農桿菌介導的苜蓿次級體細胞胚的遺傳轉化
2012-02-09 09:34:06劉文婷段琦梅劉景玲孫延芳
生物工程學報 2012年2期
劉文婷,段琦梅,劉景玲,孫延芳
1 西北農林科技大學生命科學學院,陜西 楊凌 712100
2 遼寧工程技術大學理學院,遼寧 阜新 123000
苜蓿Medicago sativa L. 是世界上分布最廣泛的一種豆科多年生牧草,素有“牧草之王”的美譽,是目前國內外最重要的豆科牧草[1]。苜蓿在土壤保持和固氮方面也有重要的作用[2]。在過去幾年中,許多研究已經探討了使用農桿菌的苜蓿遺傳轉化[3-9]。但是,轉化方法仍有很大的局限性,比如轉化效率低、基因型依賴性、轉化程序復雜和轉化時間長、玻璃化植株比例高[3,5-7,9-11]。因此,優化轉化方法有助于苜蓿轉基因的廣泛應用。
先前的研究已經探索了不同類型外植體能提供苜蓿轉化優化的可能性,也提供了多基因轉化到苜蓿植株的可行性。在裸子植物和一些谷類作物中已經使用了體細胞胚的遺傳轉化方法[12-14],結果表明,使用體細胞胚的轉化方法具有很大的優勢,轉化效率高、減少逃逸和轉化程序快[13,15-16]。苜蓿在體細胞胚再生研究中是模式植物[17-19]。但是,很少有研究使用苜蓿體細胞胚作為外植體進行遺傳轉化。Ninkovic等[20]做了使用體細胞胚的苜蓿轉化研究,但是,在研究中沒有提供Southern雜交數據,不能從分子水平證實使用體細胞胚進行苜蓿轉化方法的可行性。
本研究中,我們使用農桿菌介導的次級體細胞胚的遺傳轉化方法,試驗證明此方法速度快、且簡單易行,為多基因在同一受體的共轉化實現苜蓿優良性狀改良提供理論依據。
1 材料與方法
1.1 材料
1.1.1 植物材料
按照Tian等[21]的方法,使用葉柄外植體誘導初級體細胞胚:選取生長60~90 d的野生型苜蓿無菌苗的葉柄作為外植體,用解剖刀切成8 mm長的小段,置于SH2K愈傷誘導培養基上,20 d繼代1次,形成的愈傷組織轉到胚形成培養基BOi2Y中21 d后,體細胞胚形成,選取子葉期的初級體細胞胚用作外植體來誘導次級體細胞胚。典型的初級體細胞胚如圖 1A所示。
1.1.2 培養基
SH2K培養基:用于誘導愈傷組織。主要成分見文獻[22-23]:1 mg/L 2,4-D、0.2 mg/L Kinetin生長調節劑,4.35 g/L硫酸鉀,288 mg/L脯氨酸,53 mg/L硫代脯氨酸,200 mg/L肌醇,30 g/L蔗糖,2.5 g/L植物膠 (pH 5.8)。
BOi2Y培養基:用于誘導體細胞胚形成。主要成分見文獻[24]:0.2%酵母提取物,100 mg/L肌醇,30 g/L蔗糖,2.5 g/L的植物膠(pH 5.9)。
MSO培養基:用于誘導體細胞胚的萌發。主要成分見文獻[25]:4.3 g/L MS鹽,1 mg/L氨基乙酸,40 g/L蔗糖,8 g/L的瓊脂 (pH 5.8)。

圖1 初級體細胞胚的誘導和次級體細胞胚的形成Fig. 1 Induction of primary somatic embryos (SE) and development of secondary SE. (A) Primary mid-cotyledonary stage SE used for explants. (B) Calli formed at the cut surface of the explants after 3 weeks of culture in SH2K medium. (C) Secondary SE formed after 4 weeks of culture in BOi2Y medium. (D) Globular embryos were observed in BOi2Y medium. (E) Heart embryos were observed in BOi2Y medium. (F) Torpedo embryos were observed in BOi2Ymedium. (G) Pre-cotyledon embryos were observed in MSO medium. (H) Embryos germination, root and leaf development.
