CD4+CD25+Foxp3hi細胞在夏氏瘧原蟲感染DBA/2小鼠作用研究
2012-01-12 06:58:10馮輝潘艷艷李瑩朱曉彤曹雅明
微生物學雜志 2012年2期
關鍵詞:小鼠
馮輝,潘艷艷,李瑩,朱曉彤,曹雅明
(中國醫科大學基礎醫學院免疫學教研室,遼寧沈陽110001)
體內建立適時適度的保護性免疫應答有賴于免疫調節機制的精確調控。大量研究表明,調節性T細胞(regulatory T cell,Treg)在宿主免疫調控網絡中發揮舉足輕重的作用。Treg細胞是一群異質性細胞,從來源分為胸腺來源的特征性表達Foxp3和組成性表達CD25的天然調節性T細胞和在外周誘導產生的Tr1和Th3等。Foxp3因其是天然調節性T細胞在胸腺內發育的重要轉錄因子,對Treg發育及功能發揮起重要作用[1]。鼠瘧或流行區個體均已經發現瘧疾感染會引起Treg數量變化。近期研究發現,利用靶向基因“knockin”法下調小鼠內源性Foxp3基因表達導致Treg的抑制功能缺失[2]。流行區個體對瘧疾感染的抵抗性增強源于Treg功能性缺失[3]。對西非人群瘧疾易感性研究表明,Fulani人群表現為對瘧疾感染抵抗與體內Treg功能相關基因Foxp3和CTLA-4低表達密切相關[3]。同時,利用惡性瘧原蟲感染紅細胞(iRBC)與正常人外周血單個核細胞(PBMC)體外混合培養發現,iRBC能夠以劑量依賴性方式誘導CD4+CD25hiFoxp3hi細胞擴增,并且對炎癥性細胞因子的分泌和保護性免疫應答發揮免疫抑制作用[4]。由此,瘧疾感染Foxp3表達水平與Treg功能密切相關。本研究室曾利用不同鼠瘧模型研究顯示,小鼠對瘧原蟲的易感性與Treg活化密切相關[5-6]。目前,DBA/2小鼠對致死型夏氏瘧原蟲(Plasmodium chabadudi chabadui AS,P.c chabadui AS)呈現易感性的原因尚不明確。為深入探討Treg在瘧疾感染中的作用地位,研究P.c chabadui AS感染DBA/2小鼠Treg活化特點,探討Foxp3表達水平和Treg細胞功能與瘧疾易感性的相互關系。
1 材料與方法
1.1 材料
1.1.1 實驗動物、瘧原蟲6~8周齡DBA/2小鼠,購自中國科學院上海實驗動物中心;P.c chabaudi AS,日本愛媛大學分子寄生蟲學教研室惠贈。
