FAK-shRNA重組逆轉錄病毒載體的構建及穩定表達細胞株的篩選*
2011-11-20 02:41:45阿力亞
中國病理生理雜志 2011年4期
關鍵詞:肝癌
阿力亞, 林 武, 何 強
(中山大學附屬第一醫院肝膽外科, 廣東 廣州 510080)
FAK-shRNA重組逆轉錄病毒載體的構建及穩定表達細胞株的篩選*
阿力亞, 林 武, 何 強△
(中山大學附屬第一醫院肝膽外科, 廣東 廣州 510080)
目的研究利用RNA干擾技術,以黏附斑激酶(FAK)為靶基因,構建FAK-shRNA重組逆轉錄病毒載體,將其導入包裝細胞Phoenix中,篩選出穩定產生FAK-shRNA病毒的細胞克隆,以病毒上清感染并篩選FAK表達沉默的細胞株,觀察其對相關蛋白表達的影響。方法體外合成能轉錄產生靶向FAK短發夾RNA(shRNA)的寡核苷酸并定向克隆入pSuper.retro逆轉錄病毒載體,以脂質體法轉染Phoenix細胞株,待篩選穩定克隆成功后收獲病毒上清,感染人肝癌細胞株HCC-LM3,以嘌呤霉素篩選得到抑制FAK表達的穩定細胞株后用Western boltting鑒定FAK表達的抑制效果及相關蛋白表達情況。結果構建了重組逆轉錄病毒載體pSuper-FAK并抑制了人肝癌HCC-LM3細胞內FAK蛋白的表達。在下調FAK表達的細胞株中p-Akt和p-MAPK1/2表達明顯受到抑制。下調FAK的細胞株遷移和侵襲能力下降,細胞周期多被阻止在G0/G1期,細胞凋亡增多,增殖率明顯下降。結論重組逆轉錄病毒載體pSuper-FAK轉染包裝細胞Phoenix后,其產生的FAK-shRNA病毒可以抑制HCC-LM3細胞內的FAK蛋白表達并抑制Akt及MAPK1/2磷酸化。下調FAK后可以對腫瘤細胞的生物學行為產生明顯影響。
黏著斑激酶; RNA干擾; 逆轉錄病毒載體; 肝腫瘤
黏著斑激酶(focal adhesion kinase,FAK)是一種非受體酪氨酸激酶,它在細胞間及細胞與細胞外基質黏附中起關鍵作用。研究證明,FAK在乳腺癌、宮頸癌、直腸癌、甲狀腺癌、前列腺癌、喉癌、舌癌和肺癌中表達量增高,高表達FAK通常與差的病理分型和預后相聯系,并且已經在一些實體腫瘤如乳腺癌中探索了其活化機制以及其通過下游信號轉導通路參與細胞周期調控、黏附、侵襲、轉移等的作用方式。但是對于肝癌中FAK介導腫瘤細胞生物學活性的相關研究甚少,目前尚不清楚FAK在肝癌細胞侵襲、轉移、增殖、生存、凋亡過程中所處地位。本實驗以肝癌細胞株為研究對象,構建質粒轉染并篩選出穩定低表達FAK的細胞株,為后續研究打下研究基礎。
材 料 和 方 法
1材料
1.1主要試劑 逆轉錄病毒載體pSuper.retro由廣東省人民醫院病理生理研究部惠贈,限制性內切酶BamHⅠ、HindⅢ、T4 DNA連接酶購自TaKaRa;大腸桿菌菌株E.coliDH5α由廣東省人民醫院病理生理研究部惠贈;脂質體Lipofectamine2000購自Invitrogen;質粒提取試劑盒購自Tiangen;質粒純化試劑盒購自Qiagen;上、下游引物由上海博尚生物有限公司合成。FAK兔抗人抗體購自Santa Cruz,Akt、p-Akt、MAPK1/2、p-MAPK1/2抗體購自Cell Signal;Transwell小室和培養瓶購自Corning;細胞培養基DMEM/F12和胎牛血清購自Gibco;嘌呤霉素購自Sigma;matrigel 購自BD。
1.2細胞株 人肝癌細胞株HCC-LM3由廣東省人民醫院病理生理研究部惠贈;包裝病毒細胞株Phoenix 購自Orbigen。
