小鼠神經(jīng)元中CCK8與EGF協(xié)同作用的初步研究*
2011-11-20 02:41:54劉曉燕梅志強(qiáng)
中國病理生理雜志 2011年4期
關(guān)鍵詞:小鼠
李 娟, 王 麗, 陳 熔, 劉曉燕, 梅志強(qiáng), 何 濤
(瀘州醫(yī)學(xué)院醫(yī)學(xué)分子生物學(xué)實(shí)驗(yàn)室,四川 瀘州 646000)
小鼠神經(jīng)元中CCK8與EGF協(xié)同作用的初步研究*
李 娟, 王 麗, 陳 熔, 劉曉燕, 梅志強(qiáng), 何 濤△
(瀘州醫(yī)學(xué)院醫(yī)學(xué)分子生物學(xué)實(shí)驗(yàn)室,四川 瀘州 646000)
目的探討八肽膽囊收縮素(CCK8)與表皮生長因子(EGF)在神經(jīng)元信號(hào)轉(zhuǎn)導(dǎo)中的交叉對(duì)話(cross-talk)及可能存在機(jī)制,并對(duì)CCK8與EGF的協(xié)同作用進(jìn)行初步研究。方法CCK8刺激表皮生長因子受體(EGFR)磷酸化的受體類型分析:培養(yǎng)的乳小鼠神經(jīng)元隨機(jī)分為對(duì)照組(等量的DMEM培養(yǎng)基)、CCK8刺激組(10-7mol/L)、CCKA受體拮抗劑L364 718組、CCKB受體拮抗劑L365 260組以及L364 718+L365 260聯(lián)合組(濃度均為10-8mol/L),拮抗劑加入10 min后再給予CCK8,作用5 min后收集細(xì)胞提取蛋白,Western blotting檢測(cè)磷酸化表皮生長因子受體(p-EGFR)蛋白表達(dá)量的變化;CCK8與EGF對(duì)神經(jīng)元EGFR磷酸化狀態(tài)的影響和CCK8與EGF協(xié)同作用功效分析:培養(yǎng)的乳小鼠神經(jīng)元隨機(jī)分為對(duì)照組、CCK8(10-7mol/L)組、EGF(40 μg/L)組以及CCK8+EGF組,藥物作用5 min后終止反應(yīng),處理和分析方法同前,同時(shí)通過MTT法檢測(cè)各組神經(jīng)元培養(yǎng)不同時(shí)間(24、48、72、96 h)后細(xì)胞的生長活性。結(jié)果(1)2種CCK受體拮抗劑均可降低EGFR蛋白的磷酸化水平,其中A型受體拮抗劑的作用明顯強(qiáng)于B型,2種拮抗劑同時(shí)作用對(duì)EGFR蛋白磷酸化水平的降低作用更為明顯;(2)CCK8和EGF分別刺激神經(jīng)元后,EGFR的磷酸化水平均增強(qiáng),但CCK8+EGF刺激后表現(xiàn)更為顯著;(3)CCK8與EGF聯(lián)用對(duì)增強(qiáng)神經(jīng)元生長活性和延長其存活時(shí)間的作用較兩藥單獨(dú)作用效果更為明顯(Plt;0.05)。結(jié)論(1)CCKA和CCKB受體均介導(dǎo)了CCK8激活EGFR磷酸化的信號(hào)轉(zhuǎn)導(dǎo)途徑,但A型受體發(fā)揮了更為重要的作用;(2)CCK8與EGF信號(hào)轉(zhuǎn)導(dǎo)途徑之間存在著cross-talk;(3)CCK8與EGF可能通過信號(hào)轉(zhuǎn)導(dǎo)的cross-talk協(xié)同促進(jìn)小鼠神經(jīng)元的生長與存活。
膽囊收縮素; 神經(jīng)元; 受體, 表皮生長因子