1/2 MSO培養基:用于誘導植株的形成。主要成分見文獻[25]:2.65 g/L MS鹽,1 mg /L氨基乙酸,20 g/L蔗糖,8 g/L瓊脂 (pH 5.8)。
1.1.3 農桿菌菌株和質粒
試驗用農桿菌菌株為GV3101[26],表達載體為pCambia2301 (圖2),pCambia2301質粒由加拿大南方作物與食品保護中心田立寧實驗室提供。其包含β-葡萄糖苷酶報告基因 (gus) 和抗性篩選標記新霉素磷酸轉移酶基因 (nptⅡ),這 2個基因均受CaMV 35s啟動子調控,其中gus基因包含一個內含子,其可消除殘余菌液中的 gus基因表達行為。pCambia2301載體通過電轉化的方法導入到農桿菌 GV3101菌株[27]。該載體T-DNA區域的結構見圖2。
1.2 試驗方法
1.2.1 不同培養基對N4.4.2苜蓿次級體細胞胚形成的影響
SH2K[22-23]和 B5h[28]用于誘導來自初級體細胞胚的胚性愈傷。初級體細胞胚放在愈傷誘導培養基中,胚性愈傷組織形成,21 d后在新的誘導培養基中繼代,形成好的愈傷組織轉到胚形成培養基BOi2Y中,14 d后體細胞胚形成。
1.2.2 不同苜蓿基因型對次級體細胞胚的影響
N4.4.2、XAK-6A-10、F1-1和XAK-6A-7四個苜蓿基因型用于研究不同基因型對次級體細胞胚的影響。
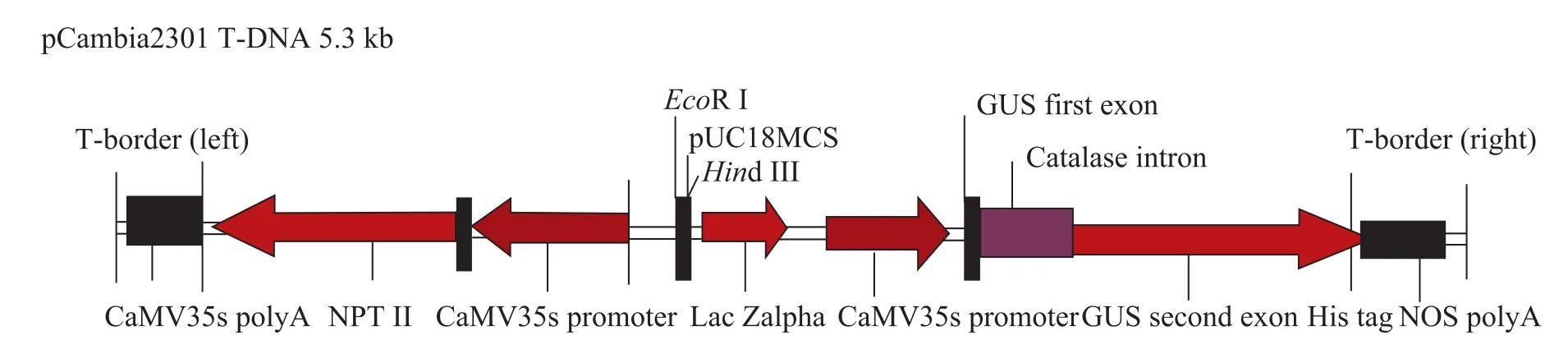
圖2 表達載體pCambia2301的T-DNA結構Fig. 2 Schematic representation of the T-DNA (5.3 kb) of pCambia2301.
