波斯菊核型分析
2012-12-24 00:53:38龍雙紅
草業科學 2012年11期
張 凡,張 芹,龍雙紅
(1.河北農業大學園林與旅游學院,河北 保定071001;2.河北省塞罕壩機械林場總場,河北 圍場068466)
波斯菊(Cosmosbipinnatus)原產于墨西哥,為菊科秋英屬一年生草本花卉,其葉形雅致,花色豐富,花期長,抗逆性強,栽培管理容易,是優良的園林綠化材料,常用作地被植物,也可用于花境、花叢或建筑墻垣的基礎栽植。目前有關波斯菊的研究主要集中在播種、育苗、栽培、應用等方面[1-3],而對于染色體核型的研究還比較少。Thomas[4]曾報道墨西哥部分波斯菊的染色體數目,楊德奎和周俊英[5]報道過山東部分地區波斯菊的核型,但仍缺乏不同地區不同環境條件下波斯菊染色體及核型更廣泛的研究。本研究對生長在海拔較高的塞罕壩地區的波斯菊染色體觀察、計數及核型分析,并與前人的研究進行比較,旨在為波斯菊的種質鑒定、起源演化及品種改良補充更有力的證據。
1 材料與方法
1.1 材料 供試波斯菊種子于2010年9月采自河北省塞罕壩機械化林場,置于4℃冰箱,保存備用。
1.2 方法 取飽滿的種子,用蒸餾水浸泡12h后,在25℃恒溫箱中培養。當幼根長約1cm時,選取根毛粗壯、根尖白嫩的幼根用0.002mol·L-1的8-羥基喹啉在20℃恒溫箱中預處理2h,然后轉入卡諾固定液20℃固定24h。取出根尖,用蒸餾水沖洗3~5遍后,放入1mol·L-1的HCl在60℃水浴中解離6min。取解離好的幼根分生組織放在載玻片中央,加1滴鐵礬-蘇木精染色劑,蓋蓋玻片,在酒精燈上加熱數秒,再輕壓使材料盡量分散。然后在Olympus BH22光學顯微鏡下觀察,選擇染色清晰、染色體分散而不重疊的分裂中期細胞進行染色體數目統計并拍照[6]。
染色體計數和核型分析依據李懋學和陳瑞陽[7]的分析標準進行,統計分散較好的30個以上分裂中期細胞,以85%以上的細胞具有相同的染色體數作為該種的染色體數,并選取分散良好的5個中期分裂相進行分析測量,取平均值計算核型數據。
核型類型分析參照Stebbins[8]的分類標準,染色體類型分析按照Levan等[9]的分類系統。核型不對稱系數按Arano[10]的方法計算,比值越大越不對稱。
2 結果與分析
2.1 波斯菊的染色體數目 選擇分散良好的30個中期分裂相的細胞計數,其中28個細胞染色體數目為24條(圖1),占計數細胞的93.3%。根據染色體的長度、臂比、著絲粒的位置、隨體的有無和位置等進行同源染色體配對,每個細胞的24條染色體可配成12對,表明均為二倍體,染色體基數為n=12。
2.2 波斯菊染色體核型分析 預處理及固定條件不同會直接影響染色體的短縮程度,所以即使是同一植物,不同研究所得到的絕對長度數值也會有明顯差異,而相對長度則是一組可比性更高的、穩定的數值。因此,本研究通過精確測量5個分散良好的中期分裂相的細胞染色體的長臂、短臂的相對長度,并通過計算得到波斯菊染色體的核型參數。在波斯菊12對同源染色體中,第6、8和11號染色體的臂比在1.71~3.00,為近中著絲粒染色體(sm型),占染色體總數的25%;其余染色體的臂比在1.01~1.70,為中間著絲粒染色體(m型),占染色體總數的75%(表1)。本研究發現2對具有隨體的染色體,分別為6和12號,隨體在染色體短臂的近端部(圖2)。因此,可以確定波斯菊的染色體核型公式為2n=2x=24=18m(1sat)+6sm(1sat)。
