飼料淀粉類酶制劑的營養機理及應用現狀
2012-04-25 09:47:18蔣蘇蘇段紅偉
草業科學 2012年6期
蔣蘇蘇,段紅偉,于 鋒
(1.蘭州大學草地農業科技學院,甘肅 蘭州 730020;2.廣州威司特生物科技有限公司,廣東 廣州 510663)
谷物是動物日糧的重要組成成分,而谷物中的淀粉是單胃動物的主要能量來源,其供能占動物需求的60%~80%。一般幼齡動物消化道內淀粉酶和麥芽糖酶活性很低,不能充分消化日糧中的淀粉,大量飼喂淀粉可引起幼齡動物腹瀉,而成年動物對淀粉的表觀消化率也只有80%~95%。在飼料工業中,淀粉類酶制劑作為一種重要的消化性酶制劑能有效地補充動物在特定條件下內源酶的不足,起到輔助動物消化道酶系,降解淀粉成為易吸收的小分子的作用。
1 日糧中的淀粉
1.1淀粉結構 淀粉是由α-D-葡萄糖單元聚合而成的多糖。不同來源的淀粉,其形狀和大小等不同。根據葡萄糖聚合方式及淀粉形態的不同,淀粉分為直鏈淀粉和支鏈淀粉。直鏈淀粉是以葡萄糖單元經α-1,4糖苷鍵連接而成的直鏈狀淀粉,分子質量變化較大。直鏈淀粉還含有相當一部分的分支直鏈淀粉,分支點隔開很遠,物理性質基本上和直線型直鏈淀粉相同[1]。支鏈淀粉是一種高分枝的束狀結構,主鏈通過α-1,4 糖苷鍵連接而成。α-1,6 糖苷鍵約占總糖苷鍵4%~5%,相對分子量較大。直鏈淀粉難溶于水,溶液不穩定,凝成性強,支鏈淀粉則相反。玉米(Zeamays)和小麥(Triticumaestivum)約含直鏈淀粉28%,馬鈴薯(Solanumtuberosum)含21%,木薯(Manihotesculenta)含17%[2]。根據支鏈淀粉側鏈連接方式可將其分為:A型、B型和C型。A型淀粉支鏈較多且排列分散,B型淀粉支鏈較少,鏈與鏈之間較集中,C型淀粉介于兩者之間。體外研究發現,A型淀粉易于動物消化利用,多為谷物類淀粉,B型淀粉經熱處理后也難以消化,而C型淀粉消化性介于A型淀粉和B型淀粉之間,大部分為豆類淀粉。
1.2淀粉的分類 根據營養學分類,淀粉可分為:1)快速消化淀粉(Rapidly Digestible Starch,RDS),即經過加熱處理的糊化淀粉,這種淀粉易于消化;2)緩慢消化淀粉(Slowly Digestible Starch,SDS),即天然淀粉,可被緩慢地完全消化;3)抗性淀粉(Resistant Starch,RS)。Englyst等[3]將抗性淀粉分為3個亞類:1)物理上不可接近的淀粉,阻礙了淀粉酶對其的降解作用。如包裹在食糜內部,或位于部分破碎顆粒或整粒谷物內,谷物顆粒的大小和組成阻止了淀粉酶的作用;2)具有特殊結構或構成,對酶具有高度抗性,影響天然淀粉的消化;3)加工過程中形成的“回生”淀粉。“回生”淀粉又稱為老化淀粉,指直鏈淀粉和支鏈淀粉組成的淀粉在糊化時分散,但在冷卻時分散的分子重新發生聚合形成的結晶體,這種結晶體能抵抗酶的水解作用,也是最主要的抗性淀粉。Brown[4]研究發現,“回生”淀粉在動物體內未經消化可轉入腸道后段。Patil等[5]研究了不同的來源的飼料中淀粉的組成(表1)。