1.1.2 主要試劑以下單克隆抗體(mAb)均購自美國BD Bioscience:抗-CD4-FITC mAb(GK 1.5)、抗-CD25-PE mAb(PC61)和FcγIII/II封閉抗體(2.4G2);抗-Foxp3-APC mAb(FJK-16S)和Foxp3胞內染色試劑盒,購自美國eBioscience公司。
1.2 方法
1.2.1 實驗動物感染和Treg細胞消除模型制備
小鼠經腹腔感染1×106P.c chabaudi AS寄生的紅細胞(pRBC),小鼠經尾靜脈采血,制備薄血膜,Giemsa染色,顯微鏡檢計數紅細胞感染率。小鼠在P.c chabaudi AS感染前1 d、感染后1 d和3 d間隔腹腔注射Anti-CD25mAb(7D4,IgM)1 mg/只/次,構建Treg細胞消除鼠瘧模型。對照組在同一時間點注射等量PBS。
1.2.2 脾CD4+CD25+Foxp3+細胞流式染色無菌取出小鼠脾臟,常規方法制備脾細胞懸液,用0.17 mol/L NH4Cl裂解紅細胞。以含10%胎牛血清(FCS)的RPMI 1640調整脾細胞終濃度為1×107/mL。取0.1 mL脾細胞懸液,預先加入FcγⅢ/Ⅱ封閉抗體1 μg封閉30 min。設陰性對照管,抗-CD4-FITC、抗-CD25-PE和抗-Foxp3-APC單標管。每份樣品同時用抗-CD4-FITC mAb和抗-CD25-PE mAb進行細胞表面雙色標記,4℃孵育
30 min。洗滌1次,棄上清。每管加0.25 mL Foxp3固定透膜劑,4℃孵育30 min。洗滌1次,棄上清。每管再加入抗-Foxp3-APC mAb進行胞內染色,4℃孵育30 min。洗滌1次,棄上清。用0.5 mL FBS/PBS重懸細胞,待流式細胞儀進行檢測。
1.2.3 細胞獲取與分析利用流式細胞儀(FACAria II,美國B&D公司)獲取細胞,使用前向散射角(FSC)及側向散射角(SSC)確定淋巴細胞群。以陰性對照為參考,將對照管所示的非特異熒光的99%以上作為本底扣除,以單標管為對照,調整不同熒光通道補償。每個樣品獲取10 000~50 000個細胞。結果以二維點陣圖顯示。利用FAC expressV3 software分析流式結果。
1.2.4 統計學分析應用SPSS 11.5統計學分析軟件,Student's t test比較分析組內和組間均值的顯著性差異。P小于或等于0.05為差異顯著。
2 結果與分析
2.1 P.c chabaudi AS感染小鼠原蟲血癥和生存率
