大鼠心肌素慢病毒RNA干擾載體的構建及其對血管平滑肌細胞分化的影響*
2011-11-20 02:44:35申志華王曉燕王國平敖啟林
中國病理生理雜志 2011年4期
揭 偉, 申志華, 王曉燕, 況 東, 王國平, 敖啟林△
(1廣東醫學院基礎學院病理教研室,廣東 湛江 524023;2華中科技大學同濟醫學院附屬同濟醫院病理研究所,湖北 武漢 430030)
·實驗技術·
大鼠心肌素慢病毒RNA干擾載體的構建及其對血管平滑肌細胞分化的影響*
揭 偉1, 2, 申志華1, 王曉燕2, 況 東2, 王國平2, 敖啟林2△
(1廣東醫學院基礎學院病理教研室,廣東 湛江 524023;2華中科技大學同濟醫學院附屬同濟醫院病理研究所,湖北 武漢 430030)
目的構建大鼠心肌素(myocardin)慢病毒RNA干擾(RNAi)重組質粒,分析該RNAi質粒的干擾效果及其對血管平滑肌細胞(VSMCs)分化的影響。方法設計并合成3對針對大鼠myocardin的shRNA片段,退火后將雙鏈oligo-DNA克隆入慢病毒質粒載體pGCSIL- GFP獲得重組干擾質粒pGCSIL- GFP - shMyocd。以pEGFP - N1 / X124G表達載體構建含Flag標簽的myocardin表達質粒pEGFP - N1- Myocd。將重組干擾質粒和過表達質粒共轉染293T細胞,Western blotting分析Flag標簽蛋白的表達,篩選出干擾效率最好的干擾質粒并大量擴增。干擾質粒轉染大鼠主動脈VSMCs,RT - PCR和Western blotting檢測myocardin及分化標志物SM22α的表達。結果成功構建大鼠myocardin的慢病毒干擾質粒pGCSIL- GFP - shMyocd和過表達質粒pEGFP - N1 - Myocd。上述2種質粒共轉染293T細胞后,1號干擾質粒轉染組Flag標簽蛋白表達下降最顯著。將1號質粒進行大包裝,獲得滴度為1×1012TU/L的慢病毒顆粒,該病毒轉染VSMCs后,myocardin表達明顯下調并伴隨分化標志物SM22α表達的顯著下降。結論成功構建大鼠myocardin慢病毒重組干擾質粒pGCSIL- GFP - shMyocd;抑制基因myocardin表達后伴隨VSMCs分化的表達下降,提示myocardin在VSMCs分化過程中的重要性。pGCSIL- GFP - shMyocd重組質粒的成功構建為后續研究特定病理條件下(如動脈粥樣硬化)VSMCs表型轉變的分子機制奠定了基礎。
心肌素; 載體構建; 慢病毒; 血管平滑肌細胞; 細胞分化
血管平滑肌細胞(vascular smooth muscle cells,VSMCs)是維持血管張力和功能的主要細胞成分,在血管壁病理過程中也發揮重要作用。在特定環境下,VSMCs可以發生表型轉化,即由收縮型轉變為合成型,合成型VSMCs的增殖和遷移能力明顯增強并能分泌細胞外基質,這在動脈粥樣硬化(atherosclerosis, AS)、血管損傷后狹窄等病灶的發生發展中起重要作用。迄今為止VSMCs表型轉變的機制仍未得到有效闡明[1]。在VSMCs的分化發育過程中,一系列標志物順序表達。大部分VSMCs標志基因在其啟動子增強區域都包含1個至多個特異性CArG [CC(A/T)6GG] 序列,這些基因都由一種較廣泛表達的順式結合因子即血清反應因子(serum response factor, SRF)及其輔助激活因子心肌素即myocardin來調控[2]。
