米氏冰草種群種子產(chǎn)量與種子雨的時間動態(tài)
2012-03-13 05:37:24林海俊王宏光金曉明劉及東閆志勇
草業(yè)科學(xué) 2012年11期
關(guān)鍵詞:植物
林海俊,王宏光,金曉明,劉及東,閆志勇
(1.呼倫貝爾市教育局,內(nèi)蒙古 呼倫貝爾 021008; 2.呼倫貝爾學(xué)院生命科學(xué)與化學(xué)學(xué)院,內(nèi)蒙古 呼倫貝爾 021008)
種子或果實靠自身的重力和外界力量(如風(fēng)力等)散布到地表的過程,被形象地稱為種子雨(Seed Rain)[1]。以往對種子雨的研究多集中在木本植物[2-6],而對草本植物種子雨的研究相對較少,沙地植物種子雨的研究則更少[7-11]。種子雨是土壤種子庫的主要來源,直接影響種群和群落的組成、結(jié)構(gòu)和動態(tài)[12],是群落更新發(fā)展的重要環(huán)節(jié)。了解植物種子雨的時間格局,對于深入研究植物種群的更新、發(fā)展動態(tài)及植被恢復(fù)具有重要科學(xué)價值。國內(nèi)外有關(guān)種子雨散布時空動態(tài)的研究已有大量報道[13-19]。米氏冰草(Agropyronmichnoi)別名為根莖冰草,屬禾本科小麥族的多年生草本植物,多分布于歐亞大陸溫帶草原區(qū),是呼倫貝爾草地的鄉(xiāng)土種,具有很高的飼用價值。另外,在長期的自然選擇過程中,米氏冰草形成了對沙地環(huán)境較強的適應(yīng)性,既可以成為沙地植被次生演替的先鋒種,又可以通過無性繁殖形成沙地植物群落的優(yōu)勢種。米氏冰草具有很強的抗旱性,在年降水量為200~380 mm地區(qū)生長良好。因此,在沙漠化治理實踐中,米氏冰草是兼顧生態(tài)效益和經(jīng)濟效益的重要固沙植物。然而,有關(guān)米氏冰草在沙地生境中有性繁殖體散布特征及規(guī)律方面的研究鮮見報道。本研究以呼倫貝爾沙漠化草地米氏冰草種群為對象,開展種子雨時空動態(tài)及有性繁殖器官特性的研究,旨在為沙化草地植被的恢復(fù)和管理提供科學(xué)依據(jù)。
1 材料與方法
1.1研究區(qū)概況 研究區(qū)位于內(nèi)蒙古呼倫貝爾市陳巴爾虎旗完工鎮(zhèn)境內(nèi),地理位置為11°44′75″~118°54′76″ E,49°08′87″~49°11′48″ N,海拔588.4~618.0 m,具溫帶大陸干旱半干旱過渡氣候特征。年均氣溫-1.5 ℃,降水量230~300 mm(70%集中于夏秋季),而蒸發(fā)量為降水量的4~6倍;年均風(fēng)速4.5 m·s-1,最大風(fēng)速為20 m·s-1,全年8級以上大風(fēng)日數(shù)平均30 d以上。地帶性植被以貝加爾針茅(Stipabaicalensis)和羊草(Leymuschinensis)為優(yōu)勢種群;沙漠化草地的植被主要以草本植物種為主,其中有米氏冰草、東北牡蒿(Artemisiamanshurica)、扁蓿豆(Melissitusruthenius)、狗尾草(Setariaviridis)、百里香(Thymusvulgaris)、豬毛菜(Salsolafoliosa)、沙蓬(Agriophyllumsquarrosum)等。群落中還零星分布著少量的灌木,如差不嘎蒿(A.halodendron)和小葉錦雞兒(Caraganamicrophylla)[20]等。