1.2.3 體細胞胚的遺傳轉化和植株的恢復
選取子葉期的體細胞胚作為外植體,用解剖刀切開,置于SH2K培養基上預培養2 d。將?20 ℃下保存的農桿菌在LB培養基上活化培養1~2 d,達到對數生長期后,測定農桿菌菌液OD600為0.5~1.0時,用農桿菌侵染預培養的外植體4 min,用無菌濾紙吸去多余菌液,轉移外植體至含20 μmol/L乙酰丁香酮的SH2K培養基上,共培養2 d,預培養和乙酰丁香酮用于提高轉化效率[29]。隨后,在含75 mg/L卡那霉素的SH2K愈傷形成培養基中培養,篩選抗性愈傷。繼代培養2~3次后,將生長旺盛、外觀黃綠色、疏松的抗性愈傷組織轉移到含 75 mg/L卡那霉素的BOi2Y胚形成培養基上,14 d后,新鮮的胚狀愈傷組織上出現綠色芽點,30 d后體細胞胚形成。將再生的綠色體細胞胚轉移到含75 mg/L卡那霉素的MSO[25]胚萌發培養基上進行培養,28 d后體細胞胚生根萌芽,根和葉形成。將形成的植株再轉到含有75 mg/L卡那霉素的1/2 MSO植株形成培養基中培養,然后選擇根系生長健壯、株高約10 cm的植株移栽于溫室中或用于轉基因體細胞胚的誘導。所有的培養均在25 ℃、24 h光周期、3 000 lx光照條件下進行。每一步的轉化培養都是在75 mg/L的卡那霉素篩選壓下進行。
1.2.4 GUS組織化學定位和熒光檢測
轉化植株形成后,取苜蓿植株各組織,用于GUS組織定位分析和熒光定量檢測分析[30],通過組織化學分析確定 GUS表達的穩定性。將植物組織浸沒在含有5-溴-4-氯-3-吲哚β-D-葡萄糖苷 (X-Gluc) 溶液的 2 mL Eppendorf管中,X-Gluc 溶液中含:0.5 mmol/L鐵氰化鉀、0.5 mmol/L亞鐵氰化鉀、體積分數0.3% Triton X-100、1 g/L X-Gluc和50 mmol/L磷酸鹽緩沖液(pH 7.0),37 ℃保溫過夜。第2天將植物組織用不同梯度的酒精脫色,在顯微鏡下觀察組織染色情況。
1.2.5 轉化植株的PCR檢測
從轉化植株中,隨機選擇11個轉基因株系,按照Lodhi等[31]的方法,分別取幼嫩的植物葉片0.5 g提取基因組DNA。
采用 PCR 擴增檢測目的片段,以pCAMBIA2301載體上的gus和npⅡ基因序列為參考,設計二者的上、下游引物,其中gus正向引物為5¢-CGTCCTGTAGAAACCCCAAC-3¢,gus反向引物為5¢-ATTGACCCACATTTGCCGT-3¢,npⅡ正 向 引 物 為 5¢-GAGGCTATTCGGC TATGACTG-3¢,nptⅡ反向引物為 5¢-ATCGG GAGCGGCGATACCGTA-3¢。gus和nptⅡ基因的預期片段長度分別為300 bp和700 bp,引物由Invitrogen公司合成。PCR反應體系共50 μL,包括100 ng DNA,200 μmol/L dNTPs,1 μmol/L的上、下引物,1 U Taq DNA聚合酶,1.5 mmol/L MgCl2,5 μL 10×Taq DNA聚合酶緩沖液。反應程序為:95 ℃變性5 min;94 ℃變性30 s,58 ℃退火30 s,72 ℃延伸45 s,30個循環;72 ℃延伸5 min。取5 μL擴增產物于1%瓊脂糖凝膠中進行電泳檢測。
Southern blotting檢測中,提取轉基因苜蓿的基因組 DNA,采用紫外分光光度法結合瓊脂糖凝膠電泳法測定DNA純度和濃度。將10 μg DNA樣品以T-DNA中邊界具有單切點的限制性內切酶EcoRⅠ進行酶切過夜,在40 V電壓下,電泳8~10 h,凝膠經變性與中和后,轉膜和紫外交聯固定DNA之后與地高辛標記的nptⅡ探針雜交,雜交操作參照《分子克隆實驗指南》[32]并稍加改進。