Myocardin是Wang[3]等用生物信息學方法從心臟特異性表達基因中發現的SRF的輔助因子。它不與DNA直接結合,而是通過與SRF結合形成三元復合物來調控依賴于CArG的基因轉錄[4]。Myocardin在干/祖細胞向平滑肌細胞分化過程中起重要作用,被認為是調節平滑肌細胞分化的分子開關成分之一[5]。前期我們發現高糖和低氧刺激誘導表型轉變的VSMCs中myocardin表達下降,但具體調控機制尚不清楚[6, 7]。因而基于myocardin分子來探討VSMCs表型轉化具有顯著的理論及潛在的臨床意義。本實驗基于慢病毒表達質粒pGCSIL - GFP構建了針對大鼠myocardin重組干擾載體,轉染VSMCs后觀察myocardin及分化標志物SM22α的表達改變,初步分析myocardin在VSMCs分化過程中作用,為進一步深入探討myocardin在VSMCs表型轉變中的作用的分子機制研究奠定基礎。
材 料 和 方 法
1材料
1.1主要試劑 質粒pGCSIL- GFP、pHelper 1.0和pHelper 2.0,大腸桿菌菌株 DH5α和293T細胞由上海吉凱生物技術有限公司提供;pEGFP - N1 / X124G質粒和DNA ladder由上海捷瑞公司提供;質粒大量抽提試劑盒購自Qiagen;AgeI、EcoR I、KpnI、T4 DNA連接酶及buffer購自NEB;BCA試劑盒購自HyClone - Pierce;ECL-Plu sKit購自Amersham;DMEM培養基、胎牛血清和DMSO購自Gibco;Lipofectamine 2000和Opti - MEM購自Invitrogen;羊抗鼠myocardin、兔抗鼠IgG - HRP和兔抗羊IgG - HRP購自Santa Cruz;鼠抗Flag購自Sigma;小鼠抗β-actin購自碧云天。
2方法
2.1Myocardin慢病毒干擾質粒構建 根據大鼠myocardin mRNA(NM_182667)序列信息,結合設計經驗和設計軟件 (http://hydra1.wistar.upenn.Edu/Projects/siRNA/siRNAindex.htm )進行評估測定,經 BLAST表明不與大鼠其它 cDNA序列同源,合成3對干擾DNA序列和1對陰性對照(NC)序列(表1),其兩端含酶切位點黏端。1號重組質粒靶序列:GAT GCA TTT GCC TTT GAA GAT;2號重組質粒靶序列:GAC ACA ATC CAG GAT CTC ACT;3號重組質粒靶序列:CAG AAA GTG ACA AGA ACG ATA;重組陰性對照(NC)質粒靶序列(scrambled sequence):TTC TCC GAA CGT GTC ACG T。將表1中的寡核苷酸各5 μL,緩沖液20 μL 和70 μL雙蒸水混合,90 ℃ 4 min,70 ℃10 min,冷卻至室溫后經12 %非變性PAGE凝膠檢測雙鏈形成效率,T4噬菌體DNA連接酶將雙鏈DNA 連接入經AgeI和EcoR I雙酶切線性化的慢病毒pGCSIL- GFP載體,轉化E.coliDH5α感受態細胞,陽性克隆經PCR和測序(ABI 3730,Invitrogen)鑒定。PCR上游引物5’- CCT ATT TCC CAT GAT TCC TTC ATA - 3’,下游引物5’- GTA ATA CGG TTA TCC ACG CG - 3’。陽性克隆進行質粒抽提,最后質粒DNA測定濃度后-20 ℃保存備用。
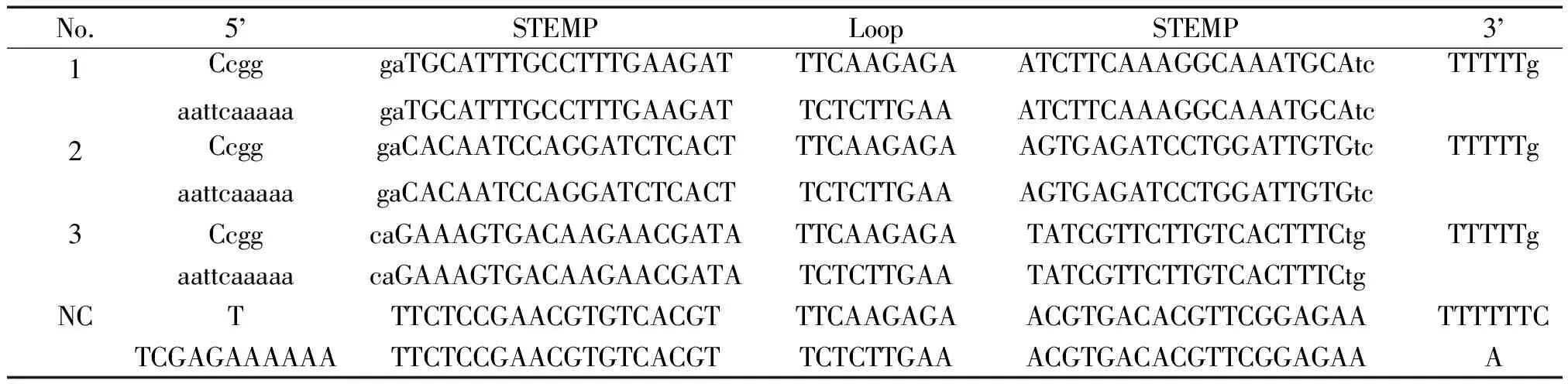
表1 病毒載體干擾序列
