非O157產志賀毒素大腸桿菌分離株的多位點序列分型研究*
2012-01-24 02:13:14白向寧趙愛蘭夏勝利熊衍文徐建國
中國人獸共患病學報 2012年6期
關鍵詞:檢測
白向寧,趙愛蘭,夏勝利,熊衍文,徐建國
產志賀毒素大腸桿菌(Shiga toxin-producing Escherichia coli,STEC)是一類重要的食源性病原菌,可引起水樣腹瀉、出血性腸炎、溶血性尿毒綜合征(Hemolytic-uremic syndrome,HUS)等病死率高的疾病[1]。自1982年美國首次報道由O157∶H7引起的暴發以來,世界各國相繼報道了不同血清型的STEC散發感染和暴發[2-5],其中以 O157∶H7血清型為主。目前發現有超過200種非O157血清型的STEC能引起人類疾病[6]。在對全球35個國家的腸道感染病例監測中發現,2002-2005年非O157STEC感染率上升了60.5%,而 O157 STEC只有13%的上升[7]。特別是2011年5月,德國出現一起由O104∶H4感染引起的暴發疫情,共造成4 075人發病,其中50人死亡(WHO,2011年7月22日數據),提示未來非O157STEC的流行或暴發可能會成為公共衛生領域的一個重要問題。
早在1998年世界衛生組織就已提出將非O157 STEC的快速檢測作為公共健康的首要問題之一(WHO,1998)。美國自2001年起開展了對非O157STEC感染的主動監測,非O157STEC感染的報告病例逐年上升[4]。歐盟于1998年通過了人感染STEC病例報告的相關法案。2005-2009年歐盟國家每年報告的STEC感染病例數在3 000例左右,其中非O157感染病例近二分之一[8]。我國1999年蘇皖出現O157∶H7感染大暴發,為此2000年衛生部發布了《全國腸出血性大腸桿菌O157∶H7感染性腹瀉監測方案(試行)》,加強了我國重點地區的O157∶H7的監測工作,但目前我國尚未系統開展非O157STEC感染調查和病原菌檢測工作。本研究通過對我國分離的部分非O157STEC菌株進行血清型鑒定、主要毒力基因檢測及分子分型分析,旨在初步了解我國這類菌株的分子流行病學特征和遺傳背景。
1 材料與方法
1.1 材料
1.1.1 菌株來源 28株非O157STEC菌株2002年4-8月分離自河南某波爾山羊場羊糞,1株2009年分離自黑龍江某農村肉牛糞。菌株經中國疾病預防控制中心傳染病預防控制所新病原室復核和O157及H7血清型鑒定排除后-80℃保存。