2方法
2.1逆轉錄病毒載體的設計及合成 根據GenBank數據庫提供的FAK基因序列(GI:439874),按shRNA的設計原則設計目的shRNA序列, BLAST對比分析。構建質粒3’端酶切位點之前加入Pol III聚合酶終止信號TTTT,以Loop(9 nt的莖環結構)相連,兩端分別加入酶切位點BamHⅠ和HindⅢ,正義鏈5’-GATCCGCCACCTGGGCCAGTATTATTTCAAGACGATAATACTGGCCCAGGTGGTTTTTTGTCGACA-3’,反義鏈3’-GCGGTGGACCCGGTCATAATAAAGTTCTGCTATTATGACCGGGT-CCACCAAAAAACAGCTGTTCGA-5’。
2.2逆轉錄病毒載體的構建 將合成的正、反義鏈經過變性、復性后形成雙鏈FAK-shRNA,定向克隆入逆轉錄病毒載體pSuper.retro后,轉化大腸桿菌菌株E.coliDH5α,以氨芐青霉素(100 mg/L)篩選出陽性克隆,提取質粒。
2.3重組質粒FAK-shRNA 的鑒定 重組質粒用BamHⅠ與HindⅢ酶進行雙酶切,酶切產物行0.8%瓊脂糖凝膠電泳。選取陽性克隆純化后送到上海博尚生物公司測序鑒定,命名為FAK-pSuper-shRNA。
2.4重組質粒轉染包裝病毒細胞株 將包裝病毒細胞Phoenix接種到6孔板中,1×105cells/well,完全培養基培養12 h后更換無血清DMEM/F12培養1 h,分別使用空載體pSuper.retro、pSuper-FAK質粒各5 μg,脂質體Lipofectamine2000 6 μL /well轉染Phoenix。轉染6 h后更換完全培養基,48 h后加入1 mg/L嘌呤霉素,篩選3-4周獲得陽性克隆。細胞株命名為293T-Phoenix-pSuper-FAK及293T-Phoenix-pSuper-control。
2.5重組逆轉錄病毒感染HCC-LM3細胞株 將篩選獲得的Phoenix陽性克隆培養至90%融合后換成無嘌呤霉素的完全培養基,24 h后收集細胞上清。HCC-LM3細胞株接種到6孔板中,每孔加入1 mL Phoenix細胞上清濾液, 37 ℃、5%CO2條件下繼續培養24 h,更換含1 mg/L嘌呤霉素的完全培養基篩選,每3-4 d換液,約40 d后篩選完成。所產生的細胞株分別命名為LM3-pSuper-FAK及LM3-pSuper-control。
2.6Western blotting檢測FAK蛋白的沉默及Akt、p-Akt、MAPK1/2、p-MAPK1/2表達情況 將篩選后的HCC-LM3細胞株接種到6孔板中,培養至每孔1×105細胞時,收集細胞。用細胞裂解液裂解,Bradford蛋白濃度測定試劑盒檢測蛋白濃度,上樣量25 μg總蛋白, 10% SDS聚丙烯酰胺凝膠電泳,將蛋白質轉移到PVDF 膜,用含5%脫脂奶粉的T-TBS 37 ℃封閉1 h,分別加入兔抗人FAK、Akt、p-Akt、MAPK1/2和p-MAPK1/2 單克隆抗體(1∶1 000 稀釋, Santa Cruz)或兔抗人GAPDH單克隆抗體(1∶8 000稀釋,Santa Cruz) 4 ℃孵育過夜。T-TBS漂洗5 min×3次;兔抗辣根過氧化物酶標記Ⅱ抗(1∶7 000稀釋,Santa Cruz)4 ℃孵育45 min; T-TBS漂洗5 min×3次;SuperSignal West Femto(Pierce)敏感曝光試劑盒曝光底片顯影。