1.3淀粉的消化利用 在畜禽體內,淀粉的消化開始于口腔,唾液α-淀粉酶水解淀粉生成寡糖和少量葡萄糖,但作用有限。唾液淀粉酶的最適pH值是6.6~6.8,在食糜進入胃后,唾液淀粉酶的作用被胃酸破壞,胃內沒有消化淀粉的酶,但可以受到胃酸的微弱水解。另外,胃的機械作用能夠破壞大多數食物的結構,從而使淀粉酶接觸到原來被蛋白質網絡或細胞壁包埋的淀粉。淀粉的主要消化部位位于小腸,淀粉在消化液中的α-淀粉酶作用下,被分解為麥芽糖、麥芽三糖及α-極限糊精。大分子的α-極限糊精又在小腸分泌的低聚α-極限糊精酶作用下分解為二糖,而后,在胰腺和腸粘膜中的麥芽糖酶、蔗糖酶和乳糖酶作用下最終分解為單糖。Rogel等[6]研究發現,空腸α-淀粉酶活性最強,大部分淀粉可在空腸內消化。
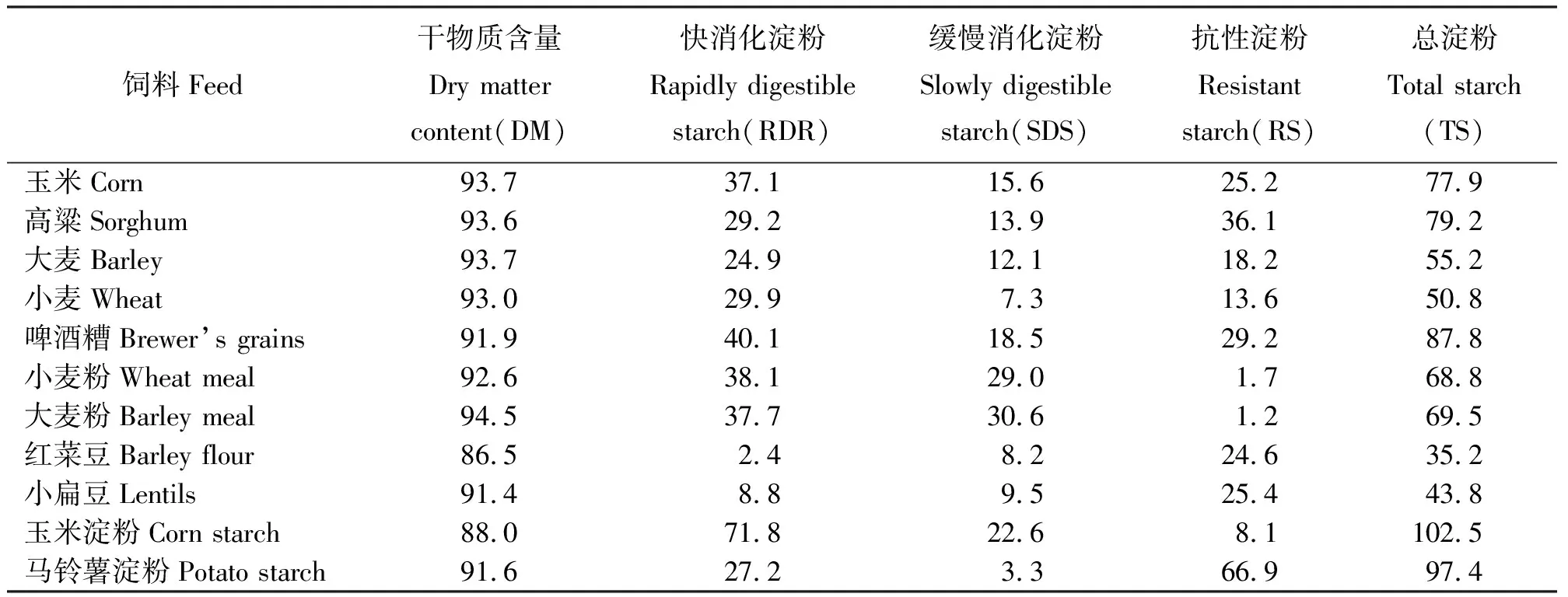
表1 不同飼料的淀粉組成[5]
1.4淀粉性質對畜禽消化利用的影響
1.4.1淀粉顆粒大小、直/支鏈淀粉比及來源 由于品種及生長階段的差異,淀粉的顆粒大小、直/支鏈淀粉的含量與比率及淀粉結晶程度也存在差異。體外法研究發現大淀粉顆粒水解速度較慢,說明淀粉顆粒的大小影響淀粉水解速度,進而影響酶對淀粉的降解。Zobel[7]研究發現,飼料淀粉中直鏈淀粉含量越高其消化性能越差。Regmi等[8]也證實,體外消化率低且直鏈淀粉含量較高的淀粉,可增強豬的腸道內營養物質的流動及微生物發酵,選擇性地促進后腸末端雙歧桿菌的生長。Zhou等[9]研究發現,玉米中的直/支鏈淀粉比是影響鴨子的真代謝能的主要因素,同時也可作評定玉米生物學效價的重要指標。戴球仲等[10]證實,用直/支鏈淀粉比不同的日糧飼喂黃羽肉仔雞,試雞的腹脂率和食后0.5 h血液中葡萄糖和胰島素含量受到顯著影響。Weuding等[11]研究發現,不同的飼料淀粉在肉雞體內消化率不同,且在特定消化部位消化率也存在差異,淀粉主要消化部位為肉雞的空腸。
1.4.2飼料加工處理 飼料在高溫、高壓、高水分及高機械力作用下,淀粉分子間氫鍵斷裂,產生水化作用,淀粉由生淀粉(β-淀粉)轉化為熟淀粉(α-淀粉),即完成淀粉的糊化作用。在飼料加工過程中,淀粉的本質并沒有發生變化,只是淀粉部分發生糊化,或產生少量的老化淀粉。淀粉的水解程度隨糊化度增加。胡友軍等[12]研究發現,糊化淀粉使仔豬淀粉表觀消化率平均提高了6.7%,血糖反應提前且峰值持續較長。許多學者研究了不同飼料加工處理方法及其對飼料淀粉消化部位、消化速率和消化程度的影響。目前,有大量關于飼料加工處理對飼料淀粉消化部位和消化率影響的研究。