1.1.2 主要試劑與儀器 大腸桿菌O及H抗原全套血清購自丹麥血清學研究所(Statens Serum Institut,Denmark),致病性大腸桿菌O抗原診斷血清購自天津生物芯片技術有限責任公司;La Taq DNA聚合酶、10×PCR Buffer、dNTPs及DL2000 DNA Marker購自大連TaKaRa公司;細菌基因組提取試劑盒(Bacteria Gene DNA kit)購自康為世紀公司。PCR儀(SensoQuest Labcycler)購自德國Senso公司;凝膠成像系統(Gel Doc2000)購自美國Bio-Rad公司;引物由生工生物工程(上海)有限公司合成;PCR產物純化及測序由北京天一輝遠生物科技有限公司完成。
1.2 方法
1.2.1 血清型鑒定 O血清型鑒定使用丹麥血清研究所和天津生物芯片有限公司生產血清套裝,鑒定步驟按照說明書進行。H血清型的分型首先采用fliC基因全長引物(見表1),PCR擴增后測序,根據序列比對結果判定型別,最后以H抗原診斷血清復核。
1.2.2 DNA模板的制備 細菌基因組提取按試劑盒說明書進行,制備好的模板置-20℃凍存備用。
1.2.3 毒力基因檢測 對29株菌的3個毒力基因stx1、stx2、eaeA進行PCR鑒定。PCR引物序列及產物大小見表1,PCR擴增條件為:預變性95℃15 min,變性94℃30s,退火57℃90s,延伸72℃90s循環30次,最后72℃ 延伸10min[9]。
1.2.4 管家基因擴增 根據E.coli MLST數據庫(http://mlst.ucc.ie/mlst/dbs/Ecoli)提供的大腸桿菌MLST分型方案,選擇7個管家基因adk、fumC、gyrB、icd、mdh、purA、recA,PCR 引 物 序列、退火溫度及產物大小見表1。PCR反應采用50 μL體系,包括10μmol/L的上下游引物各2μL,2 mmol/L dNTPs 2.5μL,含 MgCl2的 10×PCR Buffer 5.0μL,5U/μL的La Taq DNA聚合酶0.5 μL,加雙蒸水至49μL,模板DNA 1.0μL。PCR反應條件為:預變性95℃4min;變性94℃30s,退火45s,延伸72℃1min,循環30次;72℃ 延伸7 min。每次擴增均以無DNA模板的體系作為空白對照。取PCR產物3μL以1%瓊脂糖凝膠電泳進行檢測。PCR產物經純化后,用相應測序引物(表1)進行雙向測序。
1.2.5 序列分析 用SeqMan II(DNAStar)軟件并結合菌株等位基因的正反向序列圖譜對等位基因序列進行拼接和校正。將校正后的序列在E.coli MLST數據庫中進行比對,確定菌株的等位基因型和序列類型。用 MEGA 4.0、eBURST V3軟件分析不同ST型克隆群及菌株之間的親緣關系。
2 結 果
2.1 血清型鑒定結果 29株非O157STEC菌株的血清型鑒定結果見表2,主要血清型為O103∶H8(8/25)和 O21∶H25(6/29),屬于 O91∶H14、O12∶H21、O45∶H2、O15∶H21血清型的分別有2株菌,有5株菌分別屬于5種不同的血清型,另有2株菌的O血清型未能確定。
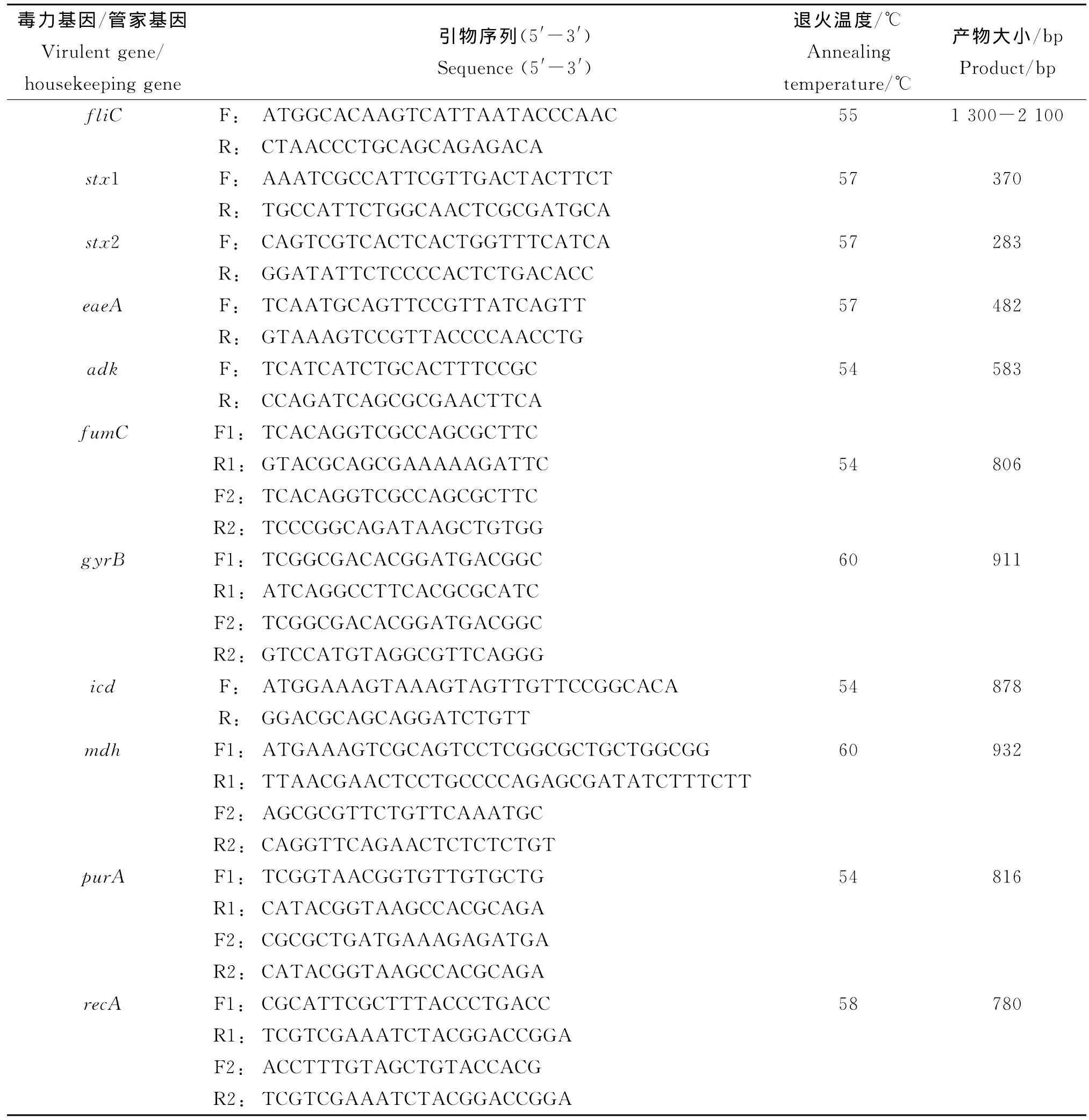
