日本血吸蟲調寧蛋白樣蛋白P14基因疫苗對小鼠免疫保護作用研究*
2012-06-07 06:03:14唐小牛汪禮文汪學龍
中國人獸共患病學報 2012年6期
關鍵詞:小鼠
唐小牛,汪禮文,姚 勇,汪學龍
血吸蟲病是一種嚴重危害人體健康的人獸共患寄生蟲病。目前是一個全球性的重要公共衛生問題,主要流行于非洲、拉丁美洲和亞洲的70多個國家,尤其是熱帶和亞熱帶地區,受血吸蟲病威脅的人口達6億,感染者高達2億,其中2千萬患者有較重的臨床癥狀并伴有不同程度的勞動力喪失[1-2],每年約有50萬患者死于血吸蟲病。因此,世界衛生組織在70年代就提出了防治血吸蟲病的規化措施[3]。多年來,防治血吸蟲病的主要策略是以化療為主,輔以滅螺及環境衛生管理等綜合性措施。控制血吸蟲病最重要的措施仍然是應用安全有效的抗血吸蟲藥物吡喹酮,但僅僅依靠吡喹酮化療只能減少或降低人群血吸蟲感染率,而不能阻斷疫區血吸蟲病的傳播和再感染發生[4-5],所以化療需要反復進行[6-7]。而且該藥對哺乳期婦女并不安全,因此有大部分感染人群不能接受治療。因此繼續尋找新的血吸蟲病防治措施成為專家學者關注的焦點[8]。
開展的對血吸蟲病流行區人群化療后再感染狀況的一系列調查研究表明,人體對血吸蟲感染有獲得性免疫力[9-11],這為血吸蟲病疫苗的研制提供了理論依據。WHO認為應優先考慮發展人用血吸蟲病疫苗作為化療或其它防治措施的必要補充,發展血吸蟲病疫苗已成為各國專家學者的共識,是世界范圍內血吸蟲病防治科研工作的研究熱點[12-13]。
本研究前期試驗中,我們對日本血吸蟲P14鈣結合蛋白樣蛋白基因真核表達載體的構建、表達和鑒定[15]。在此用無內毒素大量質粒提取試劑盒大量制備了真核表達重組質粒pcDNA3.1(+)-SjP14、pcDNA3.1(+)-SjGST 和空質粒 pcDNA 3.1(+),免疫BALB/c小鼠,觀察小鼠的免疫保護力,探討pcDNA3.1(+)-SjP14作為日本血吸蟲疫苗的可能性。
1 材料與方法
1.1 實驗材料
1.1.1 陽性釘螺 日本血吸蟲陽性釘螺購自江西省血吸蟲病防治研究所。
1.1.2 實驗動物 體重18~22g、5~8w齡雌性BALB/c小鼠購買自安徽醫科大學實驗動物中心。
1.1.3 DNA 疫 苗 pcDNA3.1(+)-SjP14 和pcDNA3.1(+)-SjGST 由本室構建[14-15]。
1.1.4 主要試劑 無內毒素大量質粒提取試劑盒為北京百泰克生物技術公司產品。
1.2 方法
1.2.1 DNA疫苗的大量制備 -80℃取出帶重組質粒的E.coliBL21,于3mL/3μL Amp LB液體培養基中,37℃,225r/min振搖3h后,轉入100 mL/100Amp LB液體培養基中,37℃,225r/min振搖過夜,采用北京百泰克生物技術公司無內毒素大量質粒提取試劑盒,按試劑盒說明書大量提取質粒pcDNA3.1(+)-SjP14、pcDNA3.1(+)空質粒和pcDNA3.1(+)-SjGST。