研究樣地在呼倫貝爾市陳巴爾虎旗呼和諾爾蘇木境內(nèi)的沙漠化草地內(nèi),為2005年進行人工補播的米氏冰草草地,經(jīng)過了5年的自然恢復(fù)。其中,樣地1(以下簡稱半固定沙地)是以米氏冰草為單優(yōu)勢種群的半固定沙地,位于沙丘上部,植被蓋度為35%~45%;樣地2(以下簡稱固定沙地)是以米氏冰草為單優(yōu)勢種群的固定沙地,位于沙丘下部,植被蓋度為65%~75%。兩種樣地的面積均大于2 000 m2。
1.2研究方法 種子產(chǎn)量用單位面積內(nèi)植物有性繁殖體所形成的種子數(shù)量來表示;種子雨強度用植物有性繁殖后代散布到周邊環(huán)境中的單位面積內(nèi)的數(shù)量來表示。試驗期間每天的氣候因子的測定數(shù)據(jù)由呼倫貝爾市陳巴爾虎旗氣象局提供(表1)。
1.2.1種子雨強度 米氏冰草種子雨的采集時間選擇在其成熟期進行,即2009年8月2日-9月27日。每隔5 d收集一次種子。種子雨收集器為直徑0.2 m、高0.08 m、底部具鐵紗網(wǎng)、上方無封閉的圓筒型竹籮筐(籮筐的底面積為0.031 4 m2)。將種子收集器隨機擺放在半固定沙地和固定沙地樣地內(nèi),每個樣地10個重復(fù),共20個種子收集器,在種子收集器外圍用一次性筷子將其固定。每次取種子時,先將收集器內(nèi)的枯枝落葉和其他植物的種子等雜物分離出去,收集近5 d內(nèi)的米氏冰草種子,放入塑封袋中帶回實驗室并進行計數(shù)。
種子雨強度=種子雨數(shù)量/收集器底部面積。
1.2.2有性繁殖體性狀及種子產(chǎn)量 在半固定沙地和固定沙地樣地分別按草方格大小(草方格為用稻草建植的網(wǎng)格狀人工沙障,由于地形和人工作業(yè)的差異,草方格面積存在一定誤差,一般為1.0~2.5 m2),隨機選取10個面積不等的樣方,計數(shù)每個樣方內(nèi)出現(xiàn)的穗數(shù)。同時在半固定沙地和固定沙地樣地分別隨機抽取30個穗部,計數(shù)每穗上的種子數(shù)量。同時對隨機抽取的穗部,測定其穗長、穗寬、每穗上的小穗數(shù)量等。然后計算出種子產(chǎn)量,即:
種子產(chǎn)量=樣方內(nèi)出現(xiàn)的穗數(shù)量×平均每穗上的種子數(shù)量/樣方面積×100%。

表1 米氏冰草種子雨期間的氣候因子數(shù)據(jù)Table 1 Data of climatic factors during seed rain of Agropyron michnoi
1.2.3數(shù)據(jù)統(tǒng)計與分析 用Excel和SPSS軟件對數(shù)據(jù)進行統(tǒng)計分析,用平均數(shù)(M)代表樣本各性狀整體水平特征,用標(biāo)準(zhǔn)差(SD)反映樣本的變異度,對不同樣地的相應(yīng)指標(biāo)進行方差分析(One-Way Anova),比較不同生境之間的種子產(chǎn)量及有性繁殖體的差異顯著性。
種子雨期間,按每隔5 d的取樣日期,分別取這5 d的平均風(fēng)速、最大風(fēng)速、平均氣溫和平均降水量為變量,與種子雨強度進行相關(guān)性分析,再以其中相關(guān)性最大的參數(shù)為變化,與種子雨強度進行回歸分析。
2 結(jié)果與分析