表1 fliC,毒力基因和7個管家基因的PCR引物Tab.1 PCR primers of fliC,virulent genes and seven housekeeping genes
2.2 毒力基因檢測結果 29株非O157STEC菌株的eaeA基因均為陰性,僅stx1基因陽性的有19株菌(65.52%),僅stx2基因陽性的有4株菌(13.79%),stx1、stx2 基 因 同 時 陽 性 的 有 6 株 菌(20.69%),菌株的主要毒力基因攜帶情況見表2。
2.3 MLST分型結果 應用7個管家基因的MLST分型方法,29株非O157STEC可以分成13個ST型,結果見表2。本研究發現2個新的等位基因型fumC376、recA214和3個新的序列類型ST2460、ST2467、ST2468。不同ST型別包含的菌株數有差異,其中ST155包含10株菌(34.48%),為本次研究的優勢型別,其次為ST906(17.24%)、ST446(10.34%)、ST33(6.90%),其余9個序列型均只包含1株菌。
2.4 遺傳進化分析 利用MEGA 4.0軟件從堿基組成的角度對13個ST型別及國外STEC流行的優勢 血 清 型 (O157、O26、O103、O111、O145 及O104)菌株的6個ST型別之間做了系統發生分析,將7個管家基因的序列按順序拼接后采用Neighbor-joining法繪制進化樹(圖1),Bootstrap值的計算分析根據1 000次隨機抽樣進行。結果顯示,我國非O157STEC菌株除一些ST型別相對特異外,存在與國際上優勢血清型親緣關系較近的ST型菌株。
通過eBURST V3對MLST數據庫中所有STEC相關的ST型進行聚類分析,按至少有4個以上等位基因相似的參數原則,將不同的ST歸為特定的ST序列群,ST型中單位點差異(SLV)和雙位點差異(DLVs)最多的ST型可作為ST序列群的核心序列 型 (group founder ST)[10]。 結 果 顯 示 (圖2),ST2460與ST155聚為1個ST序列群,其他11個ST型別散在分布。本研究中的3個型別ST25、ST33、ST446與國外其他ST型聚成3個ST序列群,且這3個ST型均為對應序列群的核心序列型。
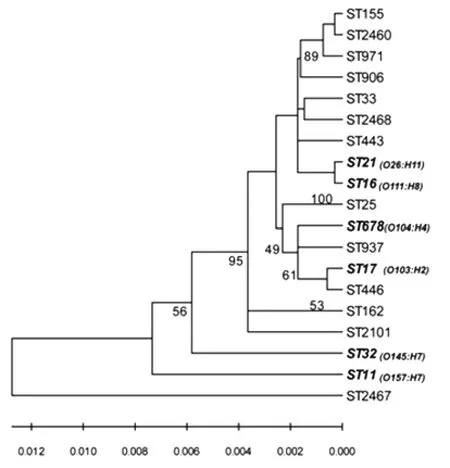
圖1 非O157產志賀毒素大腸桿菌不同ST型別的遺傳進化分析注:斜體加粗字體表示國際上優勢血清型STEC的ST型Fig.1 Phylogenetic tree analysis of different STs from 29 non-O157STEC isolatesNote:STs of dominant serotypes of STEC are indicated in bold-face italic type.