2.2Myocardin-GFP融合蛋白過表達質粒構建 大鼠myocardin mRNA CDS位于244-3 060 nt處,整個CDS區域序列較長,因此獲得整個CDS區域表達質粒較困難。我們選擇性人工合成一段從244-1 023 nt的RNA序列,該合成序列包含上述3對干擾序列作用靶點。將合成序列克隆入經EcoR II、KpnI雙酶切的pEGFP-N1 / X124G質粒從而構建pEGFP - N1 - Myocd過表達質粒,表達蛋白質與GFP形成融合蛋白。將pEGFP - N1 - Myocd過表達質粒用Lipofectamine 2000轉染293T細胞,轉染48 h后,熒光顯微鏡下觀察融合蛋白GFP的表達情況,判斷pEGFP - N1 - Myocd過表達質粒構建成功與否。
2.3有效myocardin干擾靶點質粒篩選 生長良好的293T細胞按1×108cells/L 接種于24孔板,80 % - 90 %融合時換成400 μL Opti- MEM。細胞轉染時,每孔表達載體的量均為0.5 μg,干擾質粒量分為0.25 μg和0.5 μg,Lipofectamine 2000體積均為1 μL。將質粒和Lipofectamine 2000分別溶解于Opti- MEM中,混勻,室溫靜置5 min,將稀釋好的質粒和Lipofectamine 2000混勻,室溫靜置20 min,把質粒DNA與Lipofectamine 2000的混合液加入293T細胞中,37 ℃、5% CO2培養箱中培養8 h,換成新鮮的含10 %胎牛血清的完全培養液,轉染后24 h熒光顯微鏡下觀察預估轉染率,48 h后收集細胞,抽提總蛋白進行Western blotting檢測。
2.4Myocardin干擾載體慢病毒擴增及滴度測定 對數生長期293T 細胞以含10 %胎牛血清的培養基調整細胞密度為 6×108cells/L,接種于15 cm培養皿,常規培養24 h后達 70 % - 80 %融合可用于轉染。轉染前2 h換為無血清培養基。向一滅菌離心管中加入所制備的各DNA溶液(pGCSIL - Myocd 載體 20 μg,pHelper 1.0 載體 15 μg,pHelper 2.0 載體10 μg),與相應體積的 Opti- MEM 混合均勻,總體積為 2.5 mL,在室溫下溫育5 min。取 100 μL Lipofectamine 2000 試劑在另一管中與 2.4 mL Opti- MEM 混合,室溫下溫育 5 min。稀釋后的 DNA與稀釋后的Lipofectamine 2000 進行混合,室溫下溫育 20 min,以便形成 DNA與Lipofectamine 2000稀釋液的轉染復合物。將DNA與Lipofectamine 2000混合液轉移至293T細胞的培養液中,混勻,常規培養8 h后棄去培養基,加入20 mL PBS洗滌。加入含10 %血清的細胞培養基 25 mL,繼續常規培養 48 h,收集上清,于 4℃ 4 000 ×g離心10 min除去細胞碎片,0.45μm濾器過濾上清。把病毒粗提液加入到過濾杯中,蓋上蓋子,將過濾杯插到濾過液收集管中,在 4 000 ×g離心10 min - 15 min到需要的病毒濃縮體積,病毒分裝后置于-80 ℃保存備用。取出1支病毒用逐孔稀釋法測定病毒滴度。換算公式為:滴度 (103TU/L ) = 觀察到帶有熒光的細胞個數/病毒原液量。
2.5大鼠主動脈VSMCs慢病毒質粒轉染及效果分析 雄性SD大鼠(250 g)由華中科技大學同濟醫學院實驗動物中心提供。組織貼塊法分離主動脈VSMCs并用α-平滑肌肌動蛋白免疫組化染色鑒定,實驗用第3代。VSMCs接種24孔板后約70%融合時,按MOI = 120加入pGCSIL- GFP - shMyocd,每孔培養基總體積250 μL(含4 mg/L polybrene 1μL),轉染8 h后更換新鮮培養液,48 h后收獲細胞,提取蛋白和RNA,分別用RT - PCR和Western blotting 檢測myocardin和SM22α的表達。