1.2.6 數據的統計分析
試驗數據處理采用Excel 2003軟件,并使用SAS 8.0專業統計軟件進行最小顯著性檢驗(LSD,P≤0.05),數據用“平均值±平均標準誤差”表示,各組均數比較采用單因素方差分析 (One way Anova)。所有的試驗數據至少重復3次。
2 結果與分析
2.1 不同培養基對苜蓿次級體細胞胚的影響
圖1C是誘導的愈傷組織轉移到無生長調節劑的 BOi2Y培養基后,形成的次級體細胞胚。在愈傷誘導SH2K培養基中,次級體細胞胚的響應率是 89.29%,而在 B5h培養基的響應率是63.32% (表1)。數據統計分析表明,體細胞胚在不同培養基中存在顯著差異,次級體細胞胚的響應率在SH2K培養基中明顯高于在B5h培養基中。另外,對于響應的體細胞胚來說,在SH2K培養基中,每個初級體細胞胚誘導的次級體細胞胚數目7.00±0.94,而在B5h培養基中,每個初級體細胞胚誘導的次級體細胞胚數目是3.27±0.75,方差分析表明,在不同培養基中每個初級體細胞胚誘導的次級體細胞胚存在顯著差異 (P<0.05) (表1)。
2.2 不同苜蓿基因型對次級體細胞胚的影響
從2.1中得出SH2K培養基是苜蓿次級體細胞培養最優化的培養基,因此,將 4個苜蓿基因型N4.4.2、XAK-6A-10、F1-1和 XAK-6A-7的初級體細胞胚置于 SH2K培養基中誘導次級體細胞胚。100 mg的胚性愈傷組織產生的體細胞胚的數目,在 N4.4.2、XAK-6A-10、F1-1中沒有差異,而XAK-6A-7最低。4個基因型中,初級體細胞胚產生的次級體細胞胚存在明顯的差異,基因型N4.4.2初級體細胞胚產生次級體細胞胚的響應率為89.29%,而AK-6A-10、F1-1、XAK-6A-7三個基因型分別為54.55%、38.18%、38.10%,N4.4.2與其余3個基因型之間有明顯的差異。不同基因型次級體細胞胚轉化成植株的轉化效率也有顯著性差異 (P<0.05),N4.4.2的轉化效率為 (7.84±1.62) %,顯著高于其他3個基因型 (表2)。

表1 不同培養基對苜蓿基因型N4.4.2次級體細胞胚形成的影響Table 1 Effect of different media on the development of alfalfa N4.4.2 genetype secondary somatic embryos
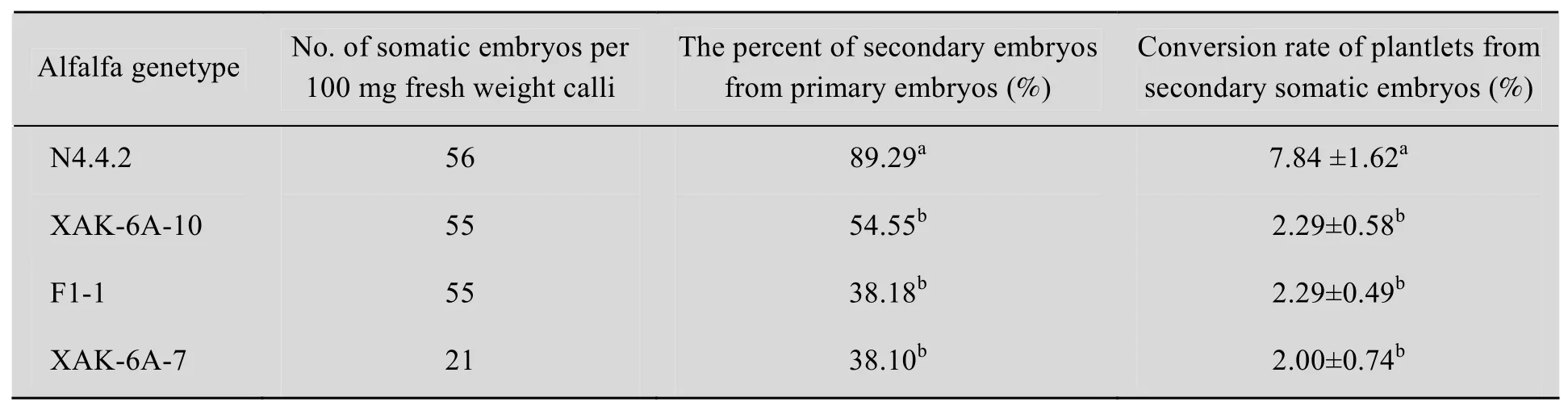
表2 不同苜蓿基因型對次級體細胞的的影響Table 2 Effect of different alfalfa genetype on the development of secondary somatic embryos
2.3 N4.4.2次級體細胞胚的遺傳轉化和轉化植株的組織化學定位分析