3 討 論
目前國際上報道的非O157STEC菌株的主要血清型 為 O26,O111,O103,O145,O91,O113,O128,O118等[2]。本研究對我國29株非O157STEC的血清分型結果發現以O103,O21為主,同時檢測出 O91,O113,O118,O12,O15等血清型的菌株,由此可見,我國部分STEC菌株在血清型分布上與國際流行株既存在相似性,也存在一些差異。這一方面可能與我們分析的菌株數量有關,另一方面可能是我們國家菌株本身的特點。我國衛生部《感染性腹瀉診斷標準》(ws271-2007)中,對腸出血性大腸桿菌的主要血清型除O157∶H7外,只列舉了O26和O111。根據本研究結果,提示在今后的STEC監測和檢測工作中,我們也應重視其他血清型的菌株。
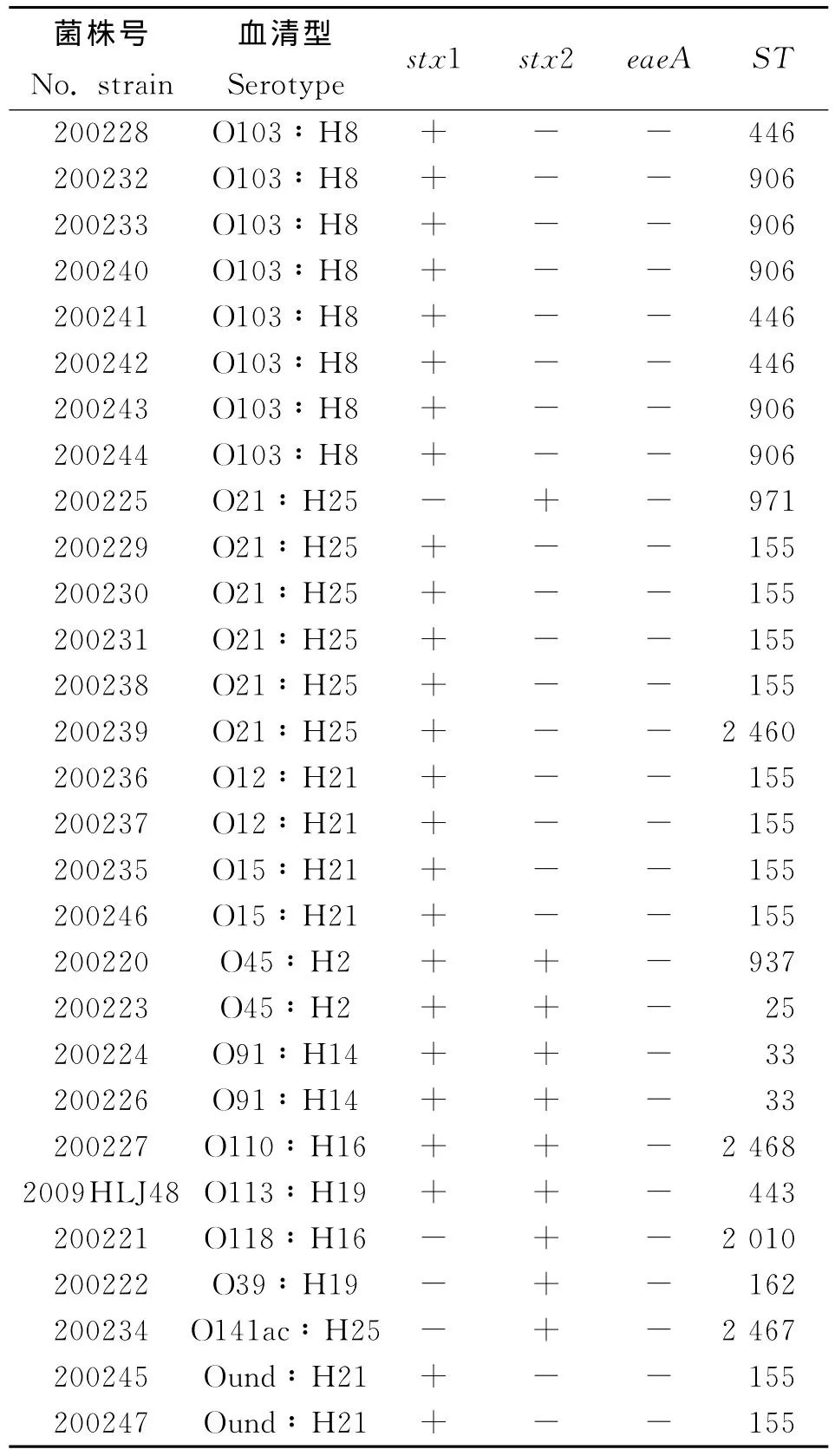
表2 29株非O157STEC的血清型鑒定、毒力因子檢測及MLST分型結果Tab.2 Serotypes,virulent gene profiles and sequence types of 29non-O157STEC isolates
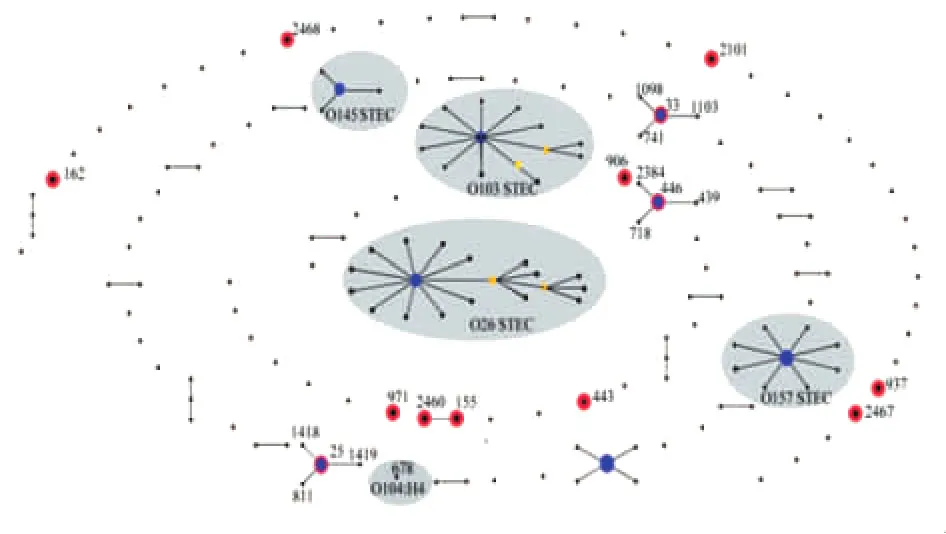
圖2 產志賀毒素大腸桿菌相關ST型別的eBURST聚類分析注:每個黑點代表1個序列型別,藍色節點表示group founder ST,黃色節點為subgroup founder ST。紅色圓圈內為本研究菌株的13個ST型別,灰色區域代表國際上幾個優勢血清型STEC的序列群。Fig.2 Population snapshots of STEC clusters for related STs in the E.coli MLST database by eBURST analysisNote:Black dots represent different STs;blue nodes represent predicted founder STs;yellow nodes represent sub-founders.Thirteen STs identified in this study are circled in red;grey area point to the ST complexes of common serotypes of STEC worldwide.
