產低溫脂肪酶菌株的篩選、純化及其部分酶學性質研究
2012-04-13 12:30:44王春雨遲乃玉張慶芳
中國釀造 2012年11期
王春雨,遲乃玉*,張慶芳
(大連大學 生命科學與技術學院,遼寧 大連 116622)
低溫脂肪酶具有高效、耐熱、作用周期短等優勢,在應用上具有中高溫脂肪酶無法取代的優越性,因此低溫脂肪酶廣泛的應用在食品、輕紡、皮革、香料、化妝品、洗滌劑、有機合成、醫藥等領域上。國外研究者對其中幾種典型的脂肪酶如來源于Candida rugosa[6]、Candida antarcticaA[7]、CandidaantarcticaB[8]、Rhizomucor miechei[9]、Thermomyces lanuginosus[10]的脂肪酶的應用進行了詳細的綜述,這些脂肪酶在油脂加工和食品工業、壽星藥物合成、生物能源、醫藥和化妝品等行業顯示出良好的前景。相比于國外,國內目前只有作為洗滌劑的堿性脂肪酶,而酯化或轉酯化用的脂肪酶還是空白。
本文從大連大黑山和渤海灣采樣,篩選產低溫脂肪酶的菌株,并對其產酶條件進行研究。
1 材料與方法
1.1 土樣來源
渤海灣海水和大連大黑山土樣。
1.2 培養基
Luria-Bertani(LB)培養基[11]:胰蛋白胨1.0%,酵母提取物0.5%,NaCl 1.0%,瓊脂1.5%,pH7.0。
初篩培養基:蛋白胨1.0%,酵母粉0.5%,NaCl 0.5%,CaCl20.02%,Tween80 1.0%,pH7.2。
發酵培養基:酵母膏0.5%,(NH4)2SO40.5%,KH2PO40.2%,NaCl 0.3%,MgSO4·7H2O 0.05%,橄欖油2%(v/v),pH7.0。
在教學中,建立關系的前提是師生之間發生交流和交往,但現在的許多課堂,只是單向的教師講授、學生接收。教師的注意力主要在“教”,沒有注意到在“教”的過程中與學生發生交流和交往,建立某種超越客觀知識的情感關系。我試圖強調的是,在“教”的過程“之中”發生了什么。當前,我們沒有很好地區別什么是教師關注自己的教學本身,什么是關注教學“之中”發生的事情。這個區別不是特別明顯,但同時這個區別又是巨大的。
牛肉膏蛋白凍培養基:牛肉膏0.3%,蛋白胨1.0%,Na-Cl 0.5%,瓊脂1.8%,pH7.0。
1.3 產低溫脂肪酶菌株的初篩
樣品稀釋至10-7后,接種于初篩培養基平板上,20℃培養4d。產脂肪酶的菌株能分解Tween80在菌落周圍形成白色沉淀圈。將產生白色沉淀圈的菌株活化后,分別接種于初篩培養基平板上,記錄菌落直徑與沉淀圈直徑之比。
1.4 產低溫脂肪酶菌株的復篩
將待測菌株活化后,接種于種子液中,20℃、160r/min培養12h,轉接于100mL發酵液中,20℃、160r/min培養2d,測其酶活。
1.5 產低溫脂肪酶菌株的紫外誘變
選出一株菌落直徑與沉淀圈直徑之比和酶活最大的菌株,將其制成種子液,在20℃、160r/min培養12h。把菌液稀釋到適當梯度,置于磁力攪拌器上,在紫外燈的功率15W,照射的距離為30cm的條件下,照射不同時間進行紫外誘變。分別吸取不同照射時間的稀釋液均勻涂布到初篩培養基上,裝入黑布袋中,倒置于恒溫箱中,在20℃培養3d,取出計算菌落數,并觀察白色沉淀圈。
1.6 脂肪酶活性測定方法