2.1種子雨時間動態(tài) 研究結(jié)果顯示(圖1),半固定沙地和固定沙地的米氏冰草種群種子散布期基本一致,均為8月8日-9月27日。在種子散布期間,半固定沙地和固定沙地的米氏冰草種群的種子雨強度的波動變化也基本相似。種子雨的最大高峰期均出現(xiàn)在9月7日。另外,在8月13日、8月28日和9月22日時,半固定沙地出現(xiàn)了3次小高峰值,而固定沙地只呈小幅度的波動。各個時期的米氏冰草種子雨強度,基本上為半固定沙地高于固定沙地。在半固定沙地和固定沙地,米氏冰草種子雨總強度分別為3 158.2和1 146.5粒·m-2,半固定沙地的種子雨總強度是固定沙地的2.8倍。

圖1 不同生境米氏冰草種群種子雨時間動態(tài)Fig.1 Temporal dynamics of seed rain of Agropyron michnoi populations in different habitats
2.2種子產(chǎn)量與有性繁殖體的關(guān)系 對米氏冰草種群有性繁殖體的研究顯示(表2),其穗長、穗寬、單穗質(zhì)量及每穗上的小穗數(shù)半固定沙地上分別是固定沙地的1.3倍、1.5倍、1.5倍和1.2倍,且均表現(xiàn)出顯著差異(P<0.05或P<0.001),表明沙地生境的變化對米氏冰草穗部形態(tài)特征產(chǎn)生一定影響。米氏冰草的種子百粒重為半固定沙地比固定沙地的高1.17倍,且存在顯著差異(P<0.05),而每穗上的種子數(shù)及單位面積種子產(chǎn)量,在兩樣地之間卻無顯著差異,表明半固定沙地的米氏冰草種子比固定沙地的大,但在單株種子數(shù)量及單位面積種子數(shù)量上均相同。
在半固定沙地和固定沙地,米氏冰草種群種子產(chǎn)量分別為10 961和11 502粒·m-2(表2)。其中,在半固定沙地和固定沙地,米氏冰草種子雨數(shù)量分別占其種子產(chǎn)量的29%和10%,表明米氏冰草種子成熟后,種子雨進入土壤種子庫的數(shù)量因生境的變化而有所差異。
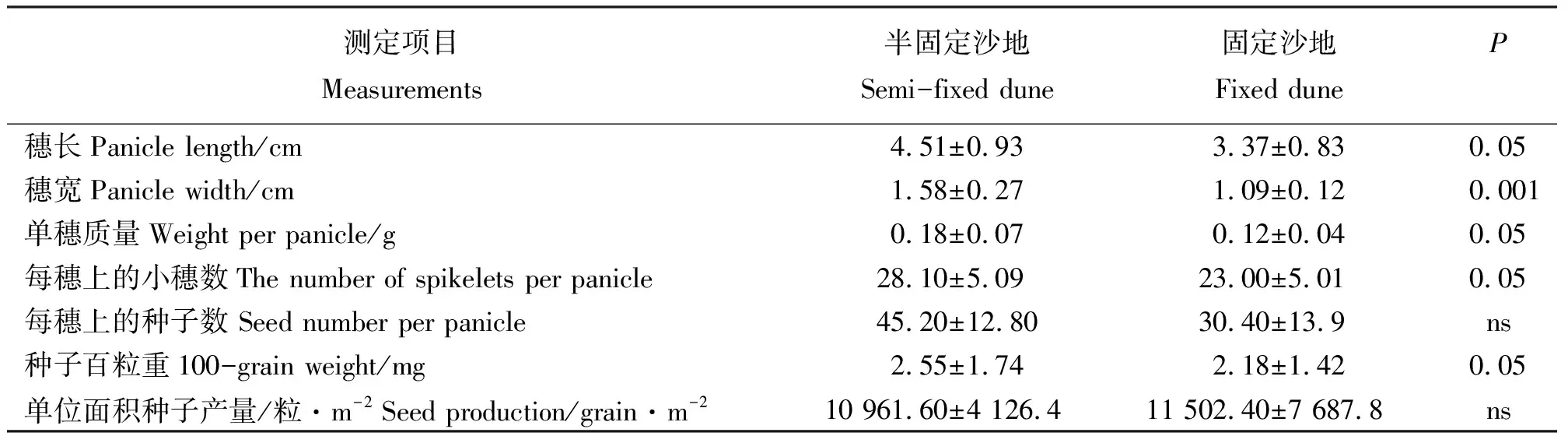
表2 不同生境米氏冰草有性繁殖體特征Table 2 Characteristics of sexual propagules of Agropyron michnoi populations in different habitats
2.3種子雨強度與最大風(fēng)速的關(guān)系 以種子雨期間每5 d的最大風(fēng)速為變量,與種子雨強度進行回歸分析(圖2),半固定沙地種子雨強度與風(fēng)速之間以Y=-572.7+270.8X規(guī)律變化,固定沙地則以Y=-253.7+111.7X規(guī)律變化,且其回歸方程均呈現(xiàn)出正態(tài)分布,顯著性差異均小于0.001。
3 討論與結(jié)論
種子擴散指種子離開母株的運動,是種子尋求萌發(fā)的最佳時間和空間的過程[14]。由于種子通過擴散可以減少幼苗與母株的競爭、幼苗之間的競爭以及母株附近的密度制約性死亡。遠(yuǎn)離母株的種子與母株附近的種子相比,出苗成活率較高[6,21]。因此,米氏冰草種子雨強度的大小及其分布格局對其在沙地的成功定居具有重要意義。種子擴散是發(fā)生在多個空間和時間尺度上的最基本的生物過程,包括種子雨的擴散及種子雨落地后的第2次擴散。以往的研究[22]表明,植物種之間的種子產(chǎn)量千差萬別,不同種群之間的種子雨密度相差較大;由于環(huán)境條件如土壤深度、地形、土壤養(yǎng)分等的差異,相同植物種群內(nèi)部不同個體之間的種子雨也有差異。本研究表明,在呼倫貝爾沙化草地,不同生境對米氏冰草種群的種子雨散布時期無影響,但卻對其種子雨強度有顯著影響。前者可能是由于兩樣地距離較近,氣候條件基本相似所致,但較大尺度上的生境變化是否對米氏冰草的種子雨散布時期有影響,尚不清楚;后者可能是兩樣地的土壤養(yǎng)分和水分條件存在一定差異所致。但米氏冰草采取何種調(diào)整有性繁殖分配的策略,是否與生境土壤養(yǎng)分和水分條件的變化有關(guān),還有待于進一步研究。