從2.1和2.2得出,SH2K培養基是次級體細胞胚優化培養基,N4.4.2苜蓿基因型是最佳基因型。因此,SH2K培養基和N4.4.2苜蓿的初級體細胞胚用于苜蓿的遺傳轉化。用農桿菌侵染N4.4.2苜蓿的初級體細胞胚,在含有卡那霉素篩選的SH2K培養基中連續繼代2~3次后,在外植體表面形成愈傷組織 (圖 1B)。隨后愈傷組織轉到胚形成培養基BOi2Y中,14 d后次級體細胞胚形成,不同時期的體細胞胚包括球形胚 (圖1D)、心形胚 (圖1E)、水雷胚 (圖1F)、子葉胚 (圖1G)。形成的體細胞胚轉到植株形成培養基MSO中,20 d后,次級體細胞胚萌發,胚軸、葉和根延伸生長,轉化苜蓿植株形成。隨后,轉化植株移栽至溫室 (圖 3C)。通過組織化學定位分析不同組織的GUS表達,愈傷組織 (圖3A)、體細胞胚 (圖3B)、葉 (圖3D)、根 (圖3E)、花 (圖3F)不同組織中均有GUS表達。在轉化試驗中,從供試的237個來自不同初級體細胞胚中,獲得了36個轉基因株系,共156個轉基因苜蓿植株。轉化效率是65.82% (表3)。
2.4 轉化植株的PCR分析和Southern雜交分析
用 PCR方法檢測轉化和未轉化苜蓿植株,以轉化植株和未轉化植株葉片的DNA為模板,使用gus和nptII基因的特異引物進行PCR擴增,
PCR產物經1%瓊脂糖凝膠電泳檢測可知,轉化苜蓿植株中分別可以檢測到300 bp和700 bp的 DNA擴增片段,而未轉化的野生型苜蓿植株中沒有DNA擴增片段 (圖4)。
同時,提取轉化植株葉片中的基因組DNA,用EcoR Ⅰ酶酶切基因組DNA。由于EcoR Ⅰ酶在T-DNA區有單一酶切位點,因此,用EcoR Ⅰ酶酶切、nptⅡ基因作為探針雜交,每一個雜交帶可以看作獨立整合位點[33],轉基因植株表現出不同整合情況,整合 nptⅡ基因的不同拷貝數 1~4 (圖5),Southern雜交證實了nptⅡ基因已經整合到植物基因組。

圖3 轉基因苜蓿的形成和GUS在不同轉基因植物中的組織化學定位分析Fig. 3 Development of transgenic alfalfa plants and GUS expression in different tissue by histochemical analysis. (A) GUS expression in calli from primary somatic embryos infected by Agrobacterium strain GV3101. (B) GUS expression in secondary somatic embryos. (C) Transgenic alfalfa plant in greenhouse. (D) GUS expression in leaf from recovered transgenic alfalfa plants. (E) GUS expression in root from recovered transgenic alfalfa plants. (F) GUS expression in inflorescence from recovered transgenic alfalfa plants.

表3 N4.4.2基因型苜蓿次級體細胞胚的遺傳轉化Table 3 Genetic transformation of alfalfa N4.4.2 genetype secondary somatic embryos
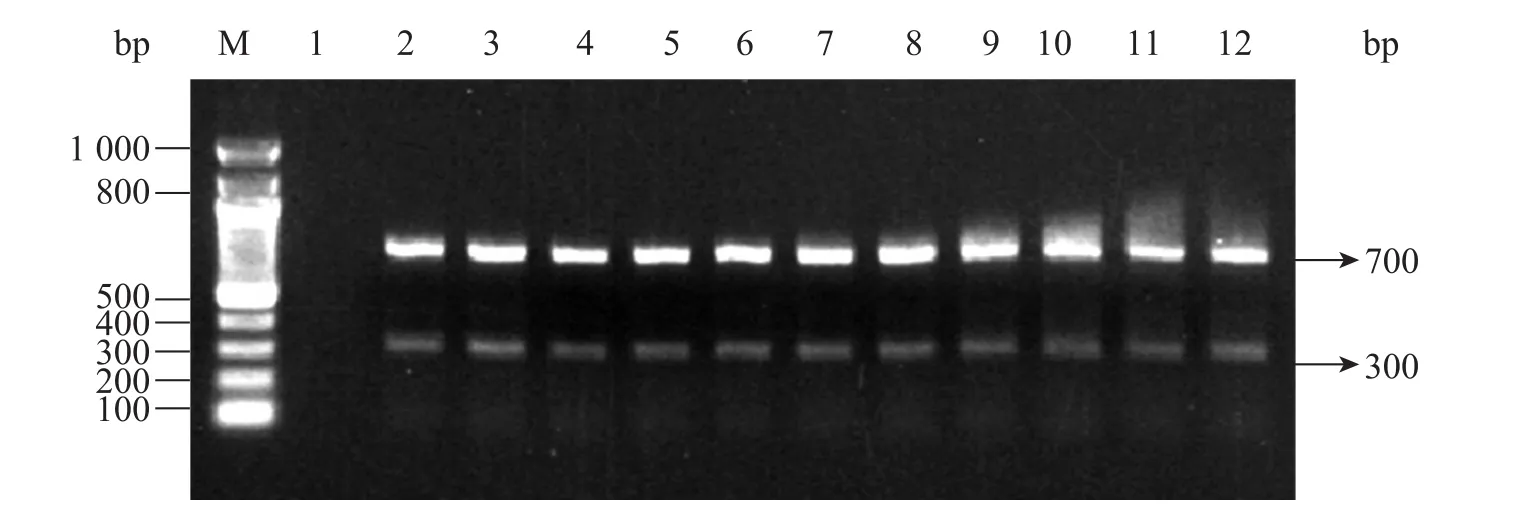
圖4 轉化和未轉化的苜蓿植株中GUS和nptII基因的PCR檢測Fig. 4 PCR analysis of GUS and nptII gene in transformed and non-transformed alfalfa. M: Marker DNA; 1: non-transformed control plant; 2?12: transformed plants.