Hale[13]總結了18種飼料加工方法,并將其分為干處理和濕處理兩大類,同時研究發現由于處理方法的不同飼料淀粉的消化出現差異。Muir等[14]發現,到達后腸中的淀粉量與谷物中抗性淀粉的含量及加工方式有關,即抗性淀粉含量越高,如未經蒸煮碾磨等方式處理的谷物,過小腸而未能被消化的淀粉顆粒越多。Tricarico等[15]研究發現,不同加工方式的玉米中添加α-淀粉酶(添加量580或160 DU· kg-1,DM)可提高干物質攝入量和平均體質量日增加量。每千克干物質添加580 DU的α-淀粉酶后,背最長肌的面積最大。Carre等[16]將淀粉來源為小麥、玉米和豌豆(Pisumsativum)的飼料,經粉碎(過2 mm篩)后蒸汽制粒處理,發現淀粉消化率明顯增大(表2)。
2 淀粉酶
2.1淀粉酶類型和來源 催化淀粉水解的酶統稱為淀粉酶(Amylase),按照底物水解方式分為4類:α-淀粉酶(α-Amylase)、β-淀粉酶(β-Amylase)、糖化酶(Glucoamylase)和脫支酶(Debranchins enzyme)。微生物中產淀粉酶的菌種非常廣泛,通過發酵等工藝,不同的微生物可以產生不同的淀粉酶(表3)。
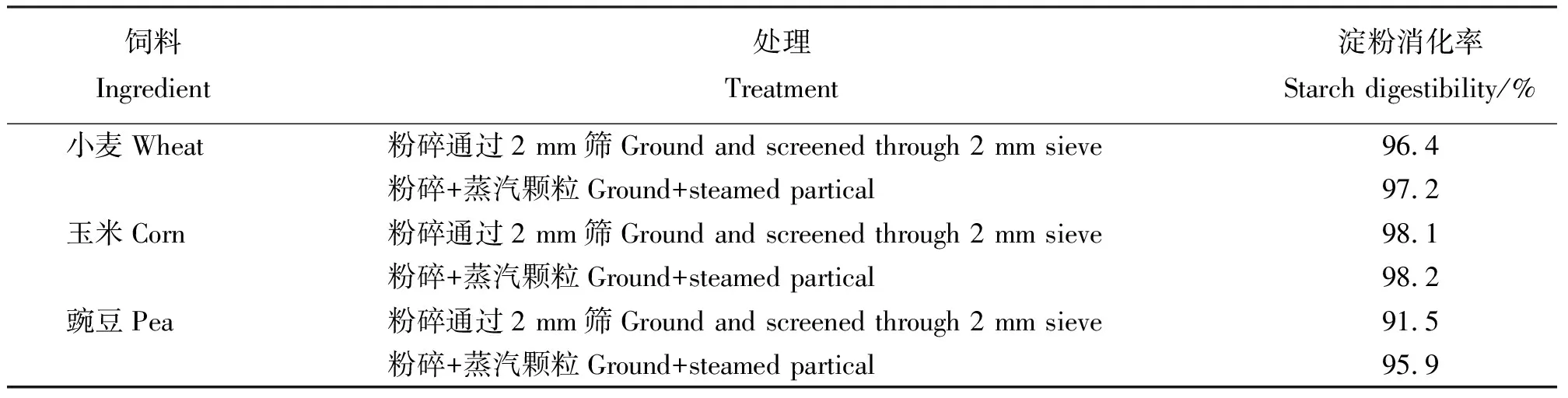
表2 飼料加工處理對成年肉雞淀粉消化率的影響[16]

表3 微生物來源的淀粉酶類型[17]
2.2淀粉酶的作用機理及其特性
2.2.1α-淀粉酶 α-淀粉酶是一種內切淀粉酶,以隨機切斷2-1,4-糖苷鍵的方式作用于淀粉分子,對分支點的α-1,6-糖苷鍵無效,且不能越過分支點水解α-1,4-糖苷鍵,水解后葡萄糖還原末端C1的構型為α-型,故稱為α-淀粉酶。直鏈淀粉子在α-淀粉酶作用下,可迅速水解為麥芽糖、麥芽三糖和較大的寡糖,再進一步水解為麥芽糖和葡萄糖。α-淀粉酶水解淀粉的最初階段,淀粉分子降解,引起黏性迅速降低,工業上稱為液化作用。
酶是有“活性”的蛋白質,其催化活力和穩定性受到很多因素的影響,其中最主要的是溫度和pH值。酶都有最適的溫度和pH值范圍,在此范圍內酶活力最好。不同來源的淀粉酶也有其最適溫度和pH值范圍。芽孢桿菌和地衣芽孢桿菌所產的α-淀粉酶耐熱性最高,其最適溫度為90~92 ℃,屬耐高溫型淀粉酶;枯草桿菌所產的α-淀粉酶,最適反應溫度為70 ℃,屬中溫型淀粉酶;真菌來源的α-淀粉酶,最適溫度為55 ℃。通常淀粉酶在pH值5.5~8.0時穩定,在pH值4.0以下時易失活,最適pH值為5.0~6.0。除黑曲霉產生的α-淀粉酶耐酸(pH值2.0~3.0),其他酶一般不耐酸,pH值低于4.5時易失活。純化的α-淀粉酶在50 ℃以上容易失活,但在大量Ca2+存在或淀粉的水解產物糊精存在時酶對熱的穩定性會增強[17]。