植物繁殖分配與植物的生長、植物的生活史、植物所處的演替狀態(tài)以及植物的生存環(huán)境均有關(guān)系[23-27]。米氏冰草是無性系植物,既可通過地下根莖的生長進行無性繁殖,也可通過形成有性繁殖體進行種子繁殖,其穗部為有性繁殖器官[25]。
許多干旱地區(qū)的植物,如四合木(Tetraenamongolia)種群在環(huán)境惡劣的灘地,其生殖值、地上年凈生物量的生殖分配比生境條件好的丘地要高[28]。本研究結(jié)果也與此相似,即米氏冰草種群的穗長、穗寬、單穗質(zhì)量、每穗上的小穗數(shù)及種子百粒重等指標(biāo),在環(huán)境條件相對惡劣的半固定沙地均顯著高于固定沙地。另外,質(zhì)量較大的種子更有利于埋入土壤,有效地獲取土壤中的水分,促進其萌發(fā)和幼苗的生長。
在本研究中,雖然不同生境米氏冰草種子單穗上的種子數(shù)量與單位面積的種子產(chǎn)量均無顯著差異,但整個種子雨期間收集的累計種子數(shù)量只占其種子產(chǎn)量的10%~33%,且半固定沙地比固定沙地高3倍。這可能是因為隨著沙地植被的恢復(fù),動物數(shù)量也有所增加,致使大部分成熟脫落的種子被動物所采食。Gashwiller[29]發(fā)現(xiàn),鼠類的破壞造成道格拉斯杉樹(Pseudotsugamenziesii)種子的損失率達63%,僅有12%的種子在第2年萌發(fā)。沈澤昊等[30]對山地常綠落葉闊葉混交林種子雨的地形格局研究發(fā)現(xiàn),地形對植被空間格局異質(zhì)性的影響機制之一是作用于種子雨的空間分布。在呼倫貝爾沙地,半固定沙地一般位于沙丘的坡頂或坡中,風(fēng)力較大,環(huán)境較干燥,而固定沙地位于沙丘的坡下,風(fēng)力小,環(huán)境相對不算太干燥。這可能是半固定沙地的種子比固定沙地的種子更易散布的原因之一。
因此,沙地微生境的變化對米氏冰草種子雨散布的時期及波動規(guī)律無影響,而對其有性繁殖體的形態(tài)特征具有顯著影響。
[1] Harper J L.Population Biology of Plants[M].London: Academic Press,1977:124-201.
[2] 馬萬里,荊濤,Kujansuu J,等.長白山地區(qū)胡桃楸種群的種子雨和種子庫動態(tài)[J].北京林業(yè)大學(xué)學(xué)報,2001,23(3):70-72.
[3] 馬杰,李慶芬,孫儒泳.嚙齒動物和鳥類對東靈山地區(qū)遼東櫟種子丟失的影響[J].生態(tài)學(xué)雜志,2004,23(1):107-110.
[4] 路紀(jì)琪,李宏俊,張知彬.嚙齒動物分散貯食的影響因素[J].生態(tài)學(xué)雜志,2005,24:528-532.
[5] 肖治術(shù),張知彬,王玉山.以種子為繁殖體的植物更新模型研究[J].生態(tài)學(xué)雜志,2003,22(4):70-75.
