分析后,得到7類遺傳模型,分別是一對主基因(代號為A),兩對主基因(代號為B),多基因(代號為C),一對主基因+多基因(代號為D),兩對主基因+多基因(代號為E),三對主基因(代號為F)和三對主基因+多基因(代號為G),包括加性-加性×加性上位性、加性、等加性、顯性上位、隱性上位、累加作用、重疊作用、互補作用和抑制作用,兩對主基因考慮有無上位性效應、有無連鎖等共53種遺傳模型(見表3)。
根據AIC準則,在把長的遺傳模型中,模型E-1-1、E-1-6和E-2-6的AIC值相對其他模型較低,經過一組適合性檢驗(見表4),模型E-1-1有4個統計量達到顯著水平,模型E-1-6有2個統計量達到顯著水平,而模型E-2-6只有1個統計量達到顯著水平,因此選擇E-2-6為把長遺傳的最適模型,即把長表現為兩對具有累加作用的連鎖主基因+多基因混合遺傳。
在瓜長的遺傳模型中,模型E-1-7、E-1-8和E-1-9的AIC值相對其他模型較低,經過一組適合性檢驗(見表4)。模型E-1-9有2個統計量達到顯著水平,而模型E-1-7和E-1-8只有1個統計量達到顯著水平,因此選擇模型E-1-7和E-1-8都可以,但經似然比檢驗這兩個模型的差異顯著性,結果均為差異顯著,而模型E-1-8的AIC值比E-1-7模型低,因此選擇E-1-8為瓜長遺傳的最適模型,即瓜長表現為兩對具有重疊作用的主基因+多基因混合遺傳。
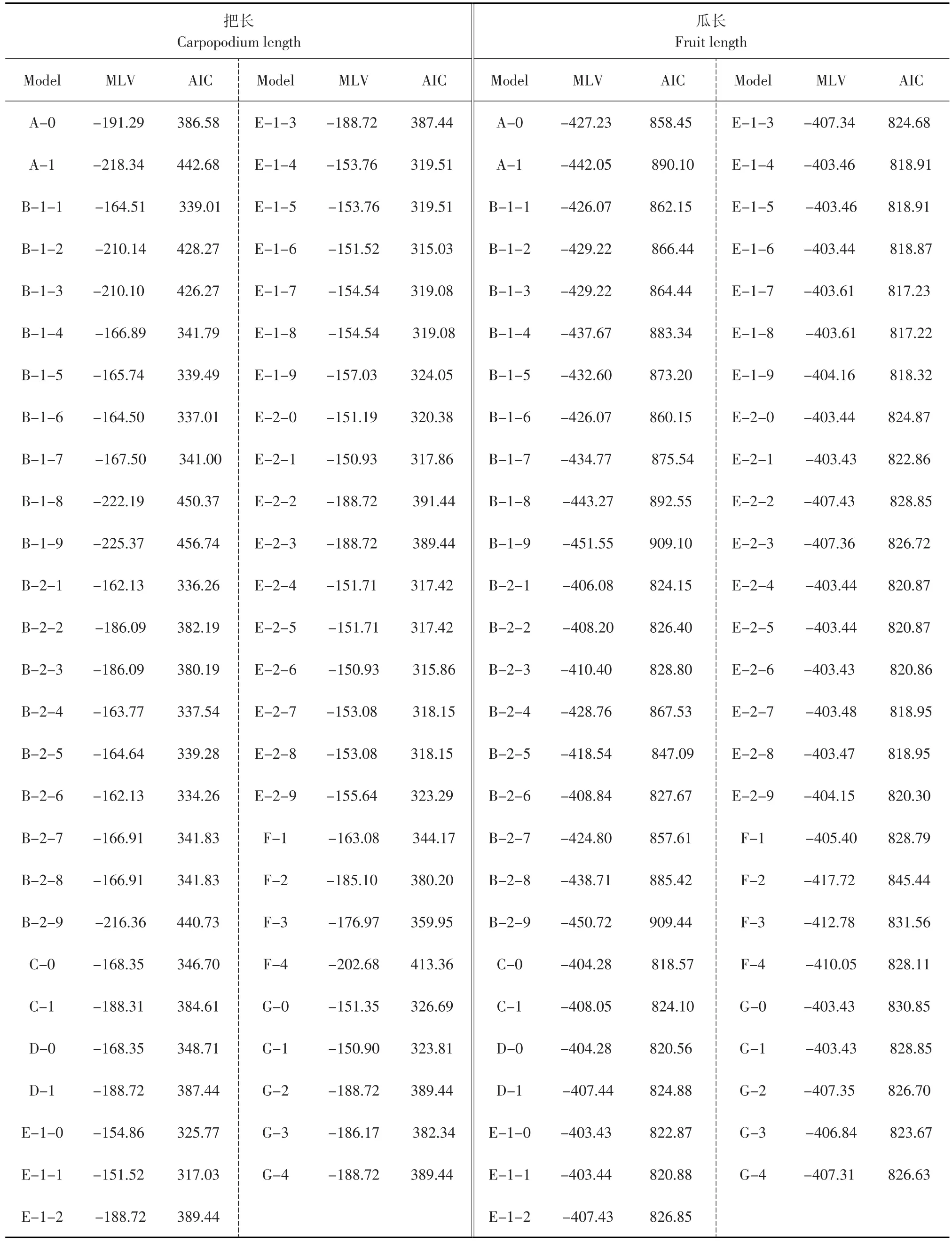