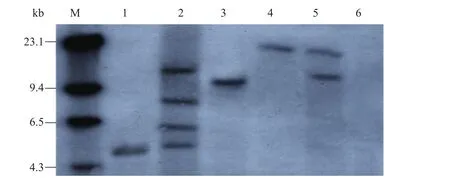
圖5 Southern雜交分析轉基因植株nptⅡ基因的拷貝數Fig. 5 Southern blotting analysis of transgenic plants to detect the copy number of npt Ⅱgene. M: molecular weight marker; 1–5: transgenic plants; 6: non-transgenic control plant.
3 討論
優良品種選育是苜蓿優質飼草高產的基礎,隨著基因工程技術的發展,轉基因技術為苜蓿優良品種選育開辟了一條新的途徑。苜蓿的遺傳轉化再生體系又是其轉基因的重要前提與基礎,它的再生體系主要是通過體細胞胚的發生途徑來實現。該途徑有很多優點,諸如體細胞胚具有在實驗室條件下容易獲得,并且轉化率較高;容易得到大量的轉化植株,便于后代的篩選。
本研究參照 Tian等[21]的方法,選擇不同苜蓿基因型和培養基,利用苜蓿初級體細胞胚為外植體進行農桿菌介導的遺傳轉化,建立并優化了農桿菌介導的紫花苜蓿遺傳轉化系統。研究表明,從胚性愈傷組織到體細胞胚的形成通常需要不同的培養基[34-35]。在本試驗中,首先在含有生長調節劑的SH2K和B5h培養基中誘導胚性愈傷組織,然后,轉移愈傷組織到無生長調節劑的胚形成培養基 BOi2Y中,誘導形成體細胞胚。在這2種培養基中,我們觀察到來自初級體細胞胚形成的愈傷組織是非常快速的,大量的愈傷組織的形成僅需要7 d時間。而來自葉柄、莖、葉外植體和別的類型的外植體愈傷組織的形成需要20 d[21]。同時,在這2種培養基中,初級體細胞胚在SH2K培養基中愈傷組織響應率是89.29%,誘導的體細胞胚的數目是 (7.00±0.94) 個,明顯優于B5h培養基。也就是說,單個初級體細胞胚通過SH2K培養基誘導次級體細胞胚數目達到了7個,在B5h培養基中誘導的次級體細胞胚的數目只達到了3個。而且,在實驗中還發現,來自畸形的初級體細胞胚誘導形成的次級體細胞胚是正常的。這說明初級體細胞胚作為外植體在SH2K培養基中大大提高了次級體細胞胚的誘導頻率。
植物的基因型也是影響苜蓿遺傳轉化成功與否的重要因素。在本試驗中,4個苜蓿基因型N4.4.2、XAK-6A-10、F1-1和XAK-6A-7置于優化的SH2K培養基中誘導次級體細胞胚,結果表明,N4.4.2與其他3個苜蓿基因型誘導的次級體細胞胚的響應率和次級體細胞胚轉化成植株的轉化率存在顯著差異,N4.4.2是4個基因型中最優化的基因型。因此,苜蓿農桿菌介導的遺傳轉化應注意選擇合適的基因型。這與先前的研究是一致的[3,6,9-10]。
N4.4.2基因型和SH2K培養基是本研究的優化基因型和培養基,用于農桿菌介導的遺傳轉化。組織化學定位、PCR和Southern雜交檢測表明,gus基因已經整合入苜蓿基因組中并得到正確的轉錄。從供試的237不同初級體細胞胚中,共獲得36個轉基因株系,156個轉基因植株。轉化效率是 65.82%。次級體細胞胚的遺傳轉化方法提高了苜蓿遺傳轉化效率。
目前,大多數轉化技術只是將單個目的基因的轉化用于植物的改良。但是,在植物體內,代謝途徑或數量性狀的遺傳修飾常常需要多個相關基因協同表達,因此,將相關多基因轉化到同一植株中,就顯得很有必要。本研究首次報道了苜蓿次級體細胞胚的遺傳轉化。在今后的試驗中,我們試圖轉入抗旱基因獲得轉基因苜蓿植株,隨后誘導轉基因苜蓿次級體細胞胚,將獲得的抗旱轉基因苜蓿植株次級體細胞胚作為外植體,轉入抗寒基因,最終實現多基因的共轉化苜蓿植株,為苜蓿多基因共轉化開辟途徑。
總之,在不同苜蓿品種中,初級體細胞胚能有效誘導次級體細胞胚,并能通過次級體細胞胚的遺傳介導再生為轉基因苜蓿植株。比較苜蓿植株其他的器官做外植體的常規轉化方法,這種新的轉化體系能縮短培養周期、提高誘導率。而且,提供了一個快速導入多基因到苜蓿植株中的方法。為苜蓿品質改進、分子生物學和基因組學的研究提供理論依據,這種方法也適用于別的轉化方法效率低的一些苜蓿品種,拓寬了苜蓿遺傳轉化方法。
REFERENCES
[1] Huang SX, LV DY, Shao JH, et al. Regeneration of transgenic plants through protoplasm of alfalfa (Medicago sativa L). Chin Sci Bull, 1991, 36(17): 1345?1347.黃紹興, 呂德揚, 邵嘉紅, 等. 紫花苜蓿原生質體轉基因植株再生. 科學通報, 1991, 36(17): 1345?1347.