DBA/2對照組小鼠經腹腔感染1×106P.c chabaudi AS寄生的紅細胞(pRBC),于感染后5 d外周血可見瘧原蟲感染紅細胞,隨后紅細胞感染率迅速上升。在感染后5~7 d,紅細胞感染率從1%升至28%。在感染后8~9 d,紅細胞感染率達到峰值(40.3%)(圖1b),小鼠于感染峰值后全部死亡(圖1a)。在Treg消除組,瘧原蟲感染的紅細胞在外周血中延遲出現(感染后6~7 d),紅細胞感染率在原蟲血癥達峰值前(5~7 d)明顯低于對照組(P<0.01)。原蟲血癥達峰值(37.5%)時間延遲至感染后10 d(圖1b)。隨后,紅細胞感染率陡然下降。在感染后10~15 d從35%降至3%,在感染后20 d瘧原蟲被清除,小鼠生存期明顯延長(圖1a)。
2.2 P.c chabaudi AS感染小鼠CD4+T細胞表面CD25和Foxp3表達水平
與未感染小鼠相比,對照組小鼠感染P.c chabaudi AS后Foxp3表達水平明顯增加。Foxp3平均熒光強度(MFI)顯示,Foxp3表達水平在感染后3d出現上升且維持增高水平,直到感染后10 d,小鼠出現死亡(P<0.01,圖2)。與對照組相比,Treg消除組從感染后3 d,Foxp3表達出現降低,隨后有所回升。但在感染后5~8 d,Foxp3表達水平明顯低于對照組(P<0.01),直到感染后10 d Foxp3表達水平回升至對照組水平(圖2b)。與Foxp3表達水平相比,Treg消除組CD25表達在感染后3 d出現一個短暫性明顯降低(P<0.01),隨后迅速恢復至對照組水平(圖2c)。

圖1 P.c chabaudi AS感染DBA/2小鼠CD25消除和對照組感染過程Fig.1 P.c chabaudi AS infection in DBA/2 control and CD25-depleted mice
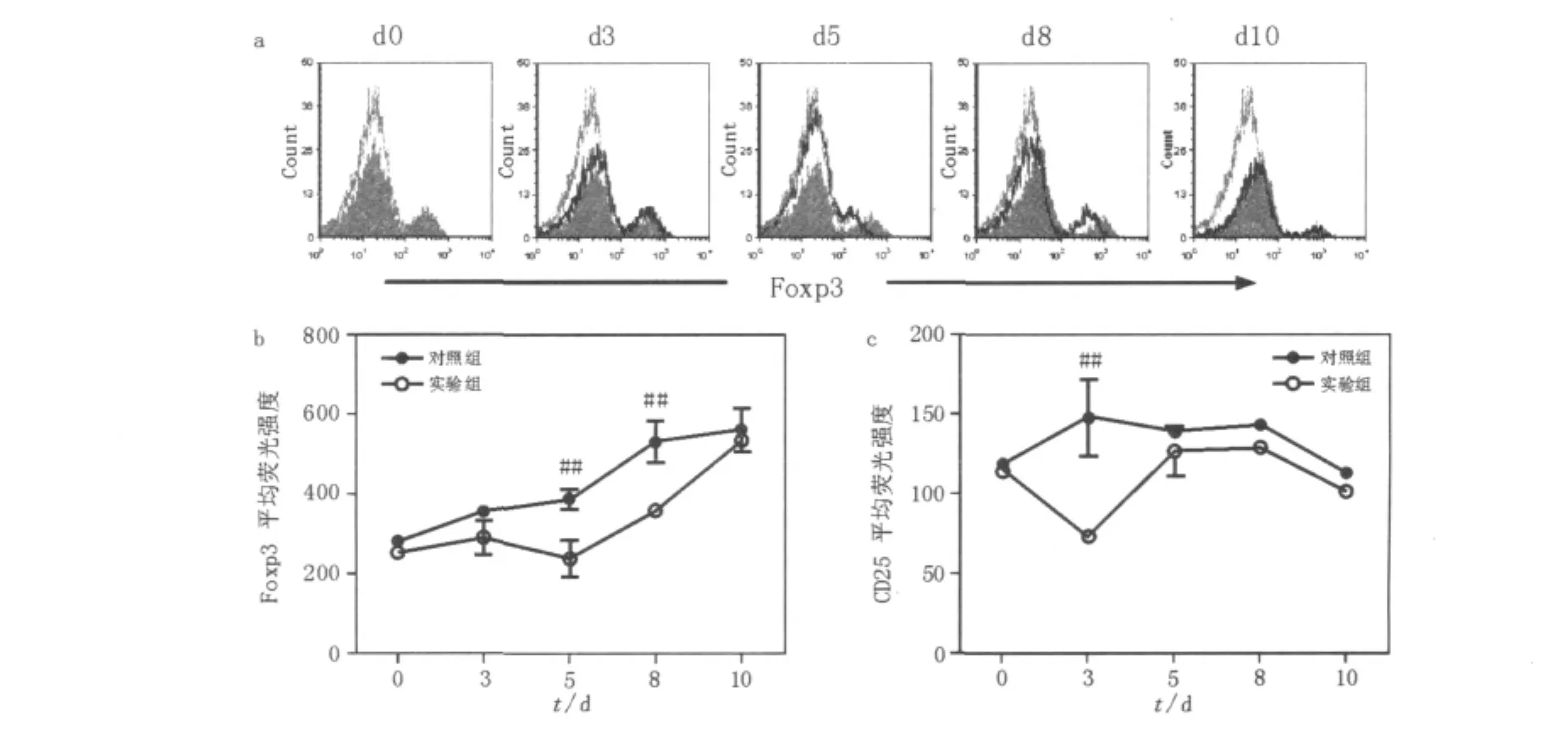
圖2 P.c chabaudi AS感染中小鼠脾臟CD4+T細胞表面CD25和Foxp3表達水平比較Fig.2 The expression levels of Foxp3+on CD25+from CD4+T cells from DBA/2 control and CD25-depleted mice infected with P.c chabaudi AS