表3 黃瓜把長和瓜長各個遺傳模型的極大似然值(MLV)和AIC值Table 3 Estimated MLV and AIC of cucumber carpopodium length and fruit length in different genetic models
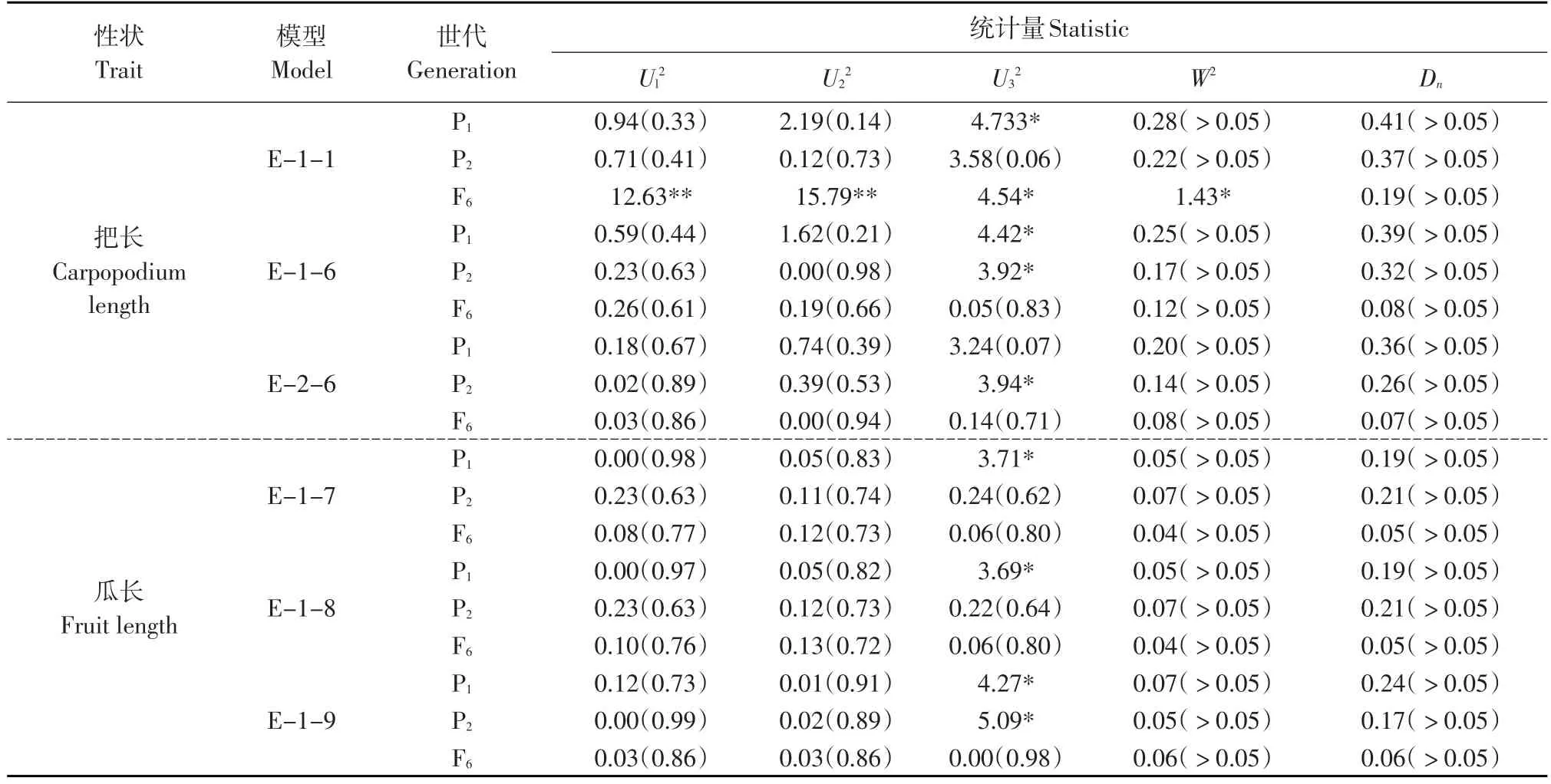
表4 備選模型的適合性測驗Table 4 Fitness test of candidate models
2.3 遺傳模型的參數估計
根據把長和瓜長最適模型的各個成分分布均值和權重,采用最小二乘法估計兩個最適模型的一階遺傳參數和二階遺傳參數(見表5)。由表5中的一階遺傳參數可知,把長兩對主基因之間表現為加性效應(d=0.36),同時兩對主基因之間還存在上位性效應(即加性×加性互作效應,i=0.69);瓜長兩對主基之因表現為加性效應與上位性效應相互綜合(i*=-2.49)。結果表明,黃瓜把長和瓜長的遺傳均受加性效應的影響。
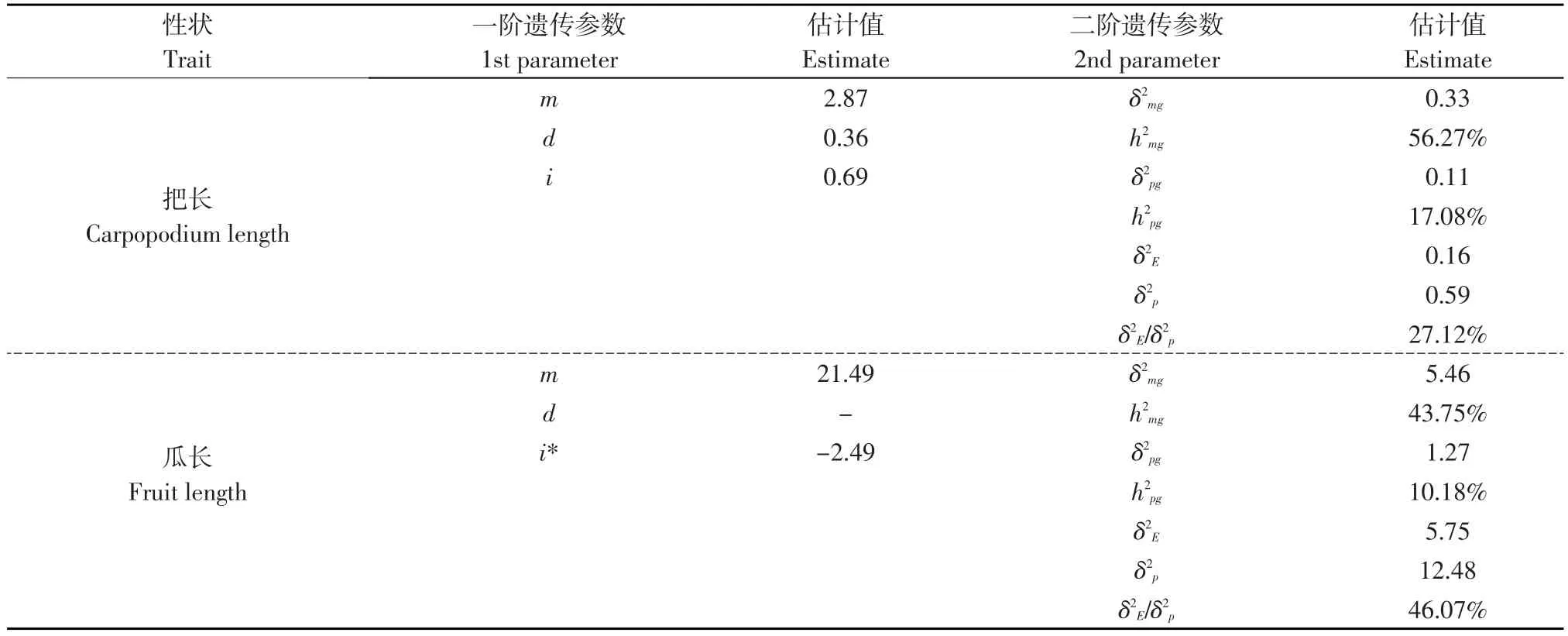
表5 黃瓜把長和瓜長性狀的遺傳參數估計Table 5 Estimation of genetic parameters of carpopodium length and fruit length in cucumber
由表5中的二階遺傳參數可知,把長和瓜長的主基因遺傳率分別為56.27%和43.75%,多基因遺傳率分別為17.08%和10.18%,兩個性狀的主基因遺傳率都明顯高于多基因遺傳率,表明黃瓜把長和瓜長兩個數量性狀的遺傳主要為主基因的遺傳。