1.2.2 免疫動物 將BALB/C小鼠隨機分為4組,每組10只。分別為:生理鹽水組給予100μL/鼠/次;空質粒組100μg/鼠/次;pcDNA3.1(+)-SjP14組,給予 100μg/鼠/次;pcDNA3.1(+)-SjP14+ pcDNA3.1(+)-SjGST 組分別給予100 μg/鼠/次。經肌肉注射,共3次,每次間隔2w。
1.2.3 尾蚴攻擊感染 末次免疫后2w經腹部皮膚感染日本血吸蟲尾蚴(30±1)條/鼠。
1.2.4 用 ELISA 方法檢測IgG1、IgG2a、總IgG 于首次免疫前1d、尾蚴攻擊前1d、解剖前1d分別經眶靜脈采血,分離血清,-20℃保存備用。用ELISA方法檢測IgG1、IgG2a、總IgG。
1.2.5 免疫保護效果觀察 尾蚴攻擊6w后解剖小鼠,行左心室-門靜脈灌注沖洗收集成蟲,計算減蟲率;留取肝臟,部分用5%KOH消化后在顯微鏡下行蟲卵計數,計算減卵率。部分肝臟用于組織病理學分析(HE染色法),觀察肝細胞變化及肉芽腫情況。
減蟲率:分別計數每只小鼠中血吸蟲成蟲數,計算每組小鼠的平均成蟲數和雌蟲數,按如下公式計算:


減卵率:稱取鼠肝總重量,按常規方法用5%的KOH消化1g肝組織,計算每克肝組織蟲卵數(EPG),按下述公式計算肝組織減卵率:
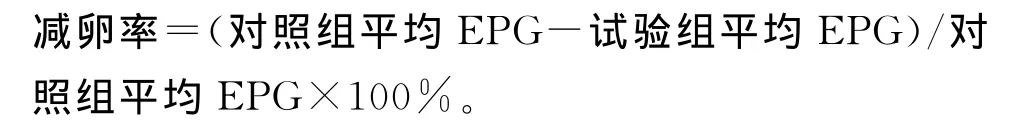
1.2.6 小鼠肝臟切片HE染色
1.2.7 統計學處理 數據錄入excel2003,采用spss11.0進行統計分析,計量資料以±s表示,多組間比較采用方差分析,以α=0.05為檢驗水準。
2 結 果
2.1 小鼠肝臟組織學的變化 大體解剖可見生理鹽水組肝臟表面可見彌漫性粟粒樣蟲卵結節分布,結節隆起密集,體積較大,空質粒組肝臟呈暗褐色,質硬,表面有溝紋,可見大量灰白色蟲卵結節。pcDNA3.1(+)-SjP14核酸疫苗組小鼠肝臟色澤略帶暗紅色,質地較軟,表面較光滑,隱約可見少量灰白色小點,蟲卵結節較少;SjP14+SjGST核酸疫苗聯用組,肝臟顏色呈鮮紅色,表面光滑,蟲卵結節少,質軟。組織切片鏡下顯示(圖1),生理鹽水組肝組織中可見大量的蟲卵肉芽腫形成,肉芽腫體積較大,周圍有大量淋巴細胞、嗜酸性細胞、巨噬細胞和漿細胞浸潤,大量的肝細胞變性壞死;空質粒組也見大量肉芽腫,周圍有大量淋巴細胞和巨噬細胞浸潤,但是肝細胞變性壞死度較重;pcDNA3.1(+)-SjP14組肝臟病較生理鹽水組明顯減輕,小鼠肝細胞混濁腫脹,偶見壞死的肝細胞,匯管區卵肉芽腫數量減少,炎性細胞浸潤主要以嗜酸性粒細胞和淋巴細胞為主,可見少量巨嗜細胞和漿細胞pcDNA3.1(+)-SjP14+SjGST病變程度最輕,肝小葉結構完整,蟲卵肉芽腫周圍炎癥反應輕,細胞浸潤不明顯,肉芽腫面積較小。
2.2 血清IgG1、IgG2a、總IgG的檢測 圖2、圖3、圖4分別顯示,免疫6w后,SjP14組小鼠血清抗體IgG1、IgG2a和總IgG均較對照組明顯升高,SjP14+SjGST聯合應用作用更顯著。