取3mL 50mmol/L磷酸鹽緩沖液(pH值為7.0)和1mL橄欖油,放入水浴恒溫磁力攪拌37℃中預熱5min,加入0.1mL酶溶液,磁力攪拌10min,立即加入8mL笨,繼續攪拌2min,終止反應,同時萃取生成的脂肪酸。將溶液轉移至離心管中,4000r/min離心10min,有機相和水相分離澄清。取上層有機相4mL于小錐形瓶中,加入1mL顯色劑,磁力攪拌上攪拌3min,4000r/min離心10min,取上層含有脂肪酸銅的苯溶液,用分光光度計在波長710nm處測OD值。
由脂肪酶活力定義得活力計算公式:脂肪酶活力(單位/mL)=CV/TV’,公式中C脂肪酶的濃度(μmol/mL);V脂肪酸/苯溶液的體積(mL);T作用時間(min);V’酶液的用量(mL)。
1.7 產脂肪酶菌株的鑒定
1.7.1 形態學、生理生化特征
產脂肪酶菌株的形態學觀察、生理生化實驗參照文獻[12]方法進行。
1.7.2 16SrDNA序列測定和分析
以菌株CZWOO1總DNA為模板,利用真細菌的16SrDNA通用引物進行PCR,產物經瓊脂糖凝膠電泳分析,得到一條1.5bp左右的條帶(見圖1)。以Seq Forward、Seq Internal和16SSeqF1為引物進行DNA測序。序列分析結果表明,菌株CZWOO1的16SrDNA序列得到有效擴增,其大小為1429bp。進入NCBI主頁,應用BLAST程序與數據庫中的已有細菌16S rDNA序列進行相似性比較分析。利用ClustalX1.83進行匹配比對,然后用MEGA5.05軟件構建系統發育樹。
圖1 3%瓊脂糖凝膠電泳Fig.1 3% Agarose gel electrophoresis
2 結果
2.1 菌種篩選
從59株來自大連大黑山土樣和渤海灣海水的細菌中篩選到10株產低溫脂肪酶的菌株,其中以菌株007的直徑比最大且酶活最高,其酶作用溫度最低(25℃)。對該菌株進行紫外誘變,當誘變130s時,致死率達到90.7%,得到一株直徑比和酶活明顯高于原菌株的菌株,命名為CZWOO1。
2.2 菌種鑒定
2.2.1 形態學、生理生化特征
菌株CZWOO1在LB培養上20℃培養3d,菌落乳白色、圓形、隆起、不透明、表面光滑、濕潤、邊緣整齊,直徑約0.9cm。菌體革蘭氏陰性、短桿狀。常規生理生化特征見表1。
2.2.2 分子生物學鑒定
序列分析結果表明,菌株CZWOO1的16SrDNA序列全長為1429bp。將序列輸入GenBank進行BLAST相似度比較,下載相似度序列,利用ClustalX進行多序列匹配比對,通過MEGA5.05軟件計算出序列的系統進化距離,采用Neighbor-Joining構建系統進化樹(見圖2)。菌株CZWOO1與Bacillusthuringiensis(NR 043403.1)親緣關系最近,同源性達到100%,因此菌株CZWOO1屬于芽孢桿菌屬中的蘇云金芽孢桿菌,命名為BacillusthuringiensisCZWOO1。
2.3 脂肪酶的分離純化及二級結構預測

表1 菌株CZWOO1 生理生化特征Table 1 Physiological and biochemical characteristics of the strain CZWOO1
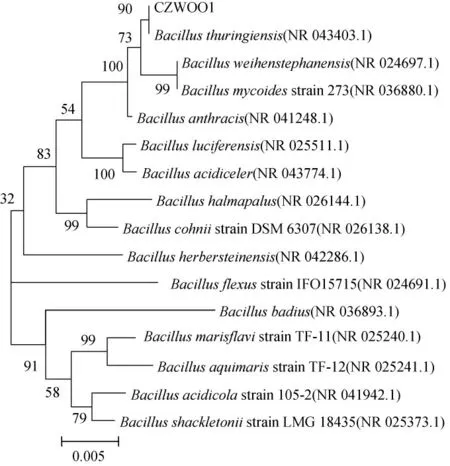