另外,把長和瓜長的環境方差占表型方差的27.12%和46.07%,說明環境因素對把長和瓜長兩個性狀的影響較大,特別是瓜長,有相當大的非遺傳變異。
3 討 論
目前對黃瓜數量性狀的遺傳分析多采用多世代分離群體聯合(P1、P2、F1、F2、B1、B2)的分離分析法[16-18],而對于單世代分離群體(F2、BC、DH、RIL)的使用較少。本研究選用了單世代分離群體中的F6代重組自交系群體(Recombinant inbred line,RIL)進行遺傳分析。由于RIL群體是兩純合親本雜交F2代經連續多代自交并單粒傳代后形成的群體,在這種群體中每個株系都是純合的,所以利用RIL群體進行遺傳分析的最大優點在于能夠進行重復試驗以驗證試驗結果,而且RIL群體只存在加性效應以及與加性效應有關的互作效應(上位性效應)的變異,能夠檢測出來的遺傳模型要比F2代和幾種回交世代及多世代聯合群體簡單。但是RIL群體的構建是很費時的,在實際的研究中,人們常常使用自交6~7代的“準”RIL群體,因為到F6代后,單個基因座的雜合率大約只有3%,已經基本接近純合,利用這樣的“準”RIL群體進行數量性狀的遺傳分析也是可行的[19]。
前人對黃瓜把長和瓜長的遺傳分析進行了較為深入的研究,但結論大都不一致。