2.6mRNA表達分析 Trizol試劑抽提總RNA,2 μg RNA逆轉錄為cDNA。應用半定量PCR檢測目的基因mRNA表達。Myocardin上游引物5’- GGG TCT GAA CAC TCT TTG C - 3’, 下游引物5’- ATT ACC GTG GAG GCT TGG A - 3’,產物大小254 bp;SM22α上游引物5’- AGC CAG TGA AGG TGC CTG AGA AC - 3’,下游引物5’- TGC CCA AAG CCA TTA GAG TCC TC - 3’,產物大小181 bp;β- actin上游引物5’- CAT TGT TGC CCA TCA ACG ACC - 3’,下游引物5’- TCA CAC CCA TCA CAA ACA TG - 3’,產物大小173 bp。PCR退火溫度53 ℃。引物均由Invitrogen公司合成。產物經1.5 %瓊脂糖凝膠電泳,紫外燈下觀察并拍照(Gel Doc1 000, Bio - Rad)。實驗重復3次。
2.7Western blotting 冰上裂解細胞15 min后將樣品轉移入EP管中,超聲破碎儀破碎細胞(200 W共4次,每次5 s,間隔2 s),4 ℃ 12 000×g離心15 min,經BCA法測蛋白濃度后調整樣品蛋白終濃度為2 g/L。蛋白經12 % SDS - PAGE后轉移到PVDF膜,5 %脫脂牛奶的TBST溶液室溫封閉PVDF膜1 h,Ⅰ抗4 ℃過夜孵育,TBST洗膜3次,每次10 min,Ⅱ抗室溫下孵育2 h,TBST洗膜3次,每次10 min,ECL試劑顯色后拍照。應用抗體為:小鼠抗Flag(F1804,1∶5 000),小鼠抗β-actin(AA128,1∶ 5 000),羊抗鼠IgG - HRP(sc-2005,1∶8 000),羊抗鼠myocardin(sc-21561,1∶500),兔抗羊IgG- HRP (sc-2030,1∶8 000)。
3統計學處理
結 果
1成功構建慢病毒干擾質粒
將合成的3對針對myocardin不同靶點序列和1對陰性對照雙鏈oligo-DNA連接入經AgeI和EcoR I雙酶切pGCSIL - GFP線性化慢病毒載體質粒,經氨芐西林抗性基因篩選,挑選出陽性克隆經PCR(圖1)和測序驗證(未發表資料)無誤,證實合成的oligo-DNA正確連接入線性化pGCSIL- GFP載體;連接入shRNA片段的陽性克隆PCR片段大小為343 bp,沒有連接入vshRNA片段的空載體克隆PCR片段大小為306 bp。電泳鑒定其大小在預期范圍,證實成功構建了慢病毒干擾質粒,標記為pGCSIL- GFP - shMyocd和pGCSIL - GFP - NC。
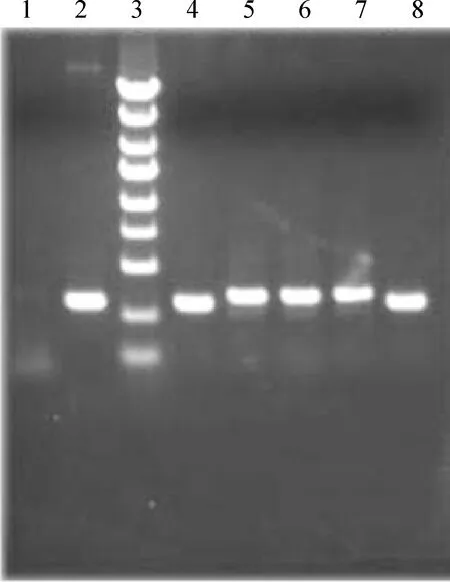
Figure 1. Electrophoresis of positive clones of recombinant pGCSIL-GFP vector. lane 1: ddH2O;lane 2: pGCSIL - GFP;lane 3: marker (5.0 kb, 3.0 kb, 2.0 kb, 1.5 kb, 1.0 kb, 750 bp, 500 bp, 250 bp, 100 bp);lane 4-8: positive clones.