圖1 各組小鼠肝臟病理切片(HE染色)A.生理鹽水組 B.空質粒組 C.SjP14組 D.SjP14+SjGST組Fig.1 Photomicrographs of liver in each group(HE stain)A:NS group;B:pcDNA3.1(+)vector group;C:SjP14group;D:SjP14+SjGST group

圖2 各組小鼠血清特異性IgG1抗體水平Fig.2 Specific IgG1antibody in sera of mice

圖3 各組小鼠血清特異性IgG2a抗體水平Fig.3 Specific IgG2aantibody in sera of mice

圖4 各組小鼠血清特異性總IgG抗體水平Fig.4 Total IgG antibody in sera of mice
2.3 減蟲率和減卵率 由表1可見,SjP14核酸疫苗能明顯減少被尾蚴攻擊小鼠靜脈內成蟲數量,SjGST和SjP14核酸疫苗共同免疫則能提高SjP14疫苗抗尾蚴感染的作用,減蟲率分別達到44.6%(P<0.05),56.2%(P<0.01)。由表2可見,SjP14組和SjGST和SjP14聯合組小鼠肝臟內蟲卵數明顯低于生理鹽水和空質粒對照組(P<0.01),SjGST和SjP14聯合組減卵率由單用SjP14的61.5%提高到73.6%。
2.4 減卵率(表2)
表1 各組小鼠的成蟲檢獲情況(±s)Tab.1 Reduction rate of worm burden in each group(±s)

表1 各組小鼠的成蟲檢獲情況(±s)Tab.1 Reduction rate of worm burden in each group(±s)
與對照組比較:*P<0.05,**P<0.01Compared with control group:*P<0.05,**P<0.01
組別Group小鼠數(只)No.mice成蟲數(條/鼠)Worm burden減蟲率(%)Reduction rate(%)生理鹽水Saline 10 22.4±1.9空質粒Empty plasmid 10 21.8±1.8 SjP14 10 12.3±1.4 44.6*SjP14+SjGST 10 9.8±1.6 56.2**
表2 各組小鼠的蟲卵檢獲情況(±s)Tab.2 Reduction rate of liver egg in each group(±s)
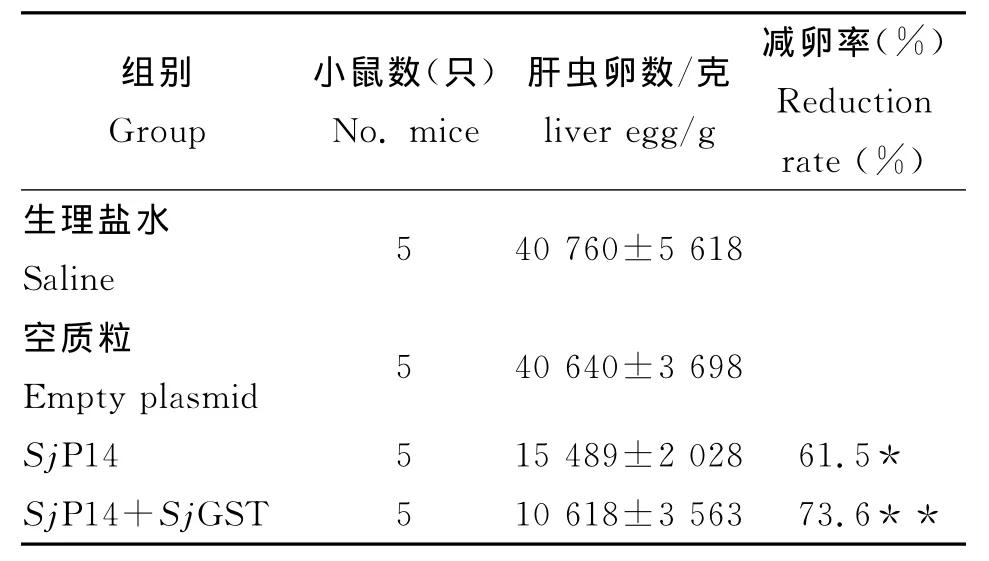
表2 各組小鼠的蟲卵檢獲情況(±s)Tab.2 Reduction rate of liver egg in each group(±s)
與對照組比較:*:P<0.01,**:P<0.01Compared with control groups:*:P<0.01,**:P<0.01
?