顧興芳等研究認為,黃瓜把長的遺傳以加性效應為主,顯性效應較弱,狹義遺傳力高,能穩定遺傳,且受環境影響較小[11];馬德華等研究表明,把長的狹義遺傳力較低,易受環境影響,應注意高代選擇;瓜長的狹義遺傳力較高,高代選擇較為有效,受環境影響較小[12];張文新等認為,把長的狹義遺傳率較高,能夠在早世代進行選擇,選擇潛力較大,受環境影響較大,加性效應占主導地位[13];尹維娜等研究結果表明,把長和瓜長符合“加性-顯性”效應模型,以加性效應為主,兩者受環境影響較小[14];曹齊衛等認為把長的遺傳以顯性效應為主,受環境的影響較大;瓜長的遺傳加性效應為主,受環境影響較小[15];馬娟等認為把長受加性效應和顯性效應均較明顯,但加性效應更重要,環境對把長的遺傳影響較大[18]。本研究的結果表明,把長和瓜長的主基因遺傳相對較高,把長和瓜長均受加性效應的影響,在F6代中把長和瓜長的環境方差所占比例較高,表明環境對把長和瓜長的影響較大。本研究結果與前人研究存在一定的差異,原因很可能是試驗材料不一致或分析方法不同造成的。
4 結論
黃瓜把長的遺傳為兩對具有累加作用的連鎖主基因+加性多基因混合遺傳模型(E-2-6),兩對主基因存在加性效應,同時還存在上位性互作效應,主基因遺傳率為56.27%,多基因遺傳率為17.08%;黃瓜瓜長的遺傳為兩對具有重疊作用的主基因+加性多基因混合遺傳模型(E-1-8),兩對主基因表現為加性效應與上位性互作的綜合效應,主基因遺傳率為43.75%,多基因遺傳率為10.18%。黃瓜把長和瓜長遺傳主要為主基因的遺傳,環境對把長和瓜長的遺傳影響較大。
[1] 侯鋒.黃瓜[M].天津:天津科學技術出版社,1999:32-76.