2成功構建myocardin-Flag-GFP融合蛋白過表達質粒
pEGFP - N1 / X124G質粒經EcoR I和KpnI酶切后去除X124G過渡序列,接入myocardin(244 nt - 1 023 nt)和Flag表達序列后構建pEGFP - N1 - Myocd真核表達質粒,表達myocardin - Flag - GFP融合蛋白。該過表達載體轉染293T細胞后,熒光顯微鏡下觀察到綠色熒光,證實過表達載體正常工作,見圖2。
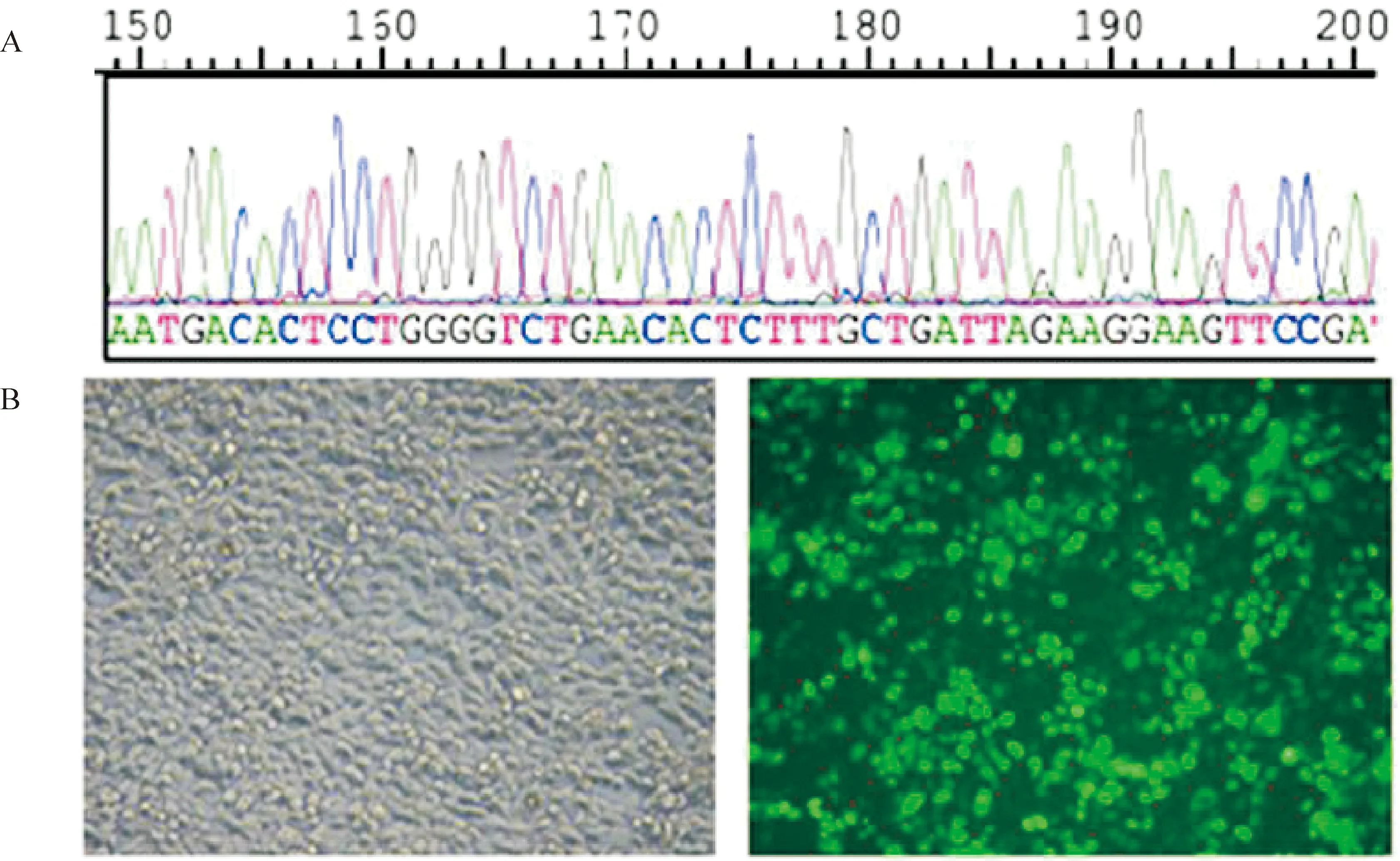
Figure 2. Construction and transfection of myocardin-overexpression vector.A: sequencing result of partial myocardin CDS (244 nt - 284 nt);B: green fluorescent was observed in 293T cells transfected with myocardin-overexpression vectors.