2.3 P.c chabaudi AS感染小鼠脾臟CD4+CD25+Foxp3+Treg細胞數量
鑒于P.c chabaudi AS感染后,對照組小鼠脾臟Foxp3表達出現明顯增加。根據Foxp3表達的熒光強度,將高表達Foxp3的Treg細胞單獨劃分為CD4+CD25+Foxp3hi細胞(圖3a)。與未感染小鼠相比,在感染后3 d,對照組CD4+CD25+Foxp3+細胞百分含量和絕對值數量均明顯增加(P<0.01,圖3d),在瘧血癥峰值時CD4+CD25+Foxp3+細胞絕對值達到峰值。CD4+CD25+Foxp3hi細胞的百分含量和絕對值與CD4+CD25+Foxp3+具有相似的變化趨勢。然而,CD4+CD25+Foxp3hi細胞總數比CD4+CD25+Foxp3+細胞增加幅度明顯(圖3b、c、d、e)。與對照組相比,在感染后3~8 d,Treg消除組CD4+CD25+Foxp3hi細胞總數明顯降低(P<0.01,圖3b、c),在感染后10 d出現回升。在感染后3 d,Treg消除組脾臟CD4+CD25+Foxp3hi細胞總數是對照組的30%,隨著對照組CD4+CD25+Foxp3hi細胞總數的增加,Treg消除組CD4+CD25+Foxp3hi細胞所占比例進一步降低。在對照組CD4+CD25+Foxp3hi細胞總數達峰值時,Treg消除組CD4+CD25+Foxp3hi細胞總數是對照組的15%。
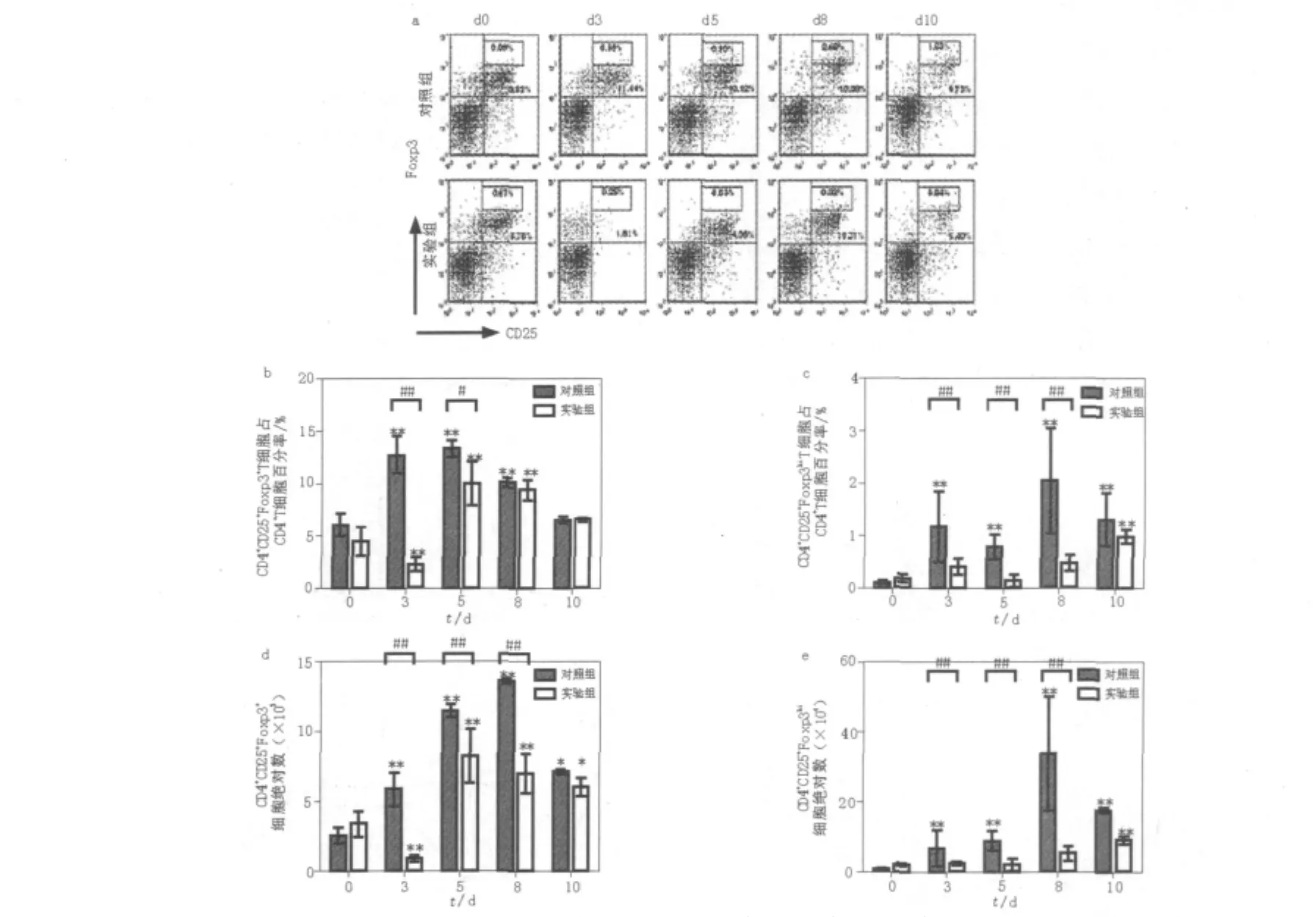
圖3 P.c chabaudi AS感染中小鼠脾臟CD4+CD25+Foxp3+細胞數量比較Fig.3 The percentage and absolute number of splenic CD4+CD25+Foxp3+cells from DBA/2 control and CD25-depleted mice infected with P.c chabaudi AS infection
3 討論
本研究結果顯示,DBA/2小鼠對P.c chabaudi AS感染易感,在原蟲血癥達峰值后小鼠相繼死亡。在感染過程中,DBA/2小鼠CD4+CD25+Foxp3+細胞數量明顯增加,且以CD4+CD25+Foxp3hi增加更為明顯。伴隨著瘧血癥峰值出現CD4+CD25+Foxp3hi細胞數量亦達到峰值。