[2] 蓋鈞鎰.植物數量性狀遺傳體系[M].北京:科學出版社,2003:120-166.
[3] 王金社,李海旺,趙團結,等.重組自交家系群體4對主基因加多基因混合遺傳模型分離分析方法的建立[J].作物學報,2010,36(2):191-201.
[4] 蓋鈞鎰.植物數量性狀遺傳體系的分離分析方法研究[J].遺傳,2005,27(1):130-136.
[5] Gai J Y,Wang J K.Identification and estimation of a QTL model and its effects[J].Theoretical and Applied Genetics,1998,97(7):1162-1168.
[6] Wang J,Podlic D W,Cooper M,et al.Power of the joint segregation analysis method for testing mixed major-geneand polygene inheritance models of quantitative traits[J].Theoretical andAppliedGenetics,2001,103(5):804-816.
[7] 張鵬,秦智偉,王麗莉,等.黃瓜果實彎曲性遺傳分析[J].東北農業大學學報,2010,41(1):29-33.
[8] 孫小鐳,王永強,王冰,等.黃瓜嫩果果皮葉綠素含量的遺傳[J].園藝學報,2004,31(3):327-331.
[9] 李博,秦智偉,周秀艷.黃瓜果肉顏色遺傳分析及SSR分子標記[J].東北農業大學學報,2010,41(12):21-25.
[10] 羅曉梅,司龍亭,尹維娜.黃瓜黃色線與瓜長比的主基因+多基因的遺傳分析[J].華北農學報,2008,23(2):88-91.
[11] 顧興芳,方秀娟,韓旭.黃瓜瓜把長度遺傳規律研究初報[J].中國蔬菜,1994,9(2):33-34.
[12] 馬德華,呂淑珍,沈文云.黃瓜主要品質性狀配合分析[J].華北農學報,1994(2):65-68.
[13] 張文新,魏毓棠,于紅茹,等.黃瓜數量性狀遺傳的研究初報[J].沈陽農業大學學報,2005,36(2):227-229.
[14] 尹維娜,司龍亭.黃瓜早期若干數量性狀遺傳分析[J].中國農學通報,2006,22(1):232-235.
[15] 曹齊衛,張衛華,王志峰,等.黃瓜果實性狀的Hayman遺傳分析[J].西南農業學報,2009,22(3):773-776.
[16] 嵇怡,徐強,陳學好.黃瓜株高性狀遺傳模型分析[J].揚州大學學報:農業與生命科學版,2009,30(3):75-79.
[17] 閆世江,張繼寧,劉潔.黃瓜苗期低溫弱光下MDA含量主基因-多基因聯合遺傳分析[J].西北植物學報,2009,29(3):458-462.
[18] 馬娟,司龍亭,田友.黃瓜把長主基因+多基因混合遺傳分析[J].西北農業學報,2010,19(10):161-165.
[19] 方宣鈞.作物DNA標記輔助育種[M].北京:科學出版社,2001:22-28.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
核科學與工程(2021年4期)2022-01-12 06:30:26
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
今日農業(2020年19期)2020-12-14 14:16:52
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
電子制作(2018年18期)2018-11-14 01:48:24
中學物理·高中(2016年12期)2017-04-22 11:53:03
山東工業技術(2016年15期)2016-12-01 05:31:22
光學精密工程(2016年6期)2016-11-07 09:07:19