3有效干擾靶點慢病毒鑒定及擴增
將不同靶點pGCSIL - GFP - shMyocd載體和pEGFP - N1 - Myocd過表達載體共轉染293T細胞,熒光顯微鏡下觀察綠色熒光表達來判斷轉染效率;轉染成功后的細胞經提取蛋白,Western blotting檢測Flag的表達來判斷干擾載體的敲減效應,結果證實設計的3個靶點中2個具有明顯基因表達敲減效應,見圖3。我們選擇性包裝了1號質粒(#1)和陰性對照質粒(NC),其滴度分別達到1×1012TU/L和5×1012TU/L。
4慢病毒干擾質粒降低VSMCs中myocardin和收縮基因表達
將包裝的1號慢病毒干擾質粒轉染VSMCs后,RT - PCR和Western blotting結果均顯示pGCSIL - GFP - shMyocd質粒顯著降低VSMCs中myocardin表達,同時伴隨VSMCs收縮標志物SM22α mRNA和蛋白表達水平的顯著下降,見圖4。
討 論
慢病毒RNA干擾系統是1型人類免疫缺陷病毒(human immunodeficiency virus type 1,HIV-1)為基礎改造的病毒基因載體質粒,載體轉入細胞后轉錄出的RNA鏈折疊成具有發夾結構的shRNA。shRNA發夾結構被Dicer切除后產生siRNA,在細胞內產生基因沉默。慢病毒載體不但可以感染非分裂期細胞,也可以感染分裂期細胞,還具有容納外源性目的基因片段大、穩定整合持久表達、免疫反應小等優點[8]。慢病毒干擾系統是目前應用較多的一種RNAi體系。為探討myocardin在VSMCs表型轉變中的表達及作用,我們遵從RNAi設計的一般原則[9-12],設計并合成了針對大鼠myocardin 3個不同的靶點shRNA序列和1個陰性對照shRNA序列,并利用BLAST進行查詢,確定其為特異性序列,合成的DNA兩端設計有AgeI或EcoR I 酶切位點及黏端。將合成的雙鏈shRNA通過DNA連接酶連接入慢病毒干擾空載體(pGCSIL - GFP),經抗性基因篩選,PCR和克隆測序證實了目的序列正確連接入克隆。
為驗證設計的慢病毒干擾載體能正常工作,我們進一步構建了一個myocardin的過表達載體。Flag是一個由十幾個氨基酸組成的蛋白標簽,由目的蛋白和標簽(tag)共同構成的蛋白嵌合體,常標記于重組蛋白質的氨基端以幫助分析靶蛋白的生物學功能,用于目的細胞暫時不易得到或目的細胞轉染效率非常低基因靶點的篩選[11]。鑒于myocardin整個CDS堿基近3 000 bp,完整CDS克隆很難得到,根據文獻[13,14],我們將Flag標記引入myocardin過表達質粒。我們將一段長800多bp的CDS序列(已經包含設計的3條干擾質粒序列)合成并克隆入經EcoR I和KpnI雙酶切線性化pEGFP - N1 / X124G表達載體,該表達載體再接上Flag標簽,合成后的過表達載體pEGFP - N1 -myocardin形成myocardin - GFP融合蛋白并表達Flag。合成后的表達質粒經轉染293T工具細胞后,熒光顯微鏡下見到明顯的綠色熒光,證實該過表達質粒正常工作。進一步將此myocardin過表達質粒與不同干擾位點的慢病毒干擾質粒共轉染工具細胞,Western blotting檢測工具細胞內Flag的表達情況,結果證實設計的3條針對myocardin的慢病毒干擾質粒有2個位點(#1和#3)具有明顯的敲減效應,敲減效率gt;75%。我們選擇了#1質粒進行慢病毒的大量包裝,最后得到滴度分別為1×1012TU/L和5×1012TU/L的pGCSIL - GFP - shMyocd和對照lentivirus - GFP - NC病毒載體。
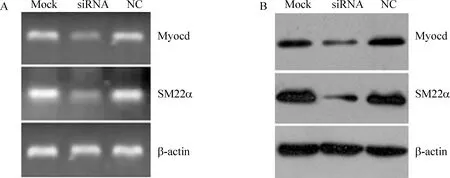
Figure 4. Myocardin lentiviral RNAi vector attenuated myocardin expression in VSMCs accompanied with the decrease of SM22α expression.A: RT - PCR;B: Western blotting. Mock: pGCSIL - GFP empty vector;siRNA: pGCSIL - GFP - shMyocd;NC: pGCSIL - GFP - NC.