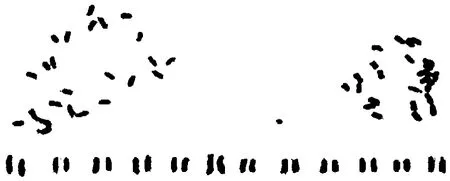
圖1 波斯菊的染色體及核型Fig.1 Chromosome and karyotype of Cosmos bipinnatus
波斯菊染色體由14條中長染色體(M2)和8條中短染色體(M1)及2條短染色體(S)組成,染色體相對長度平均值在6.20%~9.06%(表1)。根據Stebbins[8]的“不對稱核型分析”分類標準,波斯菊最長與最短的染色體長度比值為1.46,臂比大于2∶1的染色體為2對,占染色體總數的16.7%,應屬于“2A”型;核型不對稱系數為43.25%,對稱程度較高,為較對稱類型。
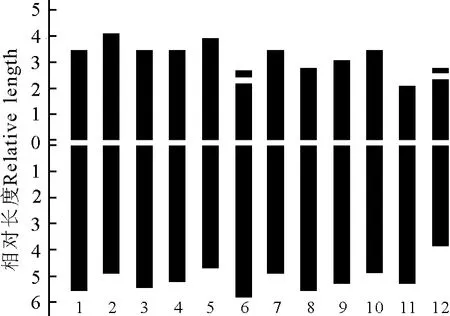
圖2 波斯菊的染色體核型模型圖Fig.2 Karyotype diagram of Cosmos bipinnatus
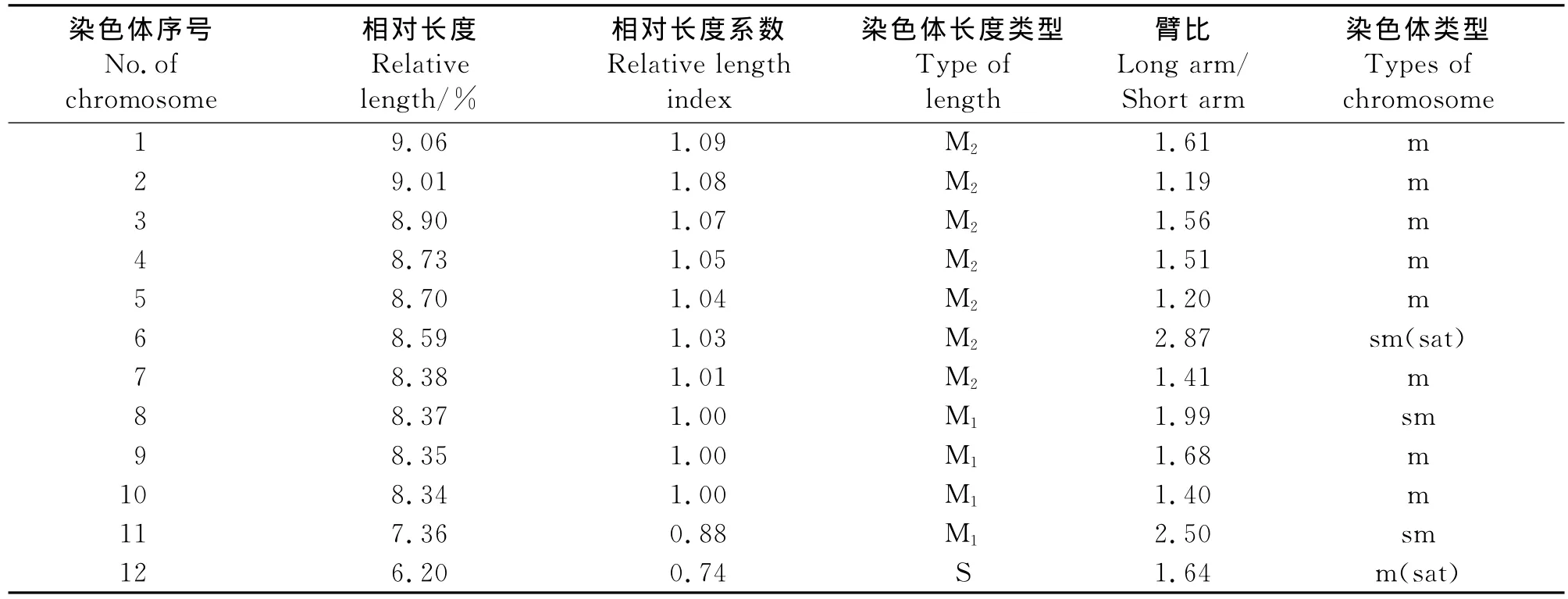
表1 波斯菊的核型參數Table 1 Karyotype index of Cosmos bipinnatus
3 討論
菊科秋英屬植物全世界約25種,我國常見栽培的有波斯菊和硫華菊(C.sulphureus)兩種[11]。國外有關該屬植物染色體數目報道的有10種左右,染色體基數分為24和12兩類[4]。本研究中波斯菊的染色體基數為12,與國外的報道一致,也與楊德奎和周俊英[5]研究報道的染色體數目相同,雖然以上研究取材的地點差異很大,但其染色體數目受環境影響較小,具有很高的穩定性和再現性,因此可以確定波斯菊為二倍體,有24條染色體。
本研究與楊德奎和周俊英[5]的報道一致,認為波斯菊的核型類型為“2A”型,屬于較對稱類型。而與其同屬的硫華菊的染色體核型公式為2n=24=2sm+22st,核型類型為“4B”型,是很不對稱的核型[12]。根據高等植物染色體的進化理論[8],可以確定波斯菊比同屬的硫華菊更原始。
山東地區的波斯菊的核型公式為2n=24=16m+2m(sat)+6sm,其染色體只有1對帶隨體[5],而本研究取材于塞罕壩的波斯菊的染色體中有2對帶隨體,分別是中部著絲粒染色體(m)和近中部著絲粒染色體(sm),并且染色體的長度與前人報道也有一定差異。這可能與生境不同有關,在其他種的染色體研究中也有不同居群核型差異的報道[13-15],要證實這一結論還需要擴大取樣地點進一步研究,甚至有必要結合分子生物學的手段進行深入研究。另外,染色體長度在預處理過程中受藥劑種類、濃度及處理時間的影響較大,試驗中所采取的處理方法、觀察手段的差別,也可能是導致試驗結果不同的原因之一,在今后的研究中應進一步優化方法,提高核型分析的精確度。