3 討 論
核酸疫苗能夠把一個或幾個編碼目標抗原的cDNA片段克隆到真核表達載體內,并表達出近似天然的抗原,從而誘導出較強的體液和細胞免疫應答。DNA疫苗進入體內,少量被宿主細胞攝取后進入細胞核,在載體啟動子的操縱下,抗原基因轉錄為mRNA,該mRNA又被胞漿粗面內質網上的核糖體翻譯生成細胞質內蛋白質。少量表達的蛋白在細胞內被水解酶降解成8~12個氨基酸的短肽,這些短肽含有不同的抗原表位,相關蛋白通過轉運進入內質網腔,與I型MHC分子的重鏈及β2-微球蛋白結合在一起,通過高爾基復合體轉移出細胞質,定位于細胞表面,被CD+4T細胞識別,刺激MHCI型限制分子的CD+4細胞毒性T細胞產生抗原特異性CTL,對表達外源基因的細胞發揮細胞毒性作用[16]。
已進行與日本血吸蟲疫苗相關的動物實驗,獲得了不少日本血吸蟲抗原,但這些抗原只能起到部分保護作用,它們仍然沒有達到致弱尾蚴保護作用的水平[12]。因此有必要繼續尋找新的疫苗抗原分子,促進血吸蟲疫苗研發的進程。
現已發現調寧蛋白具有很多生物學功能,包括(1)參與平滑肌收縮的調控;(2)參與細胞信號轉導;(3)維持細胞骨架;(4)參與血管病變及腫瘤的基因調節。不管脊椎動物還是無脊椎動物在其生命活動當中調寧蛋白都發揮著重要作用。調寧蛋白樣蛋白已經在許多無脊椎動物中被克隆和表征,包括最近報道的棘球絳蟲、盤尾絲蟲、秀麗小桿線蟲及日本血吸蟲等4種寄生蟲[17-19]。目前已知的日本血吸蟲疫苗的抗原免疫保護都沒有達到50%水平,而旋盤尾絲蟲的OV9M基因編碼的堿性調寧蛋白樣蛋白具有高度免疫原性,所以本實驗選擇日本血吸蟲的堿性調寧蛋白樣蛋白作為抗原,以觀察該抗原的免疫保護力。結果表明,pcDNA3.1(+)-SjP14DNA 疫苗能誘導小鼠產生顯著的抗日本血吸蟲感染的保護性免疫作用,pcDNA3.1(+)-SjP14 +SjGST 聯合應用其作用增強。與生理鹽水組和空質粒組相比較,其減蟲率和減卵率均有顯著性差異。小鼠肝臟切片HE染色顯示其肝細胞壞死、蟲卵量、炎性細胞的浸潤均明顯減輕。本實驗還發現特異性IgG1和IgG2a抗體水平均明顯升高,以初次免疫后12w時最明顯,攻擊感染后有較明顯下降,提示可能誘導了Thl/Tcl型和Th2/Tc2型混合型應答。通過以上分析提示SjP14有希望作為一種新的、有效的抗日本血吸蟲感染的候選疫苗分子。
[1]Chitsulo L,Engels D,Montresor A,et al.The global status of schistosomiasis and its control[J].Acta Trop,2000,77(1):41-51.DOI:10.1016/S0001-706X(00)00122-4
[2]Department of International Cooperation,Ministry of Health,China.Schistosomiasis fact sheet[R].World Health Organization Bulletin,2008,404(2):13-14.(in Chinese)衛生部國際合作司國際合作處.血吸蟲病的實況報道[R].世界衛生組織簡報,2008,404(2):13-14.
[3]Special Programme for Research and Training in Tropical Diseases.Tropical disease research:progress 1975-94:highlights 1993-94:twelfth programme report of the UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases(TDR)[M].Geneva:World Health Organization,1995:77.
[4]Chen MG.Schistosomiasis in the world and its control progress[J].Chin J Schisto Control,2002,14(2):81-83.(in Chinese)陳銘剛.世界血吸蟲病流行情況及防治進展[J].中國血吸蟲病防治雜志,2002,14(2):81-83.
[5]Jiang Y,Jiang ZJ.Overview on drug research for schistosomiasis control[J].Chin J Schisto Control,2001,13(1):59-60.(in Chinese)江艷,蔣作君.血吸蟲病防治藥物研究概況[J].中國血吸蟲病防治雜志,2001,13(l):59-60.