毒力基因檢測結果顯示,29株菌的eaeA基因均呈陰性,stx1基因陽性的菌株占了大多數(86.20%),僅stx2基因陽性菌株只有13.79%,這與王濤等[11]報道的我國O157STEC分離株的毒力基因特點明顯不同。研究推測,相對于O157∶H7,非O157STEC可能更多地攜帶stx1基因。Hedican等[12]對O157STEC和非O157STEC的毒力基因特點和致病性做了對比分析,與本研究結果相似。國外相關研究表明[13],eaeA陰性的非O157STEC及僅攜帶stx1毒力基因的非O157STEC同樣會導致嚴重的疾病,如出血性腹瀉、溶血性尿毒綜合癥等。我國非O157STEC菌株eaeA基因依賴以外的粘附機制,尚有待進一步研究。
對非O157STEC進行分子分型,對了解菌株的遺傳進化特征,暴發溯源等具有重要意義。目前用于病原微生物分子分型主要有基于表型和基因型兩大類。基于基因型的MLST作為一種準確的分子分型方法,自1998年Maiden[14]等首次運用于腦膜炎奈瑟菌的分型以來,以其分辨率高、數據可靠、重復性好、不同實驗數據便于比較、有利于全球范圍的分子流行病學數據標準化等特點而廣泛應用于其他病原菌的流行病學、遺傳進化、致病性等方面的研究。
本研究利用MLST方法對29株非O157 STEC進行分子分型研究,發現了2個新的等位基因型和3個新的序列型。大多數型別(12/13)散在分布,推測這些型別之間具有較遠的遺傳關系,具有明顯的分子多態性。Mellmann等[15]通過采用MLST方法,對從德國HUS患者分離的非O157 STEC進行分型分析,發現 ST16、ST17、ST21、ST25型別的菌株,O抗原檢測結果分別為O111、O103、O26、O128,推測這些ST型別及血清型的菌株可能與HUS密切相關。本次研究中發現有ST25型別的菌株,雖然我們分析的菌株O血清型和ST型別與他們的結果存在一定差異,但經系統發生分析發現,我們的菌株與國際流行株具有一定的親緣關系,提示我國應同時加強對這類STEC菌株的檢測和監測,掌握菌株的分布和變異規律,及時發現和甄別暴發,進而對于有效預防和控制這類疾病在我國的暴發和流行將具有重要的意義。
[1]Paton JC,Paton AW.Pathogenesis and diagnosis of Shiga toxinproducing Escherichia coli infections[J].Clin Microbiol Rev,1998,11(3):450-479.