2.7細胞遷移實驗 在Transwell下室的DMEM培養液中加入10%血清,在Transwell上室分別滴加1×105LM3-pSuper-FAK及LM3-pSuper-control細胞懸液(無血清)。將上室置于下室之中, 在37 ℃溫箱孵育24 h后取出上室,以棉簽將殘留在上室內表面上的細胞輕輕拭去。上室用PBS洗滌后,遷移至上室膜外上的細胞用0.1%結晶紫染色,以清水洗3遍,顯微鏡下(×100)計算遷移細胞數, 結果取5個視野細胞計數的平均值。
2.8細胞侵襲實驗 操作按Corning公司Transwell書明書進行,鋪matrigel按照BD公司說明書進行。在Transwell上室分別滴加5×104LM3-pSuper-FAK及LM3-pSuper-control細胞懸液(無血清)。在37 ℃溫箱孵育36 h后取出上室,以棉簽將殘留在上室內表面上的細胞輕輕拭去。上室用PBS洗滌后,遷移至上室膜外上的細胞用0.1%結晶紫染色,以清水洗3遍,顯微鏡下(×100)計算遷移細胞數, 結果取5個視野細胞計數的平均值。
2.9流式細胞儀檢測 LM3-pSuper-FAK及LM3-pSuper-control細胞的細胞周期和凋亡 將LM3-pSuper-FAK及LM3-pSuper-control細胞分別制成單個細胞懸液,70%預冷乙醇固定, PBS洗滌,再用碘化丙啶(propidium iodide,PI)處理備檢。用ProfileⅡ型流式細胞儀,在488 nm 激發波長下測定細胞DNA,分析細胞周期及檢測細胞早期凋亡情況。
3.0MTT檢測 LM3-pSuper-FAK及LM3-pSuper-control細胞的細胞增殖率將LM3-pSuper-FAK、LM3-pSuper-control及未處理的LM3細胞消化,調整細胞懸液濃度,細胞計數后每孔加入200 uL鋪96孔板使待測細胞調密度至1 000 cells/well,設置凋零孔(培養基、MTT、二甲基亞砜)及空白對照孔(培養液、二甲基亞砜),邊緣孔用PBS填充。37 ℃培養箱孵育48 h后,每孔加入20 uL MTT溶液4 h,每孔加入150 uL二甲基亞砜,避光置于搖床上低速振蕩10 min,使結晶物充分溶解。在酶聯免疫檢測儀A570處測量各孔的吸光度值。
3統計學處理
結 果
1測序鑒定重組質粒
將提取的重組質粒送交上海博尚生物公司測序,測序結果發現目的片段成功連接于載體中,見圖1。

Figure 1. The DNA sequence of a recombinant.
2Westernblotting檢測FAK蛋白的沉默
經Western blotting檢測分析表明,內參為均一顯影的條帶,FAK顯影條帶密度不同,見圖2。未處理組及對照組FAK蛋白表達量與LM3-pSuper-FAK組比差異顯著, LM3-pSuper-FAK 組蛋白表達量明顯減少。
3沉默FAK對LM3中Akt、p-Akt、MAPK1/2、p-MAPK1/2表達的影響
經Western blotting檢測分析表明,下調FAK后可見p-Akt和p-MAPK1/2蛋白表達明顯減少,Akt及MAPK1/2未見明顯改變,見圖3。
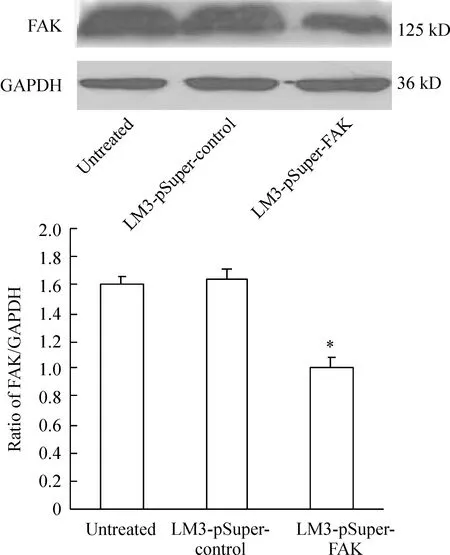
Figure 2. Western blotting analysis of FAK in HCC-LM3 cells±s.n=3.*Plt;0.05 vs LM3-pSuper-control group.