[6]Yuan HC,Zhang SJ,Liu ZL,et al.Research on epidemiological factors and optimization control strategy for schistosomiasis in lakeshore areas[J].Chin J Schisto Control,1995,7(4):1993-2001.(in Chinese)袁鴻昌,張紹基,劉志德,等.湖灘地區血吸蟲病流行因素與優化控制策略的研究[J].中國血吸蟲病防治雜志,1995,7(4),1993-2001
[7]Wu ZD,Zhang SJ,Pan BR,et asl.Reinfection withSchistosoma japonicumafter treatment with praziquantel in Poyang lake region,China[J].Southeast Asian J Trop Med Public Health,1994,25(1):163-169.
[8]Bergquist NR,Colley DG.Schistosomiasis vaccines:research to development[J].Parasitol Today,1998,14(3):99-104.DOI:10.1016/S0169-4758(97)01207-6
[9]Gryseels B.Human resistance toSchistosomainfection:Age or experiencef[J].Parasitol Today,1994,10(10):380-384.DOI:10.1016/0169-4758(94)90226-7
[10]Waine GJ,McManus DP.Schistosomiasis vaccine development--the current picture[J].Bioessay,1997,19 (5):435-443.DOI:10.1002/bies.950190511
[11]Zhang Z,Wu H,Chen S,et al.Association between IgE antibody against soluble egg antigen and resistance to reinfection withSchistosomiasis japonica[J].Trans R Soc Trop Med Hyg,1997,91(5):606-608.DOI:10.1016/S0035-9203(97)90047-X
[12]Bergquist NR,AI-Sherbiny M,Barakat R,et al.Blueprint for schistosomiasis vaccine development[J].Acta Trop,2002,82(2):183-192.DOI:10.1016/S0001-706X(02)00048-7
[13]Bergquist NR,Leonardo LR,Mitchell GF.Vaccine-linked chemotherapy:can schistosomiasis control benefit from an integrated approach[J].Trends Parasitol,2005,21 (3):112-117.DOI:10.1016/j.pt.2005.01.001
[14]Luo QL,Shen JL,Wang XL,et al.Immunodiagnosis of acute schistosomiasis using 26ku glutathione-S-transferase recombinant protein ofSchistosoma japonicum[J].Acta U Medicinalis Anhui,2005,40(6):491-494.(in Chinese)羅慶禮,沈繼龍,汪學龍,等.重組日本血吸蟲26ku谷胱甘肽-硫-轉移酶的表達、純化及其免疫特性分析用于急性血吸蟲病免疫診斷[J].安徽醫科大學學報,2005,40(6):491-494.
[15]Wang LW,Tang XN,Yao Y,et al.Construction,expression and identification of the eukaryotic expression vector carryingSchistosoma japonicumP14gene[J].Acta U Medicinalis Anhui,2011,46(4):321-324.(in Chinese)汪禮文,唐小牛,姚勇,等.日本血吸蟲P14鈣結合蛋白樣蛋白基因真核表達載體的構建、表達和鑒定[J].安徽醫科大學學報,2011,46(4):321-324.
[16]Simmonds RS,Sheaer MH,Kennedy RC.DNA vaccines-From principle to practice[J].Parasitol Today,1997,13(9):328-331.DOI:10.1016/S0169-4758(97)01106-X
[17]Martin RM,Gasser RB,Jones MK,et al.Identification and characterization of myophilin,a muscle-specific antigen ofE-chinococcus granulosus[J].Mol Biochem Parasitol,1995,70(1-2):139-148.DOI:10.1016/0166-6851(95)00020-2
[18]Irvine M,Huima T,Prince AM,et al.Identification and characterization of anOnchocerca volvuluscDNA clone encoding a highly immunogenic calponin-like protein[J].Mol Biochem Parasitol,1994,65(1):135-146.DOI:10.1016/0166-6851(94)90122-8
[19]Goetinck S,Waterston RH.TheCaenorhabditis elegansmuscle-affecting geneunc-87encodes a novel thin filament-associated protein[J].J Cell Biol,1994,127(1):79-93.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