圖2 菌株CZWOO1基因系統發育進化樹Fig.2 Phylogenetic trees for 16S rDNA sequence of CZWOO1
蘇云金芽孢桿菌CZW001脂肪酶發酵上清經65%硫酸銨沉淀,用0.05mol/LTis-Hcl緩沖液(pH8.2)溶解,并用相同的緩沖液4℃透析12h獲得粗酶液。經截留分子量為10ku的中空纖維柱超濾濃縮,將濃縮液加到預先用Tis-Hcl緩沖液(0.05mol/L pH8.2)平衡好的DEAE-纖維素-52層析柱(Φ1.6×30cm)上,用250mL 0~1mol/L的NaCl線性梯度洗脫。利用平板定性檢測脂肪酶活性,收集含脂肪酶各管,測定其酶活,合并含酶活各管。將樣品過Sephadex G-100(Φ1.6×50cm)柱,用0.05mol/L Tis-Hcl緩沖液(pH8.2)洗脫,收集有酶活性部分,結果見圖3。蛋白曲線呈現一個小峰和一個大峰,而低溫脂肪酶酶組分主要集中在大峰內,25管~45管范圍內的蛋白峰酶活性比較高,說明Sephadex G-100凝膠柱已經除去了一部分高分子量和低分子量的蛋白,將目的酶蛋白同其他雜蛋白分離開來。
蘇云金芽孢桿菌CZW001脂肪酶發酵上清液經硫酸銨沉淀、超濾離心、DEAE-纖維素-52離子交換層析和Sephadex G-100凝膠過濾層析等純化步驟,脂肪酶純化倍數為75.5倍,活性回收率為37.6%。其中DEAE-纖維素-52離子交換層析效果最佳,僅此一步就純化了56倍(純化效果見表2)。SDS-PAGE凝膠電泳[9]顯示純化后的脂肪酶為單一條帶(見圖4),分子量約為40ku。純化倍數為75.5倍,活性回收率為37.6%,跑12%SDS-PAGE電泳為單一條帶(見圖4),分子量約為40ku。
對多次分離純化的低溫脂肪酶進行圓二色譜分析,見圖5。用Jascow32軟件分析及Yang-chen公式計算其二級結構成分:α-螺旋10.6%,β-折疊40.1%,β-轉角18.8%,無規則卷曲30.5%。
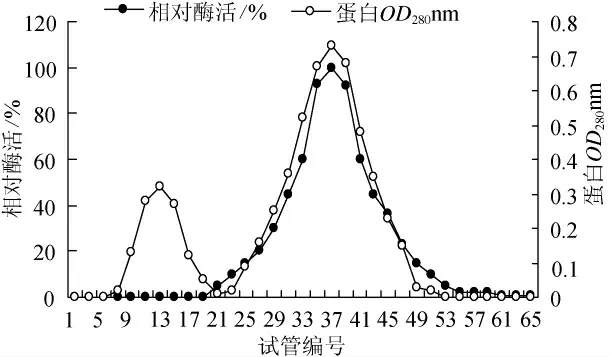
圖3 Sephadex G-100 凝膠過濾Fig.3 Gel filtration chromatogram of Sephadex G-100
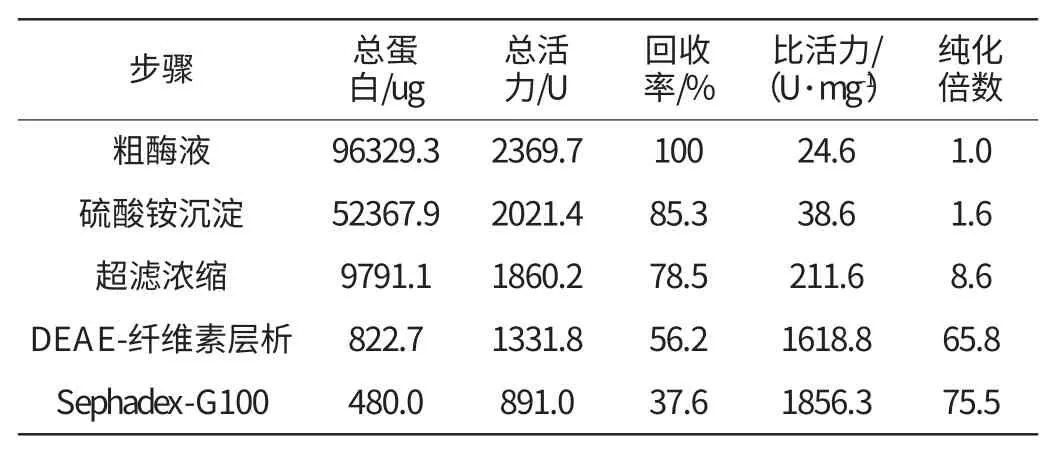
表2 脂肪酶的純化Table 2 Purification of Bacillus thuringiensis CZWOO1 lipase
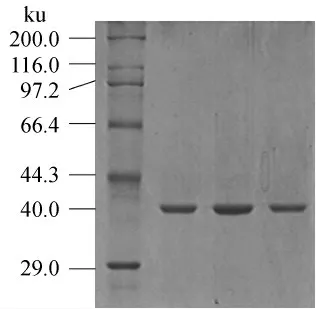
圖4 12%SDS-PAGE電泳圖Fig.4 12% SDS-PAGE of enzyme solution
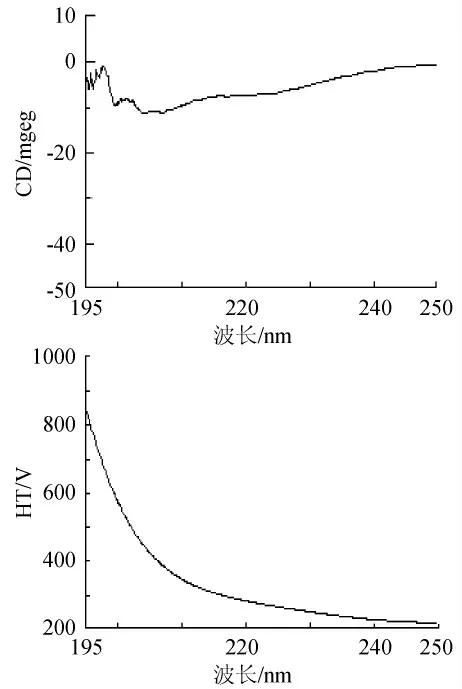
圖5 低溫脂肪酶圓二色譜圖Fig.5 The CD spectrum of cold-active amylase
2.4 脂肪酶性質
2.4.1 pH值對CZWOO1脂肪酶活力的影響
在30℃、不同pH值條件測定菌株CZWOO1脂肪酶活力,結果見圖6。該酶的適宜作用pH值范圍在7~9,最適pH值為8。
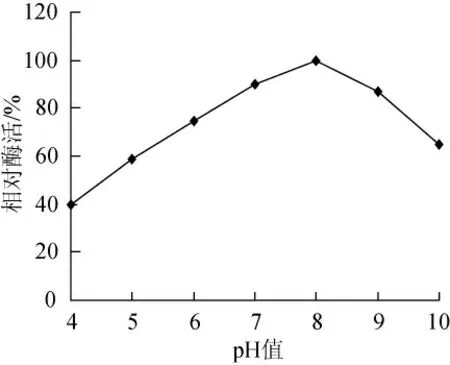