致謝:對河北農業大學園藝學院張成合老師在制片過程中給予的指導和幫助表示感謝!
[1]杜壽輝,徐濤,何素瑞,等.昆明地區波斯菊植物結實率及種子萌發特性研究[J].安徽農業科學,2010,38(28):15493-15494.
[2]潘永飛,戴忠良,潘葉飛,等.播種期對波斯菊結實和種子發芽率的影響[J].金陵科技學院學報,2009,25(2):64-67.
[3]張娟,楊玉想.大花波斯菊在北方地區的栽培與應用[J].現代園藝,2010(9):20.
[4]Thomas E M.Systematic studies in the Coreopsidinae:Cytotaxonomy of Mexican and Guatemalan Cosmos[J].American Journal of Botany,1968,55(3):345-353.
[5]楊德奎,周俊英.山東10種植物的核型分析[J].廣西植物,1999,19(4):349-354.
[6]李懋學,張贊平.作物染色體及其研究技術[M].北京:中國農業出版社,1996:5-17.
[7]李懋學,陳瑞陽.關于植物核型分析的標準化問題[J].武漢植物學研究,1985,3(4):297-302.
[8]Stebbins G L.Chromosomal Evolution in Higher Plants[M].London:Edward Arnold,1971:85-104.
[9]Levan A,Fredga K,Sandberg A A.Nomenclatrue for centromeric position of chromosomes[J].Hereditas,1964,52:201-220.
[10]Arano H.Cytological studies in subfamily carduoideae(Compositae)of Japan,IX[J].Botannical Magazine(Tokyo),1963,76:32-40.
[11]賀士元.河北植物志(第三卷)[M].石家莊:河北科學技術出版社,1991:75-76.
[12]汪小蘭,李懋學.10種菊科花卉的染色體觀察[J].武漢植物學研究,1987,5(2):111-117.
[13]時麗冉,高汝勇,李會芬,等.紫茉莉染色體數目及核型分析[J].草業科學,2010,27(1):53-54.
[14]張曉燕,毛培春,孟林,等.三份偃麥草種質的染色體核型分析[J].草業學報,2011,20(4):194-200.
[15]周自瑋,奎嘉祥,鐘聲,等.云南野生鴨茅的核型研究[J].草業科學,2000,17(6):48-51.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年18期)2018-11-14 01:48:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