4下調FAK表達對LM3細胞遷移侵襲的影響
經過Transwell 實驗檢測分析表明,下調FAK后可見LM3-pSuper-FAK組遷移率及侵襲能力明顯下降。遷移至下室的細胞數目對照組為(52.12±11.38) cells/視野,實驗組為(22.67±4.36) cells/視野,見圖4。穿過metrigel的細胞數對照組為(23.53±4.54)cells/視野,實驗組為(5.34±0.73)cells/視野,兩組比較差異顯著(Plt;0.05),見圖5。
5下調FAK表達對LM3細胞的細胞周期和凋亡影響
經過流式細胞儀檢測分析表明,LM3-pSuper-FAK組凋亡細胞數占總檢測細胞數的(6.00±1.23)%,LM3-pSuper-control組為(0.88±0.12)%,兩者比較差異顯著(Plt;0.01)。LM3-pSuper-FAK G0/G1期比率是(64.0±2.4)%,相對LM3-pSurper-control組(44.8±1.6)%,有更多的細胞阻滯在G0/G1期,S期細胞數量明顯減少,見圖6。
6下調FAK對LM3細胞增殖率的影響
MTT實驗結果表明(圖7),下調FAK表達的人肝癌細胞LM3-pSuper-FAK、LM3-pSuper-control細胞的增殖抑制率分別為(27.97±0.34)%和(5.45±0.22)%,兩組相比差異顯著(Plt;0.01)。
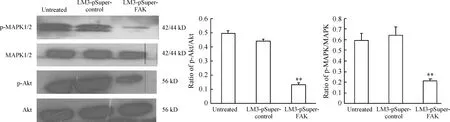
Figure 3. Western blotting analysia the expression of Akt,p-Akt,MAPK and p-MAPK protein in HCC-LM3 cells. ±s. n=3. **Plt;0.01 vs LM3-pSuper-control group.
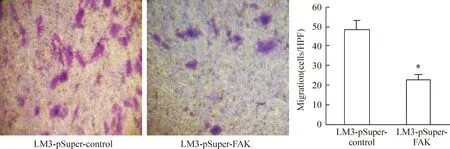
Figure 4. LM3-pSuper-FAK and LM3-pSuper-control cells were added into the upper chamber of the Transwell chamber without matrigel.After 24 h 37 ℃ incubation, the cells in the lower chamber were counted(crystal violet staining,×100).±s. n=5. *Plt;0.05 vs LM3-pSuper-control group.
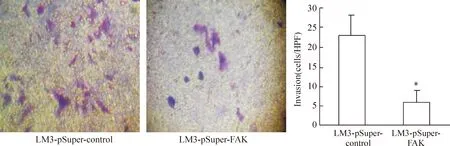
Figure 5. LM3-pSuper-FAK and LM3-pSuper-control cells were added into the upper chamber of the Transwell chambe with matrigel.After 36 h 37℃ incubation, the cells in the lower chamber were counted(crystal violet staning,×100).±s. n=5. *Plt;0.05 vs LM3-pSuper-control group.