膽囊收縮素 (cholecystokinin, CCK)作為一種典型的腦腸肽,在神經(jīng)系統(tǒng)功能調(diào)節(jié)中起著重要的作用,其在神經(jīng)元中的信號(hào)轉(zhuǎn)導(dǎo)將對(duì)了解腦的結(jié)構(gòu)與功能、闡明某些中樞神經(jīng)系統(tǒng)疾病的發(fā)病機(jī)理具有重要意義,因而近年來已成為備受關(guān)注的領(lǐng)域。目前研究表明,CCK具有神經(jīng)保護(hù)作用,是一種內(nèi)源性的神經(jīng)保護(hù)因子,但有關(guān)CCK在神經(jīng)系統(tǒng)的作用機(jī)制還不明了。本研究組前期研究結(jié)果顯示, 八肽膽囊收縮素(cholecystokinin octapeptide,CCK8)刺激神經(jīng)元后,表皮生長因子受體(epidermal growth factor receptor,EGFR)底物15磷酸化水平增高,進(jìn)一步的分析表明,CCK8能促進(jìn)EGFR磷酸化水平顯著升高,提示CCK與表皮生長因子(epidermal growth factor,EGF)之間可能存在著復(fù)雜的“信息相互交談”(cross-talk)[1]。本研究在此基礎(chǔ)上擬通過CCK受體抑制實(shí)驗(yàn)分析培養(yǎng)的小鼠神經(jīng)元中介導(dǎo)EGFR磷酸化水平改變的受體類型,初步探討CCK8與EGF在神經(jīng)元信號(hào)轉(zhuǎn)導(dǎo)中的相互作用,并觀察CCK8與EGF協(xié)同作用的功能效應(yīng),為闡明CCK在中樞神經(jīng)系統(tǒng)中的作用提供重要線索。
材 料 和 方 法
1材料
1.1動(dòng)物 昆明種封閉群乳小鼠(SPF級(jí)),由瀘州醫(yī)學(xué)院實(shí)驗(yàn)動(dòng)物中心提供。
1.2主要實(shí)驗(yàn)試劑 Neurobasal-A medium+B27 supplement購自Gibco;CCK8購自Sigma;EGF購自Peprotech EC;磷酸化EGFR(phospho-EGFR,p-EGFR)抗體購自CST;抗β-actin抗體、彩色預(yù)染蛋白質(zhì)分子量標(biāo)準(zhǔn)和Western封閉液均購自碧云天生物技術(shù)研究所;HRP標(biāo)記羊抗兔IgG購自北京中杉;SuperSignal? West Pico Chemiluminescent Substrate購自Millipore;CCK受體抑制劑L364 718和L365 260由第三軍醫(yī)大學(xué)西南肝膽研究中心馬寬生教授饋贈(zèng);其它所用試劑為國產(chǎn)或進(jìn)口分析純。
2方法
2.1神經(jīng)元的培養(yǎng) 取出生1-2 d的小鼠腦組織消化得到單細(xì)胞,加入DMEM/F12培養(yǎng)基(含10%胎牛血清)以2×106細(xì)胞接種于培養(yǎng)瓶內(nèi),置于5% CO2、37 ℃培養(yǎng),24 h后更換為含Neurobasal-A medium和B27supplement的培養(yǎng)液,以后每 2-3 d換液1次。
2.2CCK受體拮抗劑處理神經(jīng)元 將培養(yǎng)神經(jīng)元隨機(jī)分為對(duì)照組、實(shí)驗(yàn)組以及受體拮抗劑組。對(duì)照組給予等量DMEM培養(yǎng)基,實(shí)驗(yàn)組給予CCK8(10-7mol/L)。受體拮抗劑組又分為3組,先分別加入CCKA受體拮抗劑L-364 718、CCKB受體拮抗劑L365 260以及L-364 718+ L365 260,拮抗劑濃度均為10-8mol/L,孵育10 min后,再加入10-7mol/L CCK8孵育5 min,液氮終止反應(yīng)。
2.3CCK8及EGF處理神經(jīng)元 培養(yǎng)至第7 d的乳小鼠神經(jīng)元隨機(jī)分為對(duì)照組、CCK8(10-7mol/L)組、EGF(40 μg/L)組以及CCK8+EGF組,加入對(duì)應(yīng)的藥物作用5 min后,傾去培養(yǎng)基,液氮終止反應(yīng)。