2.2.2β-淀粉酶 β-淀粉酶為外切型淀粉酶,作用于淀粉非還原末端的α-1,4-糖苷鍵,依次切下一個麥芽糖單位,對分支點的α-1,6-糖苷鍵無效,且不能跨過分支點水解,生成麥芽糖及大分子的β-界限糊精,產物由α-型變為β-型麥芽糖,故稱為β-淀粉酶。β-淀粉酶的活性中心為巰基(-SH),在氧化劑、重金屬離子及巰基試劑作用下可失活,但還原性試劑如谷胱甘肽、半胱氨酸能提高其對熱的穩定性。β-淀粉酶和α-淀粉酶作用最適pH值范圍基本相同,一般在5.0~6.5,但β-淀粉酶的熱穩定性明顯低于α-淀粉酶,70 ℃以上均會失活。不同來源的β-淀粉酶差異較大,細菌β-淀粉酶最適作用溫度低于50 ℃。此酶的主要產酶菌種有曲霉、根霉和內孢霉。
2.2.3葡萄糖淀粉酶 葡萄糖淀粉酶也稱為α-1,4葡萄糖水解酶,俗稱糖化酶。糖化酶對淀粉的水解作用與β-淀粉酶相似,從淀粉非還原端開始催化α-1,4糖苷鍵斷裂,水解產物只有葡萄糖。糖化酶專一性較差,水解α-1,4糖苷鍵的同時,還能以較低的速度催化α-1,6和α-1,3糖苷鍵的斷裂。不同來源的糖化酶最適溫度和pH值存在一定差異。黑曲霉的最適溫度為55~60 ℃,最適pH值為3.5~5.5;根霉的最適溫度為50~55 ℃,最適pH值為4.5~5.5;擬內孢霉最適溫度為50 ℃,最適pH值為4.8~5.0。以上這3種是主要的產酶菌種,但其所產糖化酶各有差異。黑曲霉的酶活穩定,能在較高的溫度和較低pH值范圍內使用,但產物中混有少量葡萄糖基轉移酶,影響了葡萄糖的最終產率,根霉和擬內孢霉所產酶不含雜酶。根霉由于不適合深層發酵,工業上很難實現大量生產。目前,我國生產上最常用的是黑曲霉及其突變株發酵培養。
2.2.4脫支酶 脫支酶能高效、專一的切開支鏈淀粉分支點的α-1,6糖苷鍵,從而切下支鏈淀粉的整個側鏈,形成直鏈淀粉。α-1,6糖苷鍵是淀粉水解利用率低的主要障礙因素。所以,脫支酶和其他淀粉酶的配合使用,能有效提高淀粉利用率。根據水解底物專一性不同,可將脫支酶分為異淀粉酶(Isoamylase)、普魯蘭酶(Pullulanase)、低聚葡萄糖苷酶(Oligo-1-6-glucosidase)和淀粉-1,6葡萄糖苷酶(Amylo-1-6-glucosidase)。
目前,較為廣泛使用的脫支酶為異淀粉酶和普魯蘭酶,這兩種酶對底物的水解方式各有差異。異淀粉酶能水解分支點的α-1,6糖苷鍵,但對直鏈淀粉中的α-1,6糖苷鍵無作用。其最小作用底物是63-麥芽三糖基麥芽四糖,當分支點葡萄糖殘基少于3個時不能發揮作用,對普魯蘭糖(Pullulan)無作用。普魯蘭糖也稱為茁酶多糖,是由麥芽三糖兩端以α-1,6-糖苷鍵連接其他麥芽三糖形成的高分子多糖。普魯蘭酶可以作用于直/支鏈淀粉中的α-1,6糖苷鍵,水解所要求的底物分子最小。