[2]Johnson KE,Thorpe CM,Sears CL.The emerging clinical importance of non-O157Shiga toxin-producing Escherichia coli[J].Clin Infect Dis,2006,43(12):1587-1595.DOI:10.1086/509573
[3]Ethelberg S,Smith B,Torpdahl M,et al.Outbreak of non-O157Shiga toxin-producing Escherichia coli infection from consumption of beef sausage[J].Clin Infect Dis,2009,48(8):e78-81.DOI:10.1086/597502
[4]Mathusa EC,Chen Y,Enache E,et al.Non-O157Shiga toxinproducing Escherichia coli in foods[J].J Food Prot,2010,73(9):1721-1736.
[5]K?ppeli U,H?chler H,Giezendanner N,et al.Human infections with non-O157Shiga toxin-producing Escherichia coli,Switzerland,2000-2009[J].Emerg Infect Dis,2011,17(2):180-185.DOI:10.3201/eid1702.100909
[6]Coombes BK,Wickham ME,Mascarenhas M,et al.Molecular analysis as an aid to assess the public health risk of non-O157 Shiga toxin-producing Escherichia coli strains[J].Appl Environ Microbiol,2008,74(7):2153-2160.DOI:10.1128/AEM.02566-07
[7]Fisher IS,Meakins S,Enter-net participants.Surveillance of enteric pathogens in Europe and beyond:Enter-net annual report for 2004[J].Euro Surveill,2006,11(8):E060824.3.
[8]European Centre for Disease Prevention and Control and European Food Safety Authority.Shiga toxin/verotoxin-producing Escherichia coli in humans,food and animals in the EU/EEA,with special reference to the German outbreak strain STEC O104[R].Stockholm:ECDC;2011.
[9]Brandal LT,Lindstedt BA,Aas L,et al.Octaplex PCR and fluorescence-based capillary electrophoresis for identification of human diarrheagenic Escherichia coli and Shigella spp[J].J Microbiol Methods,2007,68(2):331-341.DOI:10.1016/j.mimet.2006.09.013
[10]Feil EJ,Li BC,Aanensen DM,et al.eBURST:inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data[J].J Bacteriol,2004,186(5):1518-1530.DOI:10.1128/JB.186.5.1518-1530.2004
[11]Wang T,Liu LY,Wang P,et al.Virulent genes of Escherichia coli O157∶H7strains isolated from China[J].Chin J Zoonoses,2010,26(6):515-518.(in Chinese)王濤,劉麗云,王娉,等.我國分離的大腸埃希菌O157∶H7的毒力基因檢測分析[J].中國人獸共患病學報,2010,26(6):515-518.
[12]Hedican EB,Medus C,Besser JM,et al.Characteristics of O157versus non-O157Shiga toxin-producing Escherichia coli infections in Minnesota,2000-2006[J].Clin Infect Dis,2009,49(3):358-364.DOI:10.1086/600302
[13]Aslani MM,Bouzari S.Characterization of virulence genes of non-O157Shiga toxin-producing Escherichia coli isolates from two provinces of Iran[J].Jpn J Infect Dis,2009,62(1):16-19.
[14]Maiden MC,Bygraves JA,Feil E,et al.Multilocus sequence typing:A portable approach to the identification of clones within populations of pathogenic microorganisms[J].Proc Natl Acad Sci U S A,1998,95(6):3140-3145.DOI:10.1073/pnas.95.6.3140
[15]Mellmann A,Bielaszewska M,K?ck R,et al.Analysis of collection of hemolytic uremic syndrome-associated enterohemorrhagic Escherichia coli[J].Emerg Infect Dis,2008,14(8):1287-1290.DOI:10.3201/eid1408.071082
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