2.4Western blotting檢測(cè)EGFR磷酸化水平 提取神經(jīng)元蛋白,Bradford法測(cè)蛋白含量,SDS-PAGE電泳后將凝膠上的蛋白轉(zhuǎn)移至PVDF膜上, Western封閉液作用1 h,按Western blotting常規(guī)操作進(jìn)行p-EGFR蛋白的檢測(cè),最后SuperSignal化學(xué)發(fā)光檢測(cè)試劑進(jìn)行顯影,凝膠化學(xué)發(fā)光儀器內(nèi)采集發(fā)光圖像,同時(shí)檢測(cè)β-actin的表達(dá)作為內(nèi)參照。Gel-Pro Analyzer 4.0凝膠成像軟件進(jìn)行圖像分析,結(jié)果以累積吸光度比值(IAp-EGFR/IAβ-actin)表示。
2.5細(xì)胞形態(tài)學(xué)觀察及MTT法測(cè)定細(xì)胞活性 培養(yǎng)24 h的神經(jīng)元分組情況同方法2.3,各組細(xì)胞每天在倒置顯微鏡下做形態(tài)學(xué)觀察。同時(shí)96孔培養(yǎng)板內(nèi)的各組細(xì)胞在加入對(duì)應(yīng)藥物分別作用24、48、72、96 h后,每孔加入MTT(1.5 g/L)液20 μL,37 ℃孵育4 h,吸去培養(yǎng)液,每孔加入100 μL DMSO,室溫振蕩10 min,酶聯(lián)免疫檢測(cè)儀上于490 nm處測(cè)量各孔的吸光度值(A)。實(shí)驗(yàn)重復(fù)3次,每組實(shí)驗(yàn)設(shè)3復(fù)孔。
3統(tǒng)計(jì)學(xué)處理
結(jié) 果
1不同CCK受體拮抗劑對(duì)CCK8刺激EGFR磷酸化的影響
比較對(duì)照組、實(shí)驗(yàn)組以及CCK受體拮抗劑組的印跡圖譜(圖1)可見,CCK8刺激后神經(jīng)元EGFR蛋白磷酸化狀態(tài)發(fā)生明顯改變。軟件分析結(jié)果顯示,對(duì)照組有p-EGFR蛋白條帶的出現(xiàn)(1.153±0.012),說明神經(jīng)元具有p-EGFR的基礎(chǔ)活性;CCK8刺激神經(jīng)元后 p-EGFR蛋白條帶明顯加深(1.761±0.046),說明CCK8可使EGFR磷酸化水平明顯升高,激活其活性;加入CCKA受體特異拮抗劑L364 718后,EGFR的磷酸化水平明顯降低(0.345±0.023),加入CCKB受體特異拮抗劑L365 260后,EGFR的磷酸化水平也出現(xiàn)降低,但是作用較L364 718弱些(0.577±0.037);而同時(shí)使用2種抑制劑可導(dǎo)致EGFR蛋白磷酸化水平降低更為明顯(0.192±0.013)。以上各組與對(duì)照組相比,差異均顯著(Plt;0.05)。

Figure 1. Detection of phosphorylated EGFR in neurons with different treatments by Western blotting. ±s.n=3. *Plt;0.05 vs control group.
2CCK8、EGF、CCK8+EGF刺激后EGFR磷酸化的變化
由圖2可見,藥物作用組EGFR磷酸化水平與對(duì)照組比均有明顯增高,其中p-EGFR與內(nèi)參照β-actin的累積吸光度比值在CCK8+EGF組表達(dá)量最高(在對(duì)照組、CCK8組、EGF組以及CCK8+EGF組比值分別為0.161±0.015、0.210±0.020、0.356±0.021和0.394±0.010),組間比較有顯著差異(Plt;0.05)。

Figure 2. Detection of phosphorylated EGFR with different stimuli by Western blotting. ±s.n=3. *Plt;0.05 vs control group.