[2] Huang WH, Liu ZX. Introduction of the Distribution and Development of Alfalfa. Beijing: Agricultural Press, 1995: 2?7.黃文惠, 劉自學. 概論苜蓿的分布和發展. 北京:中國農業出版社, 1995: 2?7.
[3] Desgagnés R, Laberge S, Allard G, et al. Genetic transformation of commercial breeding lines of alfalfa (Medicago sativa). Plant Cell Tissue Organ Cult, 1995, 42(2): 129?140.
[4] Austin S, Bingham ET, Mathews DE, et al. Production and field performance of transgenic alfalfa (Medicago sativa L.) expressing alpha-amylase and manganese-dependent lignin peroxidase. Euphytica, 1995, 85(1/3): 381?393.
[5] Samac DA. Strain specificity in transformation of alfalfa by Agrobacterium tumefaciens. Plant Cell Tissue Organ Cult, 1995, 43(3): 271?277.
[6] Xie XX, Lu Y, Liang J, et al. Establishment of an efficient Agrobacterium-mediated transformation system for genetic improvement of Alfalfa (Medicago sativa L.). J Agric Sci Technol, 2010, 12(1): 128?134.謝鑫星, 路揚, 梁晶, 等. 高效農桿菌介導的紫花苜蓿遺傳轉化體系的建立. 中國農業科技導報, 2010, 12(1): 128?134.
[7] Ziauddin A, Lee RWH, Lo R, et al. Transformation of alfalfa with a bacterial fusion gene, Mannheimia haemolytica A1 leukotoxin50-gfp: response with Agrobacterium tumefaciens strains LBA4404 and C58. Plant Cell Tiss Organ Cult, 2004, 79(3): 271?278.
[8] Montague A, Ziauddin A, Lee R, et al. High-efficiency phosphinothricin-based selection for alfalfa transformation. Plant Cell Tiss Organ Cult, 2007, 91(1): 29?36.
[9] De Marchis F, Bellucci M, Arcioni S. Handbook of New Technologies for Genetic Improvement of Legumes. Whitney: CRC Press, 2008: 29?43.
[10] Zare N, Valizadeh M, Tohidfar M, et al. Selection of regenerative genotypes from Iranian alfalfa cultivars. Int J Food Agri Environ, 2009, 7: 567?572.
[11] Zhang H, Huang QM, Su J. Development of alfalfa (Medicago sativa L.) regeneration system and Agrobacterium-mediated genetic transformation. Agri Sci China, 2010, 9(2): 170?178.
[12] Levée V, Garin E, Klimaszewska K, et al. Stable genetic transformation of white pine (Pinus strobus L.) after cocultivation of embryogenic tissues with Agrobacterium tumefaciens. Mol Breed, 1999, 5(5): 429?440.