2.3淀粉酶應用研究 研究表明,在動物日糧中添加淀粉酶,可提高動物的生產性能,改善飼料轉化率。Gracia等[18]研究發現日糧中添加α-淀粉酶,可使肉雞體質量日增加量提高9.4%(P<0.05),飼料轉化率提高4.2%(P<0.05)。蔣正宇等[19]也發現α-淀粉酶的添加,提高了肉雞前期和后期體質量增加量,且體質量日增加量與添加劑量呈線性關系(P<0.01),但飼料轉化率未受影響。Pavlovic等[20]證實,與基礎日糧組相比,實驗組降低代謝能5%的同時添加α-淀粉酶,體質量日增加量較基礎日糧組高出1.95%,料重比下降0.54%,α-淀粉酶的添加有利于動物健康和提高經濟效益。Onderci等[21]研究發現,添加產α-淀粉酶的大腸桿菌到仔雞飲水中,顯著提高了21日齡仔雞的體質量日增加量(P<0.05)、飼料轉化率(P<0.01)及飼料的表觀消化率(P<0.01),但對粗蛋白及脂肪的消化率無顯著影響;產α-淀粉酶的大腸桿菌的添加相對減少了胰腺的質量,但對肝臟質量及空腸、回腸等長度無顯著影響(P>0.05)。可能是由于禽類消化道較短,內源淀粉酶分泌有限,使得淀粉酶對食糜中淀粉的消化程度受到限制,而添加淀粉酶可彌補內源淀粉酶的不足,提高對淀粉及能量的利用率,進而改善了其生產性能。金立志[22]進行了添加含淀粉酶的復合酶試驗,發現該復合酶使肉雞平均生長速度提高3.5%,飼料轉化率提高2.5%~13.9%。劉慶華等[23]研究以玉米-豆粕型為基礎日糧,分別添加0.15 kg·t-1淀粉酶、0.20 kg·t-1復合酶和0.20 kg·t-1復合酶+0.15 kg·t-1淀粉酶,發現單獨添加淀粉酶能顯著提高蛋種雞對飼料中淀粉、干物質、有機物和能量的利用率與日糧代謝能(P<0.05);單獨或與復合酶聯合添加淀粉酶均能顯著改善蛋種雞的產蛋率與料蛋比(P<0.05),與單獨添加相比,兩者聯合添加有進一步提高養分利用率的趨勢,但效果并不明顯。以上研究表明,日糧中淀粉酶或含有淀粉酶的復合酶,均可改善動物對養分利用率。淀粉酶的使用對單胃動物淀粉的消化有重要的影響。Tricarico等[15]研究表明,粗飼料[紫花苜蓿(Medicagosativa)、棉仔餅]中添加α-淀粉酶對肉牛胴體品質有顯著影響(P<0.05),棉籽餅添加組提高28~112 d的體質量日增加量(ADG),并且在其他階段也趨向提高ADG。在兩種來源的粗飼料中,添加α-淀粉酶可提高肉牛背最長肌的眼肌面積(LM)。Klingerman等[24]也證實了外源α-淀粉酶的添加能改善泌乳期奶牛的生產性能。另外,飼料中添加的外源酶對動物機體內源酶的影響一直是人們關注的焦點。外源性酶與畜禽內源性消化酶活性的關系較為復雜,受到動物品種和飼糧類型等多方面的影響。據推測,在飼糧中添加外源性酶制劑,可增加在腸道中進一步分解或吸收的養分量,從而刺激機體消化系統的發育[25]。奚剛等[26]研究玉米-豆粕型飼糧中添加葡萄糖淀粉酶和中性蛋白酶,結果表明,37和67日齡絲毛烏骨雞的小腸胰蛋白酶、總蛋白水解酶活性明顯提高,但對內源性α-淀粉酶和脂肪酶活性無顯著影響。楊全明[27]證實了在斷奶仔豬料中添加淀粉酶和蛋白酶,對仔豬小腸腸道中淀粉酶、蛋白酶的活性沒有影響,但總蛋白酶活性提高。