3CCK8與EGF的協(xié)同作用對(duì)神經(jīng)元生長及活性的影響
3.1神經(jīng)元形態(tài)觀察 對(duì)照組神經(jīng)元培養(yǎng)至7 d時(shí),生長達(dá)到頂峰狀態(tài),具有典型的神經(jīng)元特性,胞體周圍有明亮的光暈,突起明顯并交織成網(wǎng)絡(luò)狀結(jié)構(gòu);培養(yǎng)至第9 d,神經(jīng)元貼壁數(shù)量相對(duì)減少,部分神經(jīng)元胞體變大變圓,突起斷裂,立體感消失,細(xì)胞呈現(xiàn)老化趨勢(shì),見圖3A;培養(yǎng)至第13 d,神經(jīng)元貼壁數(shù)量明顯減少,培養(yǎng)液中可見懸浮細(xì)胞,而仍然貼壁的神經(jīng)元胞體腫脹,細(xì)胞突起消失,神經(jīng)元之間的網(wǎng)絡(luò)松散,瓶底鋪滿大量膠原,細(xì)胞已老化死亡。藥物組在加入CCK8和EGF刺激劑后培養(yǎng)至第9 d時(shí),細(xì)胞生長依然旺盛,胞體豐滿,折光性強(qiáng),表現(xiàn)出較好的生長狀態(tài),見圖3B;培養(yǎng)至第13 d,神經(jīng)元才開始出現(xiàn)部分神經(jīng)元胞體變大變圓、突起消失,細(xì)胞開始老化。
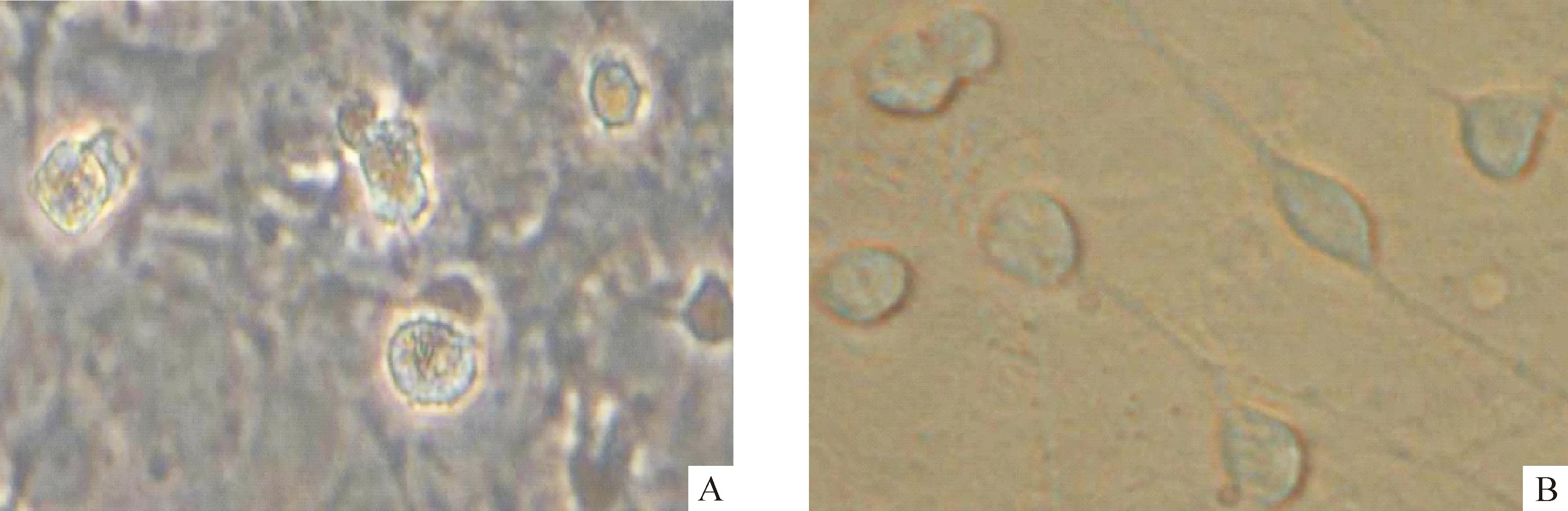
Figure 3. Neurons in control group(A) and CCK8+EGF stimulation group(B) after 9-day culture(×400).
3.2MTT法檢測(cè)細(xì)胞活性 神經(jīng)元培養(yǎng)24-96 h后,MTT測(cè)定結(jié)果顯示,對(duì)照組細(xì)胞活性基本一致;3組加藥組的神經(jīng)元在加入刺激劑培養(yǎng)24 h后,細(xì)胞活性開始增強(qiáng),并隨著時(shí)間的延長,活性增強(qiáng)越來越明顯,至96 h時(shí),細(xì)胞活性達(dá)最高峰,呈現(xiàn)時(shí)間依賴性。與對(duì)照組比,CCK8+EGF組的神經(jīng)元活性增強(qiáng)最為顯著,見圖4。

Figure 4. The growth curves of the neurons treated with CCK8, EGF and CCK8+EGF. ±s.n=9. *Plt;0.05 vs control;△Plt;0.05 vs CCK8 or EGF.