圖6 p H值對CZWOO1脂肪酶活力的影響Fig.6 Effects of pH value on lipase activity of CZWOO1
2.4.2 溫度對CZWOO1脂肪酶活力的影響

圖7 溫度對CZWOO1脂肪酶活力的影響Fig.7 Effects of temperature on lipase activity of CZWOO1
在pH8.0條件下,測定10℃~45℃溫度范圍酶活力與溫度的關系,結果見圖7。CZWOO1脂肪酶活力在25℃達到最高,在10℃是仍具有較高的酶活,相對酶活達到38%。
2.4.3 溫度對CZWOO1脂肪酶穩定性的影響
將酶液在不同溫度中保溫30min,在25℃、pH8.0條件下,測定CZWOO1脂肪酶殘余活力。結果見圖8,CZWOO1脂肪酶穩定性較差,在60℃處理30min,酶活損失達70%。
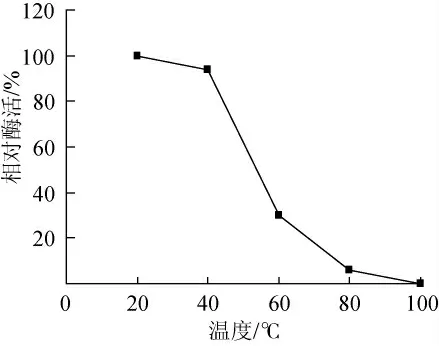
圖8 溫度對CZWOO1脂肪酶穩定性的影響Fig.8 Effects of temperature on lipase stability of CZWOO1
3 討論
菌株CZWOO1為革蘭氏陰性短桿菌,兼性厭氧,不能水解淀粉、明膠、纖維素和尿素,能水解油脂,使阿拉伯糖、木糖和甘露糖產酸,吲哚和檸檬酸鹽試驗陰性。從菌株CZWOO1基因系統發育進化樹看,菌株CZWOO1與Bacillus thuringiensis(NR 043403.1)親緣關系最近,同源性達到100%,因此菌株CZWOO1屬于芽孢桿菌屬中的蘇云金芽孢桿菌,命名為BacillusthuringiensisCZWOO1。
蘇云金芽孢桿菌CZW001脂肪酶發酵上清液經硫酸銨沉淀、透析、超濾離心、DEAE-纖維素-52離子交換層析和Sephadex G-100凝膠過濾層析得到電泳純的脂肪酶,跑12%SDS-PAGE電泳為單一條帶,分子量約為40ku。對純化后的低溫脂肪酶進行圓二色譜分析,結果表明,其二級結構成分:α-螺旋10.6%,β-折疊40.1%,β-轉角18.8%,無規則卷曲30.5%。對該脂肪酶二級結構的預測給其適冷機制的研究奠定基礎。
對BacillusthuringiensisCZW001分泌的脂肪酶的初步研究表明,該脂肪酶的適宜作用pH值范圍在7~9,最適pH值為8,最適作用溫度為25℃,對熱敏感,60℃處理30min僅殘留30%酶活性。該低溫脂肪酶在低溫下具有良好的催化活性,作用pH值偏堿性(pH7~9)等特點,應在環境治理、洗滌業等領域有著良好的應用前景,并且其對熱敏感等特點,從而避免高溫滅活對食品品質產生的破壞,在食品工業也有良好的應用潛力。
[1]QU YEN D T,SCHMIDT-DANNERT C,SCHMID RD.High-level expression of alipasefromBacillusthermocatenulatusBT L2 inPichia pastorisand some properties of the recombinant lipase[J].Prot Expr Pur,2003,28(1):102-110.
[2]林學政,邊 際,何培青.極地微生物低溫適應性的分子機制[J].極地研究,2003,15(1):75-82.