相比,Treg消除鼠的原蟲出現時間和瘧血癥峰值時間均明顯延遲,且在瘧血癥達峰值前(5~8 d)原蟲血癥水平明顯低于對照組。與之相應,CD4+CD25+Foxp3hi細胞數量明顯處于低水平。同時,Treg消除鼠生存期明顯延長。由此提示,P.c chabaudi AS感染導致Foxp3表達增加,擴增的CD4+CD25+Foxp3hi細胞有利于瘧原蟲復制和逃避宿主免疫應答,進而影響瘧疾感染的進程和最終結局。
惡性瘧患者體內Treg的活化與瘧原蟲高水平增殖密切相關[7]。同時,間日瘧原蟲感染中Treg活化數量增高與原蟲負荷增加有關[8]。但同時有研究證實Treg不能對瘧疾急性期感染發揮調控作用[9]。Hiseada等多個研究小組[10-14]證實Treg能夠輔助原蟲逃避宿主免疫監視,Treg可能是宿主對原蟲發生免疫耐受的主要原因。但近期有研究顯示,消除Treg將導致宿主貧血、原蟲負荷增加和病理性炎癥應答等不良癥狀出現[15]。由此可見,瘧疾感染中Treg對宿主發揮免疫調控作用的利弊效應仍存在爭議。
在小鼠模型中,經典消除Treg的方法是利用IgM anti-CD25 antibody 7D4[10]或IgG1 anti-CD25 antibody PC61[8]消除CD25+細胞,最終降低CD4+CD25+Foxp3+細胞數量。2種方法具有各自的優勢,7D4mAb消除Treg細胞速度快但維持時間短,而PC61mAb發揮作用慢維持時間長。本研究利用7D4mAb作用快速的特點來達到短期消除CD25+細胞的效果。結果顯示,7D4mAb對Foxp3高表達的影響非常明顯,能夠顯著降低CD4+CD25+Foxp3hi細胞數量。CD4+CD25+Foxp3hi細胞的免疫抑制功能可能更為顯著。
綜上,P.c chabaudi AS感染導致CD4+CD25+Foxp3hi細胞數量與原蟲復制密切相關。CD4+CD25+Foxp3hi細胞可能輔助原蟲逃避宿主免疫應答,進而影響瘧疾感染的進程和最終結局。因此,充分認識CD4+CD25+Foxp3hi細胞的免疫調控機理,對研制和開發有效的瘧疾疫苗和抗瘧新藥將具有重要的科學意義。
[1] Fontenot JD,Gavin MA,Rudensky AY.Foxp3 programs the development and function of CD4+CD25+regulatory T cells[J].Nat Immunol,2003,4(4):330-336.