3 展望
隨著飼料工業及養殖業的發展,飼料資源短缺的問題也日漸突出,而玉米作為動物主要的能量來源,需求量持續增大,進口量大幅增加,其價格從2010年上半年上升態勢迅猛。淀粉酶制劑的添加可提高谷物類原料的利用率,改善動物生產性能等點。但由于動物自身能產生少量的淀粉酶,使得人們忽視了對淀粉酶的添加利用。再者,幾種淀粉類酶制劑組合使用在現在飼料中的研究也越來越多,也是淀粉類酶制劑在飼料工業中應用發展的一個方向。淀粉酶制劑在飼料中的使用仍然處于發展階段,對淀粉酶的研究內容在不斷擴大,如淀粉酶與其他酶制劑的協同關系、對于動物內環境的影響,以及最經濟及最有效的添加量等方面。總之,淀粉酶類酶制劑發展有著廣闊的前景。
[1]Takeda Y,Hizukuri S,Juliano B O.Purification and structure of amylase from rice starch [J].Carbohydrate Research,1986,148(22):299-308.
[2]劉亞偉.玉米淀粉生產及轉化技術[M].北京:化學工業出版社,2003:159-169.
[3]Englyst H N,Kingman S M,Cummings J H.Classification and measurement of nutritionally important starch fractions[J].European Journal of Clinical Nutrition,1992,46(suppl 2):33-50.
[4]Brown I.Complex carbohydrates resistant starch[J].Nutrition Reviews,1996,54(11):115-119.
[5]Patil A R,Murray S M,Hussein H S,etal.Quantification of starch fractions in selected starchy feed ingredients[J].Animal Science,1988,76(suppl.):168.
[6]Rogel A M,Armison E F,Bryden W L,etal.The digestion of wheat starch in broiler chickens[J].Australian Journal of Agricltural Research,1987,38:639-649.