[3]RASHID N,SHIMADA Y,EZAKIS,et al.Low-temperature lipase from psychrotrophicPseudomonassp strain KB700A[J].Appl Environ Microb,2001,67(9):4064-4069.
[4]LEEHK,AHNMJ,KWAK SH,et al.Purification and characterization of cold active lipase from psychrotrophicAeromonassp.LPB 4[J].J Microbiol,2003,41(1):22-27.
[5]TRIPATHIM K,ROY U,JINWAL U K,et al.Cloning,sequencing and structural features of a novelStreptococcuslipase[J].Enzyme Microb Technol,2004,34(5):437-445.
[6]BENJAMIN S,PANDEY A.Candida rugosalipases:molecular biology and versatility in biotechnology[J].Yeast,1998,14:1069-1087.
[7]DOMINGUEZ DE M P,CARBONI-OERLEMANS C,TUIN B,et al.Biotechnological applications ofCandida antarcticalipase A:stateof the art[J].J Mol Catal B:Enzym,2005,37:36-46.
[8]ANDERSON E M,LAARSSON K M,KIRK O.One biocatalyst many applications:theuseofCandidaB-lipaseinorganic synthesis[J].Biocatal Biotransf,1998,16:181-204.
[9]RODRIGUE S R C,FERNANDEZ-LAFUENTE R.Lipase fromRhizomucor mieheias an industrial biocatalyst in chemical process[J].J Mol Catal B:Enzyme,2010.
[10]FERNANDEZ-LAFUENTER.Lipase fromThermomyces lanuginosus:usesand prospectasan industrial biocatalyst[J].J Mol Catal B:Enzym,2009,62(3):197-212.
[11]SAMBROOK J,FRISH E F,MANIATIS T.Molecular.Molecular Cloning:Alaboratory Manual[M].Second Edition.New York:Cold Spring Harbor Laboratory Press,1989.
[12]東秀珠,蔡妙英.常見細菌系統鑒定手冊[M].北京:科學出版社,2001.