Figure 6. Cell cycle and apoptosis tested by flow cytometry±s. n=3.*Plt;0.05, ** Plt;0.01 vs LM3-pSuper-control group.
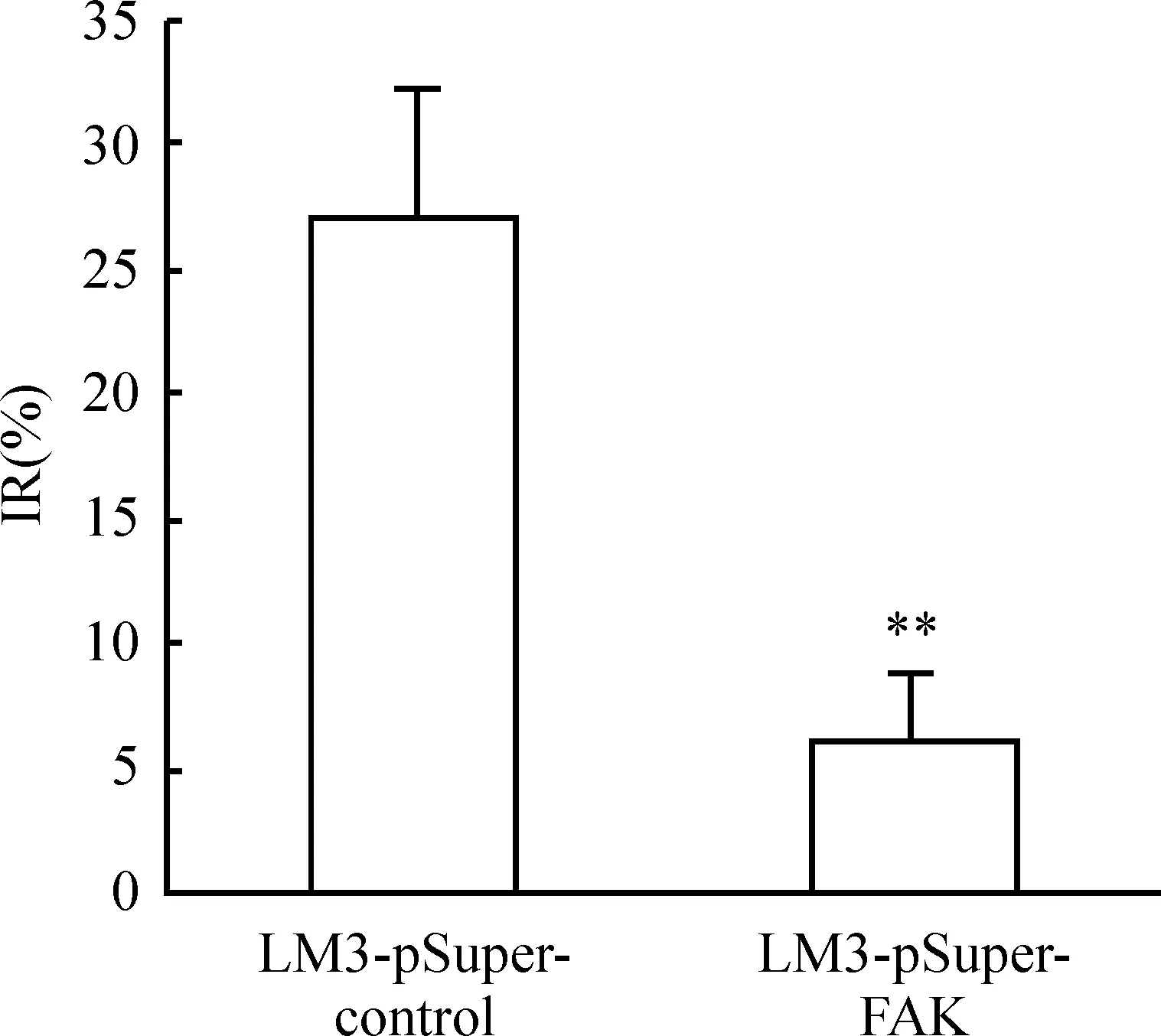
Figure 7. The proliferation of LM-3 cells after knowndown of FAK. .n=3.**Plt;0.01 vs LM3-pSuper-control group.
討 論
黏著斑激酶(FAK)在腫瘤的遷移、增殖、生存、侵襲、轉移和血管形成及白血病等起著重要的調控作用[1-6]。本課題探討了應用RNA干擾(RNA interference, RNAi)技術對FAK基因進行沉默并篩選出穩定表達細胞株,并觀察其對肝癌細胞相關蛋白表達的影響。
RNAi是指在生物體細胞內,外源性或內源性的雙鏈RNA 引起與其同源mRNA的特異性降解,抑制其相應基因的表達,其作為一種新興的逆向遺傳研究方法,與傳統的基因療法相比具有高效、高特異性及無毒副作用等特點[7]。構建shRNA表達載體基于具有合適啟動子的載體或轉錄元件,在細胞體內經過酶切持續生成大量的siRNA,其抑制目的基因的效果與合成siRNA的相似,但更加穩定。逆轉錄病毒載體能將外源基因有效地整合到宿主細胞基因組中,并隨細胞分裂而傳給后代,獲得包裝細胞株后可以穩定的得到含有目的載體的病毒顆粒,而且未經包裝時為缺陷性病毒,不具有完整功能,只有經包裝細胞系為其提供結構蛋白組分,二者相互補償后才能包裝成具有完整功能的病毒顆粒,因此感染靶細胞后不能擴增,具有很高的安全性[8]。
本實驗構建了針對FAK基因的shRNA表達載體FAK-pSuper-shRNA,并成功篩選穩定表達細胞株,得到不同抑制率的FAK陽性克隆,以Western blotting觀察FAK蛋白表達量的變化,結果顯示FAK-pSuper-shRNA重組質粒的干擾組能明顯降低FAK蛋白的表達。相對使用化合物或反義寡核苷酸等方法,利用病毒介導的RNAi更具效率,且可以得到持續傳代的穩定表達,有利于保持實驗數據的穩定性。在進行細胞株篩選的過程中我們也發現,對于人肝癌細胞株LM3,下調FAK后其生長緩慢,我們猜測相對于瞬時轉染,在篩選穩定表達細胞株時除去FAK本身對細胞增殖能力的影響外,還有細胞內環境在適應FAK低表達的過程中需要調動其他信號通路來補償FAK的功能的原因,那么可能在這種穩定表達細胞株繼續檢測敲除FAK對細胞內信號通路影響時,實驗結果會和瞬時轉染或利用特異性抑制劑所產生的結果存在區別,有研究以瞬時轉染進行RNA干擾,同樣可以較高效率抑制FAK蛋白表達[9],但是穩定表達細胞株因其具有實驗結果穩定性,可以為今后的研究打下基礎。