討 論
CCK發(fā)揮生理作用首先需要與靶細(xì)胞膜上的特異受體結(jié)合,在中樞神經(jīng)系統(tǒng)CCK受體主要有2種亞型,即CCKA和CCKB受體,兩者的分布和介導(dǎo)的功能各有不同,可能是通過不同信號(hào)轉(zhuǎn)導(dǎo)機(jī)制。國外有研究報(bào)道,在胃上皮細(xì)胞中,胃泌素通過CCKB/胃泌素受體介導(dǎo)的信號(hào)途徑激活EGFR酪氨酸磷酸化[2];Piiper等[3]也在AR42J細(xì)胞中發(fā)現(xiàn)CCK可激活EGFR的活性,引起酪氨酸磷酸化,具體機(jī)制不明,可能與ERK1/2的活化有關(guān),提示CCK可能是通過CCKB受體介導(dǎo)的MAPK信號(hào)轉(zhuǎn)導(dǎo)途徑激活EGFR磷酸化。上述結(jié)果顯示在胃腸細(xì)胞中,CCK與EGF信號(hào)系統(tǒng)之間存在著相互作用,且CCKB受體介導(dǎo)的信號(hào)轉(zhuǎn)導(dǎo)途徑可能參與其中,但在神經(jīng)系統(tǒng)中二者是否具有信號(hào)轉(zhuǎn)導(dǎo)的交叉作用還未見相關(guān)報(bào)道。
本實(shí)驗(yàn)通過CCK特異性受體拮抗劑抑制實(shí)驗(yàn),分析CCK8刺激EGFR磷酸化的受體類型,結(jié)果表明神經(jīng)元中CCK的2種類型受體都介導(dǎo)了CCK8激活EGFR磷酸化的信號(hào)轉(zhuǎn)導(dǎo)途徑,但CCKA受體介導(dǎo)的信號(hào)通路占主導(dǎo)作用。這一結(jié)果與已報(bào)道的胃腸細(xì)胞中的研究結(jié)果不一致,其原因可能與神經(jīng)系統(tǒng)與胃腸道系統(tǒng)的組織特異性有關(guān)。
神經(jīng)元作為一種終末分化細(xì)胞,無增殖能力,一旦損傷或破壞,將會(huì)永久消失,因此如何增強(qiáng)神經(jīng)元細(xì)胞活性,延長其存活時(shí)間一直是腦神經(jīng)科學(xué)的研究熱點(diǎn)。近年來,CCK在延緩機(jī)體老化進(jìn)程中所起的作用引起了廣泛關(guān)注。Sun等[4]應(yīng)用NBA2細(xì)胞無血清培養(yǎng)老化模型,觀察應(yīng)用一定濃度的CCK8后,細(xì)胞突起和微突起數(shù)量、脂褐素?zé)晒庵涤忻黠@的改變,推測(cè)一定濃度的CCK8在適當(dāng)作用時(shí)間下可以延緩老化的進(jìn)展。Voits等[5]研究提示CCK在延緩神經(jīng)細(xì)胞老化的過程中發(fā)揮了一定的作用。而EGF作為一種神經(jīng)營養(yǎng)因子,在神經(jīng)系統(tǒng)中具有重要功能,可促進(jìn)神經(jīng)系統(tǒng)的生長發(fā)育、神經(jīng)干細(xì)胞的增殖與分化,并參與神經(jīng)元的氧化性損傷等。Wagner等[6]發(fā)現(xiàn)皮質(zhì)神經(jīng)元的存活可能依賴EGFR介導(dǎo)的信號(hào)轉(zhuǎn)導(dǎo)途徑。本研究通過用免疫印跡法檢測(cè)到在神經(jīng)元中CCK8可促進(jìn)EGFR磷酸化,且CCK8與EGF聯(lián)用可產(chǎn)生一定的協(xié)同作用,表明CCK8與EGF在神經(jīng)元中的信號(hào)轉(zhuǎn)導(dǎo)確實(shí)存在cross-talk。隨后我們用CCK8、EGF分別及協(xié)同刺激神經(jīng)元,通過觀察神經(jīng)元的形態(tài)及細(xì)胞活性的變化而進(jìn)一步了解兩者間這種信號(hào)轉(zhuǎn)導(dǎo)的cross-talk是否具有功能效應(yīng)。研究結(jié)果表明,CCK8和EGF單獨(dú)作用均可增強(qiáng)神經(jīng)元活性,延長神經(jīng)元的存活時(shí)間,但CCK8與EGF協(xié)同作用更為顯著地增強(qiáng)神經(jīng)元的活性。對(duì)于CCK8如何促進(jìn)EGFR磷酸化的機(jī)制有待進(jìn)一步深入研究,但CCK與神經(jīng)營養(yǎng)因子EGF之間通過信號(hào)轉(zhuǎn)導(dǎo)的交叉對(duì)話,協(xié)同發(fā)揮對(duì)神經(jīng)元的保護(hù)作用為神經(jīng)系統(tǒng)退行性疾病的治療、干細(xì)胞分化后神經(jīng)元活性的維持等提供了重要的實(shí)驗(yàn)依據(jù)。
[1] 何 濤,李 娟,王 麗,等. CCKB受體誘發(fā)的小鼠神經(jīng)元蛋白質(zhì)磷酸化的研究[J]. 中國病理生理雜志, 2008, 24(1):47-50.