將#1干擾病毒轉染VSMCs后,發現其顯著下調VSMCs中myocardin的表達,同時伴隨VSMCs分化標志物SM22α的表達顯著下降。提示該myocardin慢病毒干擾質粒能正常工作。SM22α是VSMCs典型的收縮標志物,其啟動子區域含多個CArG序列。Myocardin與SRF形成三聯復合物后,能調控具有CArG序列心肌細胞和平滑肌細胞基因的表達[3]。當共轉染外源性SRF和myocardin過表達質粒后,VSMCs中包括SM22α在內的收縮標志物表達明顯增加[15],在骨髓間充質干細胞向VSMCs誘導分化過程中也伴隨myocardin及SM22α在內的收縮標志物表達的增加[16],因而提示myocardin在VSMCs分化過程起重要作用。我們前期的結果也證實這一點[6,7]。
VSMCs是一種表型可塑性細胞。在動脈粥樣硬化、血管損傷后內膜增生、低氧性肺動脈重塑等血管疾病中VSMCs由收縮表型轉化為合成表型。鑒于myocardin在VSMCs分化中的重要作用,預示著基于myocardin靶點對涉及VSMCs表型轉化性血管疾病如動脈粥樣硬化等血管重塑性疾病進行分子治療的可能性。本研究中有關pGCSIL- GFP - shMyocd重組質粒的構建為進一步的研究奠定了基礎。
[1] Owens GK, Kumar MS, Wamhoff BR. Molecular regulation of vascular smooth muscle cell differentiation in development and disease [J]. Physiol Rev, 2004,84(3):767-801.
[2] Yoshida T, Owens GK. Molecular determinants of vascular smooth muscle cell diversity [J]. Circ Res, 2005,96(3):280-291.
[3] Wang D, Chang PS, Wang Z, et al. Activation of cardiac gene expression by myocardin, a transcriptional cofactor for serum response factor [J]. Cell, 2001,105(7): 851-862.
[4] Wang Z, Wang DZ, Hockemeyer D, et al. Myocardin and ternary complex factors compete for SRF to control smooth muscle gene expression [J]. Nature, 2004,428(6979): 185-189.
[5] Chen J, Kitchen CM, Streb JW, et al. Myocardin: a component of a molecular switch for smooth muscle differentiation [J]. J Mol Cell Cardiol,2002,34(10):1345-1356.
[6] Jie W, Wang X, Zhang Y, et al. SDF-1α/CXCR4 axis is involved in glucose- potentiated proliferation and chemotaxis in rat vascular smooth muscle cells [J]. Int J Exp Pathol, 2010, 91(5):436-444.
[7] Jie W, Guo J, Shen Z, et al. Contribution of myocardin in the hypoxia-induced phenotypic switching of rat pulmonary arterial smooth muscle cells [J]. Exp Mol Pathol, 2010, 89(3):301-306.
[8] 主鴻鵠, 徐開林. 慢病毒載體的改進及其在血液病基因治療中的應用[J]. 中國實驗血液學雜志,2003,11(2):208-212.
[9] Berezhna SY, Supekova L, Supek F, et al. siRNA in human cells selectively localizes to target RNA sites [J]. Proc Natl Acad Sci USA,2006,103(20): 7682-7687.