[2] Wan YY,Flavell RA.Regulatory T-cell functions are subverted and converted owing to attenuated Foxp3 expression[J].Nature,2007,445(7129):766-770.
[3] Torcia MG,Santarlasci V,Cosmi L,et al.Functional deficit of T regulatory cells in Fulani,an ethnic group with low susceptibility to Plasmodium falciparum malaria[J].Proc Natl Acad Sci U S A,2008,105(2):646-651.
[4] Scholzen A,Mittag D,Rogerson SJ,et al.Plasmodium falciparum-mediated induction of human CD25Foxp3 CD4 T cells is independent of direct TCR stimulation and requires IL-2,IL-10 and TGFbeta[J].PLoS Pathog,2009,5(8):e1000543.
[5] Zheng W,Wang QH,Feng H,et al.CD4+CD25+Foxp3+regulatory T cells prevent the development of Th1 immune response by inhibition of dendritic cell function during the early stage of Plasmodium yoelii infection in susceptible BALB/c mice[J].Folia Parasitol(Praha),2009,56(4):242-250.
[6] Chen G,Liu J,Wang QH,et al.Effects of CD4(+)CD25(+)Foxp3(+)regulatory T cells on early Plasmodium yoelii 17XL infection in BALB/c mice[J].Parasitology,2009,136(10):1107-1120.
[7] Walther M,Tongren JE,Andrews L,et al.Upregulation of TGF-beta,FOXP3,and CD4+CD25+regulatory T cells correlates with more rapid parasite growth in human malaria infection[J].Immunity,2005,23(3):287-296.
[8] Bueno LL,Morais CG,Araujo FF,et al.Plasmodium vivax:induction of CD4+CD25+Foxp3+regulatory T cells during infection are directly associated with level of circulating parasites[J].PLoS One,5(3):e9623.
[9] Walther M,Jeffries D,Finney OC,et al.Distinct roles for FOXP3 and FOXP3 CD4 T cells in regulating cellular immunity to uncomplicated and severe Plasmodium falciparum malaria[J].PLoS Pathog,2009,5(4):e1000364.
[10] Hisaeda H,Maekawa Y,Iwakawa D,et al.Escape of malaria parasites from host immunity requires CD4+CD25+regulatory T cells[J].Nat Med,2004,10(1):29-30.
[11] Long TT,Nakazawa S,Onizuka S,et al.Influence of CD4+CD25+T cells on Plasmodium berghei NK65 infection in BALB/c mice[J].Int J Parasitol,2003,33(2):175-183.
[12] Nie CQ,Bernard NJ,Schofield L,et al.CD4+CD25+regulatory T cells suppress CD4+T-cell function and inhibit the development of Plasmodium berghei-specific TH1 responses involved in cerebral malaria pathogenesis[J].Infect Immun,2007,75(5):2275-2282.
[13] Tetsutani K,Ishiwata K,Ishida H,et al.Concurrent infection with Heligmosomoides polygyrus suppresses anti-Plasmodium yoelii protection partially by induction of CD4(+)CD25(+)Foxp3(+)Treg in mice[J].Eur J Immunol,2009,39(10):2822-2830.
[14] Wu Y,Wang QH,Zheng L,et al.Plasmodium yoelii:distinct CD4(+)CD25(+)regulatory T cell responses during the early stages of infection in susceptible and resistant mice[J].Exp Parasitol,2007,115(3):301-304.
[15] Cambos M,Belanger B,Jacques A,et al.Natural regulatory(CD4+CD25+FOXP+)T cells control the production of proinflammatory cytokines during Plasmodium chabaudi adami infection and do not contribute to immune evasion[J].Int J Parasitol,2008,38(2):229-238.
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國男科學雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