[2] Miyazaki Y, Shionmura Y, Tsutsui S, et al. Gastrin induces heparin-binding epidermal growth factor-like growth factor in rat gastric epithelial cells transfected with gastrin receptor[J]. Gastroenterology, 1999, 116(1):78-89.
[3] Piiper A, Elez R, You SJ, et al. Cholecystokinin stimulates extracellular signal-regulated kinase through activation of the epidermal growth factor receptor, Yes, and protein kinase C.Signal amplification at the level of Raf by activation of protein kinase Cε[J].J Biol Chem, 2003, 278(9): 7065-7072.
[4] Sun XJ, Lu QC, Cai Y. Effect of cholecystokinin on experimental neuronal aging[J]. World J Gastroenterol, 2005, 11 (4): 551-556.
[5] Voits M, Hasen?hrl RU, Huston JP,et al. Repeated treatment with cholecystokinin octapeptide improves maze perfomance in aged Fischer 344 rats[J]. Peptides,2001, 22(8):1325-1330.
[6] Wagner B, Natarajan A, Grünaug S, et al. Neuronal survival depends on EGFR signaling in cortical but not midbrain astrocytes[J]. EMBO J, 2006, 25(4): 752-762.
CrosstalkofCCK8andEGFinmouseneurons
LI Juan, WANG Li, CHEN Rong, LIU Xiao-yan, MEI Zhi-qiang, HE Tao
(MolecularBiologyLaboratory,LuzhouMedicalCollege,Luzhou646000,China.E-mail:Hetao198@163.com)
AIM: To explore the mechanism of signaling transduction and cross talk between cholecystokinin octapeptide (CCK8) and epidermal growth factor (EGF) in mouse neurons and to observe the effect of CCK8in coordination with EGF on neuron growth and cell viability.METHODSFor determining which kind of CCK receptor mediated the phosphorylation of EGF receptor, the cultured neurons were randomly divided into control group, CCK8stimulation group, CCKAreceptor antagonist group, CCKBreceptor antagonist group, and CCKA+CCKBreceptor antagonist group. Control and stimulation groups were stimulated with DMEM and CCK8(10-7mol/L) for 5 min, respectively, while antagonist groups were pre-incubated with different types of receptor antagonists (10-8mol/L) for 10 min and followed by stimulating the neurons with CCK8. For observing the effect of CCK8and EGF on the phosphorylation of EGFR in neurons and on neuron growth and cell viability, the cultured neurons were randomly divided into control group, CCK8stimulation group, EGF stimulation group and CCK8+EGF stimulation group, which were stimulated with DMEM, CCK8(10-7mol/L), EGF (40 μg/L) and CCK8+EGF for 5 min, respectively. Reactions were terminated by freezing the neurons in liquid nitrogen and the phosphorylated EGFR was detected by Western blotting. Meanwhile, the viability of the neurons was observed by MTT method after stimulated for 24 h, 48 h, 72 h and 96 h.RESULTSThe phosphorylation levels of EGFR were decreased in the neurons treated with either of the two CCK receptor antagonists, and more obvious decrease was observed when the two CCK receptor antagonists were used in combination. Compared with control group, the phosphorylation levels of EGFR in the neurons were significantly increased(Plt;0.05) after stimulated with CCK8or EGF, and the increase was more remarkable in CCK8+EGF stimulation group. CCK8or EGF improved the viability and prolonged the life span of the neuron, and synergism of these two reagents was observed.CONCLUSIONBoth CCKAand CCKBreceptors are involved in the phosphorylation of EGFR in the neurons stimulated by CCK8, and the type A receptor may play a more important role. There is cross-talk between CCK8and EGF signaling pathways in neurons. The signaling cross-talk between CCK8and EGF may be the underlying molecular mechanism responsible for the synergistic effect on the neuron growth and viabilityinvitro.
Cholecystokinin; Neurons; Receptors, epidermal growth factor
1000-4718(2011)04-0779-04
R322.81
A
10.3969/j.issn.1000-4718.2011.04.030
2010-08-03
2010-12-27
四川省教育廳資助項(xiàng)目(No.2006A053)
△通訊作者 Tel: 0830-3161283;E-mail: Hetao198@163.com
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