Akt又稱蛋白激酶B(protein kinase B,PKB),是一種絲氨酸/蘇氨酸激酶,在細胞存活途徑中起重要作用。Akt的活化形式是磷酸化,其有2個磷酸化位點,只有當2個位點完全磷酸化后才能使Akt完全活化。Akt作為PI3K/Akt 信號通路中與人類惡性腫瘤相關的下游主要效應靶基因,它可以活化許多不同類型的效應底物,對細胞生存、細胞周期進程,細胞生長等有很大的影響,活化的Akt在抑制細胞的凋亡、促進細胞的增殖、促進細胞的運動和侵襲方面起重要作用,與腫瘤發生發展密切相關。已經有研究發現在多種腫瘤中均有Akt及其磷酸化過度表達及活化[10]。并且在活化過程中可以誘導caspase-3抑制蛋白survivin的表達,其還可以在無ERK信號通路參與的情況下促進線粒體釋放細胞色素C,激活內源性的信號通路[11]。有研究表明當Akt過度激活后,可以促進抗凋亡基因bcl-2表達,抑制促凋亡基因bax表達,從而使細胞對化療藥物敏感性降低[12]
絲裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)是細胞內的一類絲氨酸/蘇氨酸蛋白激酶。MAPK1/2是發現最早、研究最廣的MAPK家族成員。在哺乳動物細胞中,它參與細胞周期、細胞增殖與分化、衰老和凋亡、基因轉錄等多種細胞生命事件的調控,磷酸化為其活化形式。MAPK1/2 在一系列反應如生長因子和有絲分裂等刺激中被激活,同時其持續活化最終促進細胞增殖和惡性轉化。p-ERK1/2 的下調則可促進細胞生長和生長刺激基因轉錄的抑制。目前已經發現 MAPK在一些惡性腫瘤中被明顯激活[13]。
本研究結果表明,當下調FAK表達之后,Akt及MAPK表達無明顯改變,但其磷酸化明顯被抑制,說明在肝癌細胞LM3中,FAK可以影響Akt及MAPK的活化程度,通過下調FAK表達,可以對惡性腫瘤中異常激活的信號通路產生影響,并起到抑制腫瘤的作用。同時通過細胞遷移、侵襲實驗發現,下調FAK之后腫瘤細胞遷移、侵襲能力下降。流式細胞儀檢測下調FAK后的LM3細胞主要停留在G0/G1期并且細胞凋亡增多。MTT實驗也證明下調FAK表達可使細胞增殖率下降。
以FAK基因為作用靶點,構建穩定低表達FAK基因的人肝癌細胞株LM3,并探討了RNAi對于該基因表達的抑制作用及沉默該基因表達后對相關蛋白表達和腫瘤細胞生物學特性的影響,為下一步實驗打下基礎并為肝癌的基因治療提供理論依據。
[1] Donato DM, Ryzhova LM,Meenderink LM,et al.Dynamics and mechanism of p130Cas localization to focal adhesions[J]. J Biol Chem, 2010, 285(27):20769-20779.
[2] Wendt MK, Smith JA, Schiemann WP. p130Cas is required for mammary tumor growth and transforming growth factor-β-mediated metastasis through regulation of Smad2/3 activity[J]. J Biol Chem, 2009,284(49):34145-34156.