[7]Zobel H F.Molecular to granules:A comprehensive starch review[J].Starch,1988,40(2):44-50.
[8]Regmi P R,Metzler-Zebeli B U,G?nzle M,etal.Starch with high amylose content and low in vitro digestibility increases intestinal nutrient flow and microbial fermentation and selectively promotes bifidobacteria in pigs[J].Journal of Nutrition,2011,141(7):1273-1280.
[9]Zhou Z,Wan H F,Li Y.The influence of the amylopectin/amylose ratio in samples of corn on the true metabolizable energy value for ducks[J].Animal Feed Science and Technology,2010,157:99-103.
[10]戴求仲,劉紹偉,李湘,等.飼糧直/支鏈淀粉比對黃羽肉雞血液生化指標和后腸微生物菌群的影響[J].動物營養學報,2010,22(4):904-910.
[11]Weuding R E,Veldman W A G,Veen P J,etal.Strach digestion rate in the small intestine of broiler chickens differs between feed stuffs[J].Journal of Nutrition,2001,131:2327-2335.
[12]胡友軍,周安國,楊風,等.糊化淀粉對斷奶仔豬消化率和血糖水平的影響[J].動物營養學報,2003,15(2):33-35.
[13]Hale W H.Influence of processing on the utilization of grains (starch) by ruminants[J].Journal of Animal Science,1973,37(4):1075-1080.
[14]Muir J G,Birkett A,Brown I,etal.Food processing and maize variety affects amounts of starch escaping digestion in the small intes tine[J].America Journal of Clinical Nutrition,1995,61:82-89.
[15]Tricarico J M,Johnston J D,Dawsona K A.Dietary supplementation of ruminant diets with anAspergillusoryzaeα-amylase[J].Animal Science,2007,85:802-811.
[16]Carre B,Escartin R,Melcion J P,etal.Effect of pelleting and assoclations with maize or wheat on the nutritive value of smooth pea (Pisumsativum) seeds in adult cockerels[J].British Poultry Science, 1987,28(2):219-29.
[17]鄭惠平.飼料工業酶技術[M].北京:化學工業出版社,2008:58.
[18]Gracia M I,Aranibar M J,Lazaro R,etal.Alpha-amylase supplementation of broiler diets based on corn[J].Poultry Science,2003,82(3):436-442.
[19]蔣正宇,周巖民,王恬,等.外源α-淀粉酶對肉雞生產性能的影響[J].家畜生態學報,2007,28(4):13-16.
[20]Pavlovic M,Resanovic R,Markovic R,etal.Effct of diet with normal or reduced energy and alpha-amylase supplemetation on growth in broilers[A].In:Slovenia.19 International Science Symposium on Nutrition of Domestic Animals “Zadravec-Erjavec Days”[C].Slovenia:Kmetijsko gozdarska zbornica Slovenije,Murska sobota,2010:205-209.
[21]Onderci M,Sahin N,Sahin K,etal.Efficacy of supplementation of A-amylase-producing bacterial culture on the performance,nutrient use,and gut morphology of broiler chickens fed a corn-based diet[J].Poultry Science,2006,85(3):505-510.
[22]金立志.飼用酶制劑應用與研究現狀及未來發展[J].飼料廣角,2002,16:5-11.
[23]劉慶華,徐秋良,李夢云,等.添加淀粉酶與復合酶對蛋種雞生產性能及養分利用率的影響[J].西北農林科技大學學報,2010,38(8):15-20.
[24]Klingerman C M,Hu W,McDonell E E,etal.An evaluation of exogenous enzymes with amylolytic activity for dairy cows[J].Journal of Dairy Science,2009,92(3):1050-1059.
[25]Owsley W F,Orr D E,Tribble L F.Effects of age and diet on the development of the pancreas and synthesis and secretion of pancreatic enzyme in the young pig[J].Animal Science,1986,63:497-504.
[26]奚剛,許梓榮,錢利純,等.添加外源酶對豬、雞內源酶活性的影響[J].中國獸醫學報,1999,19(3):286-289.
[27]楊全明.仔豬消化道酶和組織器官生長發育規律的研究[D].北京:中國農業大學,1999:34-55.