[6] Connell J H.On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees[A].In:Boer P J,Gradwell G R.Dynamics of Populations[M].Wageningen:Centre for Agricultural Publishing and Documentation,1971:298-312.
[7] Rabinowicz D,Rapp J K.Seed rain in a north American tall grass prairie:The resemblance of their abundance and compositions to dispersing seeds[J].Oikos,1980,36:191-195.
[8] Schott G W,Hamburg S P.The seed rain and seed banks of an adjacent tallgrass prairie old fields[J].Canadian Journal of Botany,1997,75:1-7.
[9] Watkinson A T,Harper J L.The demography of a sand dune annual,Vulpiafasciculate.Ⅲ.The dispersal of seeds[J].Journal of Ecology,1978,66:483-498.
[10] Westelaken I L,Maun M N.Spatial pattern and seed dispersal ofLithospermumcarolinienseon Lake Huron sand dunes[J].Canadian Journal of Botany,1985,63:125-132.
[11] Owen N W,Kent M,Dale M P.Spatial and temporal variability in seed dynamics of machair sand dune communities,Outer Hebriges,Scottland[J].Journal of Biogeograph,2001,28:565-588.
[12] Clobert J,Danchin E,Dhondt A A.Dispersal[M].Oxford:Oxford University Press,2001:103-132.
[13] Loiselle B A,Ribbens E,Vargas-Ramírez O R.Spatial and temporal variation of seed rain in a tropical lowland wet forest[J].Biotropica,1996,28(1):82-95.
[14] 班勇.植物生活史對策的進化[J].生態(tài)學(xué)雜志,1995,14(3):33-39.
[15] Barnes E M.Seed predation,germination and seedling establishment ofAcaciaeriolobain northern Botswana[J].Journal Arid of Environments,2001,49:541-554.
[16] Chambers J C.A day in the life of a seed: Movements and fates of seeds and their implications for natural and managed systems[J].Annual Review of Ecology and Systematics,1994,25:263-293.
[17] 楊允菲,祝廷成.松嫩平原大針茅群落種子雨動態(tài)的研究[J].植物生態(tài)學(xué)與地植物學(xué)學(xué)報,1991,15:46-55.
[18] 鄒春靜,徐文鐸,劉廣田.沙地云杉種群種子雨的時空分布規(guī)律[J].生態(tài)學(xué)雜志,1998,17(3):16-19.
[19] 胡星明,蔡永立,李愷.浙江天童常綠闊葉林栲樹種子雨的時空格局[J].應(yīng)用生態(tài)學(xué)報,2005,16(5):815-819.
[20] 金曉明,艾琳,盧欣石,等.不同群落中米氏冰草和羊草的年齡結(jié)構(gòu)動態(tài)[J].生態(tài)學(xué)報,2011,31(18):5406-5413.
[21] Janzen D H.Herbivores and the number of tree species in tropical forests[J].American Naturalist,1970,104:501-528.
[22] Urbanska K W,Fattorini M.Seed rain in high altitude restoration plots in Switzerland[J].Restoration Ecology,2000,8:74-79.
[23] 鐘章成.植物種群的繁殖對策[J].生態(tài)學(xué)雜志,1995,14(1):37-42.
[24] 操國興,謝德體,鐘章成.植物種群的生殖分配[J].四川林業(yè)科技,2003,24(2):25-29.
[25] 金曉明.米氏冰草繁殖生態(tài)學(xué)特征及種群競爭機制[D].北京:北京林業(yè)大學(xué),2011:36-46.
[26] 黃世華,任媛媛,張世挺.根競爭對窄葉野豌豆生長的影響:公共的悲劇[J].草業(yè)科學(xué),2011,28(2):266-271.
[27] 武建雙,沈振西,張憲洲,等.藏北高原人工垂穗披堿草種群生物量分配對施氮處理的響應(yīng)[J].草業(yè)學(xué)報,2009,18(6):113-121.
[28] 徐慶,劉世榮,臧潤國.中國特有植物四合木種群的生殖生態(tài)特征—種群生殖值及生殖分配研究[J].林業(yè)科學(xué),2001,37(2):36-41.
[29] Gashwiller J S.Conifer seed survival in Western Oregon clearcuts[J].Ecology,1967,48(3):431-438.
[30] 沈澤昊,呂楠,趙俊.山地常綠落葉闊葉混交林種子雨的地形格局[J].生態(tài)學(xué)報,2004,24(9):1981-1987.
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