[3] Zhao J, Zheng C, Guan J. Pyk2 and FAK differentially regulate progression of the cell cycle[J]. Cell Sci,2000,113(Pt 17):3063-3072.
[4] Yamamoto D,Sono Y,Hasegawa M,et al.FAK overexpression upregulates cyclin D3 and enhances cell proliferation via the PKC and PI3-kinase-Akt pathways[J]. Cell Signal,2003,15(6):575-583.
[5] Kurenvoa E,Xu LH,Yang X,et al.Focal adhesion kinase suppresses apoptosis by binding to the death domain of receptor-interacting protein[J]. Mol Cell Biol, 2004,24(10):4361-4371.
[6] 許呂宏,方建培,Le Yi,等.FAK基因沉默誘導白血病細胞凋亡[J].中國病理生理雜志,2010,26(7):1352-1355.
[7] Brummelkamp TR, Bernards R, Agami R. A system forstable expression of short interfering RNAs in mammalian cells[J]. Science, 2002, 296(5567): 550-553.
[9] Yuan Z, Zheng Q, Huang XY, et al. The role of FAK expression inhibition by RNA interference on liver cancer cells[J]. Zhonghua Wai Ke Za Zhi, 2007,45(19):1350-1353.
[10]Saglam O,Garrett CR,Boulware D,et al.Activation of the serine/threonine protein kinase AKT during the progression of colorectal neoplasia[J].Clin Colorectal Cancer,2007,6(9):652-656.
[11]Hu H,Jiang C,Li G,et al.PKB/AKT and ERK regulation of caspase-mediated apoptosis by methylseleninic acid in LNCaP prostate cancer cells[J].Carcinogenesis,2005,26(8):1374-1381.
[12]謝 霞,高 青,王艷麗.抑制PI3K/PKB通路逆轉胃癌耐藥的作用及其機制[J].第三軍醫大學學報,2009,31(3):227-229.
[13]趙 楊,趙 林,郭科軍,等. 細胞外信號調節激酶在乳腺癌中的表達及意義[J]. 中國醫科大學學報,2007,36(3):328-329.
ConstructionofFAK-shRNAretroviralvectorandscreeningofstableHCC-LM3celllinewithpersistentknockdownofFAK
Aliya, LIN Wu, HE Qiang
(DepartmentofHematology,TheFirstAffiliatedHospital,SunYat-senUniversity,Guangzhou510080,China.E-mail: 1heqiang@hotmail.com)
AIM: To construct a recombinant retroviral vector of short interfering RNA targeting focal adhesion kinase (FAK) gene and to establish a cell line with stable knockdown of FAK.METHODSThe oligonucleotides that transcribed to short hairpin RNA (shRNA) targeting FAK gene were synthesizedinvitro, cloned into retroviral vector pSuper.retro and transfected into Phoenix cell line. The stable clones were screened and high-titer virus was produced. The human hepatocellular carcinoma cell line HCC-LM3 was infected with the virus-rich supernatant. The stable LM3 cell line, which showed significantly to silence FAK and associated proteins, was selected by puromycin.RESULTSThe recombinant retroviral vector was successfully constructed. Persistent knockdown of FAK in the LM3 cell line infected with the supernatant containing the retrovirus was confirmed by Western blotting. Down-regulation of FAK resulted in the inhibition of p-Akt and p-MAPK1/2 expression and led to decreased migration and invasion of the cells. The cell cycle was blocked at G0/G1phase, and apoptosis was increased. The proliferation rate also decreased significantly.CONCLUSIONFAK-shRNA virus generated by recombinant retroviral vector pSuper-FAK can inhibit the protein expression of FAK and phosphorylation of Akt and MAPK1/2 in HCC-LM3 cells. Down-regulation of FAK shows a significant impact on biological behaviors of tumor cells.
Focal adhesion kinase; RNA interference; Retroviral vector; Liver neoplasms
1000-4718(2011)04-0688-07
R363
A
10.3969/j.issn.1000-4718.2011.04.013
2010-11-01
2011-02-25
廣東省自然科學基金資助項目(No.07001638)
△通訊作者 Tel:020-87755766-8214;E-mail:1heqiang@hotmail.com
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