[10]Matveeva O, Nechipurenko Y, Rossi L, et al. Comparison of approaches for rational siRNA design leading to a new efficient and transparent method [J]. Nucleic Acids Res, 2007,35(8):e63.
[11]Kurachi S, Koizumi N, Sakurai F, et al. Characterization of capsid- modified adenovirus vectors containing heterologous peptides in the fiber knob, protein IX, or hexon[J]. Gene Ther,2007,14(3): 266-274.
[12]陳 艷,邵建永,吳秋良,等. 線粒體細胞色素C氧化酶RNAi慢病毒載體的構建[J]. 中國病理生理雜志,2005, 21(6):1117-1123.
[13]史艷俠, 韓文杰, 彭柔君,等. 使用FLAG標簽肽及慢病毒載體共同篩選小鼠foxp3基因RNA干擾的有效靶點[J]. 中山大學學報(醫學科學版),2007,28(6): 641-644.
[14]Cordelier P, Strayer DS. Using gene delivery to protect HIV- susceptible CNS cells: inhibiting HIV replication in microglia [J]. Virus Res,2006,118(1-2): 87-97.
[15]許麗輝, 何 強. Myocardin在大鼠血管平滑肌細胞表型轉化中的作用機制[J].中國老年病學雜志,2010,30(17):2483-2486.
[16]李 燕,瞿智玲,黃 冠,等. Myocardin在骨髓間充質干細胞向平滑肌細胞分化過程中的表達[J]. 中華病理學雜志,2008,37(10):680-686.
ConstructionofalentiviralRNAinterferencevectortargetingratmyocardinanditseffectonvascularsmoothmusclecelldifferentiation
JIE Wei1, 2, SHEN Zhi-hua1, WANG Xiao-yan2, KUANG Dong2, WANG Guo-ping2, AO Qi-lin2
(1DepartmentofPathology,SchoolofBasicMedicalScience,GuangdongMedicalCollege,Zhanjiang524023,China;2InstituteofPathology,TongjiHospital,HuazhongUniversityofScienceamp;Technology,Wuhan430030,China.E-mail:Aoqilin@126.com)
AIM: To construct a lentiviral RNA interference(RNAi)vector targeting rat myocardin mRNA and to investigate its effect on the differentiation of vascular smooth muscle cells(VSMCs).METHODSThree pairs of dsDNA targeting rat myocardin mRNA were designed, synthesized and cloned into lentiviral vector pGCSIL-GFP to generate pGCSIL-GFP-shMyocd lentvirus. A Flag-tagged myocardin-overexpression vector pEGFP-N1-Myocd was constructed with pEGFP-N1/X124G. After these two vectors were cotransfected into 293T cells, the flag protein was assessed by Western blotting to analyze the knockdown effect of pGCSIL-GFP-shMyocd. The expression of myocardin and SM22α was also detected by RT-PCR and Western blotting after the pGCSIL-GFP-shMyocd viruses were transfected into primary cultured rat aortal VSMCs.RESULTSThe rat myocardin lentviral RNAi vector pGCSIL-GFP-shMyocd and myocardin-overexpression vector pEGFP-N1-Myocd were successfully constructed. After these two kinds of vectors were cotransfected into 293T cells,the No.1 interfering vector displayed the highest inhibitory effect on flag expression.After the No.1 lentvirus at the titer of 1×1012TU/L was transfected into VSMCs, the myocardin and SM22α expression was significantly attenuated.CONCLUSIONThe lentiviral pGCSIL-GFP-shMyocd RNAi vector is successfully constructed, which is useful for further study regarding the molecular mechanism of the phenotypic switching in VSMCs under special pathological conditions such as atherosclerosis. Inhibition of myocardin expression in VSMCs leads to the decrease in the expression of differentiation marker, and implies a crucial role of myocardin in VSMCs differentiation.
Myocardin; Vector construction; Lentivirus; Vascular smooth muscle cells; Cell differentiation
1000-4718(2011)04-0823-06
R329.2
A
10.3969/j.issn.1000-4718.2011.04.040
2010-09-30
2011-01-19
國家自然科學基金資助項目(No.30400192)
△通訊作者Tel:027-83650551;E-mail: Aoqilin@126.com