[13] Leelavathi S, Sunnichan VG, Kumria R, et al. A simple and rapid Agrobacterium-mediated transformation protocol for cotton (Gossypium hirsutum L.): embryogenic calli as a source to generate large numbers of transgenic plants. Plant Cell Rep, 2004, 22(7): 465?470.
[14] Taniguchi T, Ohmiya Y, Kurita M, et al. Regeneration of transgenic Cryptomeria japonica D. Don after Agrobacterium tumefaciens-mediated transformation of embryogenic tissue. Plant Cell Rep, 2008, 27(9): 1461?1466.
[15] Fiore MC, Trabace T, Sunseri F. High frequency of plant regeneration in sunflower from cotyledons via somatic embryogenesis. Plant Cell Rep, 1996, 16(5): 295?298.
[16] Sun JY, Li WM, Zhang HS, et al. Somatic embryogenesis and plant regeneration in glandless upland cotton (Gossypium hirsutum L.). Front Agric China, 2009, 3(3): 279?283.
[17] Saunders JW, Bingham ET. Production of alfalfa plants from callus tissue. Crop Sci, 1972, 12(6): 804?808.
[18] McKersie BD, Brown DCW. Biotechnology and the Improvement of Forage Legumes. Wallingford: CABI, 1997: 111?143.
[19] McKersie BD, Senaratna T, Bowley SR, et al. Application of artificial seed technology in the production of hybrid alfalfa (Medicago sativa L.). In Vitro Cell Dev Biol, 1989, 25(12): 1183?1188.
[20] Ninkovi? S, Milju?-Duki? J, Vinterhalter B, et al. Improved transformation of alfalfa somatic embryos using a superbinary vector. Acta Biol Cracoviensia Ser Bot, 2004, 46: 139?143.
[21] Tian LN, Brown DCW, Watson E. Continuous long-term somatic embryogenesis in alfalfa. In Vitro Cell Dev Biol, 2002, 38(3): 279?284.
[22] Schenk RV, Hildebrandt AC. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can J Bot, 1972, 50(1): 199?204.
[23] Shetty K, McKersie BD. Proline, thioproline and potassium mediated stimulation of somatic embryogenesis in alfalfa (Medicago sativa L.). Plant Sci, 1993, 88(2): 185?193.
[24] Blaydes DF. Interaction of kinetin and various inhibitors in the growth of soybean tissue. Physio Planta, 1966, 19(3): 748?753.
[25] Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physio Planta, 1962, 15(3): 473?497.
[26] Koncz C, Schell J. The promoter of TL-DNA gene 5 controls the tissue-specific expression of chimaeric genes carried by a novel type of Agrobacterium binary vector. Mol Gen Genet, 1986, 204(3): 383?396.
[27] Hood EE, Gelvin SB, Melchers LS, et al. New Agrobacterium helper plasmids for gene transfer to plants. Transgenic Res, 1993, 2(4): 208?218.
[28] Gamborg OL, Miller RA, Ojima K. Nutrient requirements of suspension cultures of soybean root cells. Exp Cell Res, 1968, 50(1): 151?158.
[29] Lopez SJ, Kumar RR, Pius RK, et al. Agrobacterium tumefaciens-mediated genetic transformation in tea (Camellia sinensis [L.] O. Kuntze). Plant Mol Biol Rep, 2004, 22(2): 201?202.
[30] Jefferson RA. Assaying chimeric genes in plants: the gus gene fusion system. Plant Mol Bio Rep, 1987, 5(4): 387?405.
[31] Lodhi MA, Ye GN, Weeden NF, et al. A simple and efficient method for DNA extraction from grapevine cultivars and Vitis species. Plant Mol Biol Rep, 1994, 12(1): 6?13.
[32] Sambrook J, Russell DW. Molecular Cloning: A Laboratory Manual. 3rd ed. Beijing: Science Press, 2002: 156?163.
[33] Trine JM, Kamfjord D, H?kelien AM, et al. The frequency of silencing in Arabidopsis thaliana varies highly between progeny of siblings and can be influenced by environmental factors. Transgenic Res, 2001, 10(1): 53?67.
[34] Duval Y, Engelmann F, Durand GT. Somatic Embryogenesis and Synthetic Seed I: Biotechnology in Agriculture and Forestry. Berlin: Springer Verlag, 1995: 335?352.
[35] Akhtar N, Kumari N, Pandey S, et al. Somatic Embryogenesis in Tropical Fruit Trees. Dordrecht: Kluwer, 2000: 93?144.
