基于雙單倍體牙鲆挖掘與生長性狀連鎖的微衛星標記
2011-08-08 06:38:12劉永新王桂興王玉芬孫朝徽張曉彥劉海金
東北農業大學學報 2011年9期
關鍵詞:生長
劉永新,王桂興,王玉芬,司 飛,孫朝徽,張曉彥,劉海金*
(1.中國水產科學研究院,北京 100141;2.中國水產科學研究院北戴河中心實驗站,山東 秦皇島 066100)
人工誘導雌核發育作為細胞工程育種的一種新技術,是快速建立純系的有效手段,在魚類染色體操作、遺傳改良以及性別控制等方面都具有潛在的應用價值[1]。有絲分裂雌核發育和減數分裂雌核發育是魚類雌核發育兩種主要形式。有絲分裂雌核發育是經抑制第一次卵裂而形成,其染色體組中一條是以另一條為模板復制而成,遺傳組成在理論上為純合,亦稱雙單倍體[2-3]。在雄性異配性別決定系統(XX雌性;XY雄性)的魚類品種中,通過靜水壓處理經過一代即可獲得完全純合的雙單倍體,但是雙單倍體的成活率非常低。到目前為止,成功誘導有絲分裂雌核發育獲得雙單倍體的魚類僅有20余種[3]。利用有絲分裂雌核發育后代開展的研究主要包括兩部分:其一是后代純合性的檢測,只有在全部檢測位點均為純合型的個體才能稱之為雙單倍體,如牙鲆[4-6]、非洲鯰魚[7]、歐鱸[8]、斑點叉尾[9]。其二是以純合的雙單倍體為親本再行誘導減數分裂雌核發育制備克隆系,目前已報道的魚類克隆有9個品種:斑馬魚[10]、青[11]、香魚[12]、鯉魚[13]、虹鱒[14]、石川氏鮭[15]、羅非魚[16-17]、牙鲆[18]和真鯛[19]。
牙鲆(Paralichthys olivaceus)是分布于我國沿海的重要鲆鰈魚類之一,在海水養殖中占有重要地位,其年產量約為7 000 t。隨著養殖規模的日益擴大,由于累代繁殖和近親交配引起了種質的明顯退化。現在培育的商業化苗種表現出生長速度降低、疾病抵抗力差、抗逆性差等缺點,嚴重影響了養殖業者的經濟效益。因此,培育具有優良經濟性狀的新品種、新品系已然成為牙鲆養殖的重要任務。傳統上采用動物模型BLUP方法結合家系選育技術進行新品種選育研究。隨著分子標記技術的蓬勃發展,越來越多的標記技術已經融入于優質品種選育過程中。分子標記技術具有信息量大,不易受環境的影響,選擇強度大,選種效率和準確性高等特點,已經引起人們廣泛關注[20]。其中,微衛星標記具有多態性豐富、易于檢測、進化所受選擇壓力小等特點,是近年來發展迅速、應用廣泛的分子標記之一。如:凡納濱對蝦、中國對蝦與生長性狀相關的微衛星標記分析[21-22];鯉魚與抗寒性狀相關的微衛星分子標記篩選[23];牙鲆耐高溫性狀相關的微衛星標記篩選[24]。本研究以中國水產科學研究院北戴河中心實驗站2007誘導成功的雙單倍體牙鲆群體為材料[6],尋找與生長性狀緊密連鎖的微衛星標記,以期為牙鲆良種選育研究奠定理論基礎。
1 材料與方法
1.1 材料
選擇中國水產科學研究院北戴河中心實驗站牙鲆選育基礎群體中性狀優良雌性親魚,輕擠雌魚腹部獲得卵子,存儲于500 mL燒杯中。輕擠雄性真鯛腹部獲得精子,存儲于1.5 mL注射器中。真鯛精液用Ringer氏液按照1∶50的比例進行稀釋。稀釋后的精液在15W×2的紫外線燈管(Toshiba,Japan)下進行照射,照距為28~30 cm,時間為60 s。照射后的精液經顯微鏡檢查活力,確認充分活躍后,將真鯛精液與卵粒均勻混合(精卵體積比1∶10),室溫靜置2 min,加入14~16℃海水激活,并用玻璃棒輕度攪拌。將受精后1 h的受精卵轉移至靜水壓力機中,施壓650 kg·cm-2持續10 min,之后將卵移至14~16℃的海水中孵化,獲得雙單倍體實驗群體。培育群體至三年齡,記錄體重(Body weight,BW)、全長(Total length,TL)、體高(Body depth,BD)等主要生長性狀數據,選擇具有顯著差異的個體各30尾,作為快速生長組(Fast growth group,FGG)和慢速生長組(Slow growth group,SGG),每組個體生長性狀描述性統計量見表1。同時采集兩組相應個體腹面鰭條,保存于-80℃冰箱中待用。
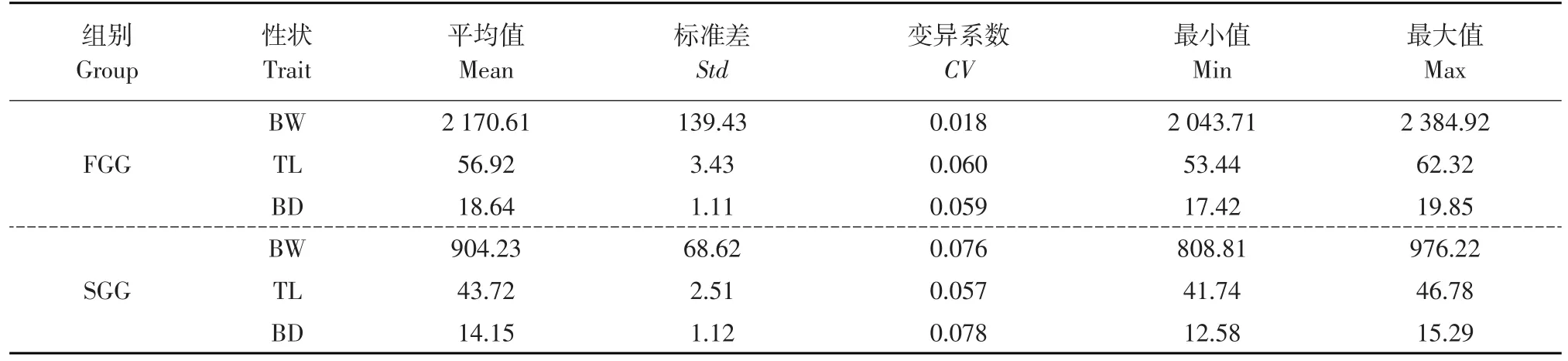
表1 FGG和SGG個體生長性狀表型值Table 1 Descriptive statistics of growth traits for FGG and SGG individuals
1.2 基因組DNA提取
將 裂 解 液(50 mmol·L-1NaCl, 30 mmol·L-1Tris-HCl, 200 mmol·L-1EDTA pH 8.0, 1% 的SDS,200 mg·L-1的蛋白酶K)加入剪碎的鰭條中,40℃消化至澄清,等體積的飽和酚∶氯仿∶異戊醇(24∶1∶1)抽提3次。等體積的去離子水稀釋,2.5倍體積預冷的無水乙醇沉淀,100 μL的70%乙醇清洗沉淀,TE溶解,-20℃保存待用。
1.3 微衛星引物
40個微衛星標記參考牙鲆遺傳連鎖圖譜獲得引物序列[25],由上海生工生物工程技術服務有限公司合成引物,合成規格2.5 OD。引物名稱、引物序列、核心序列、退火溫度、連鎖群、GenBank登錄號見表2。
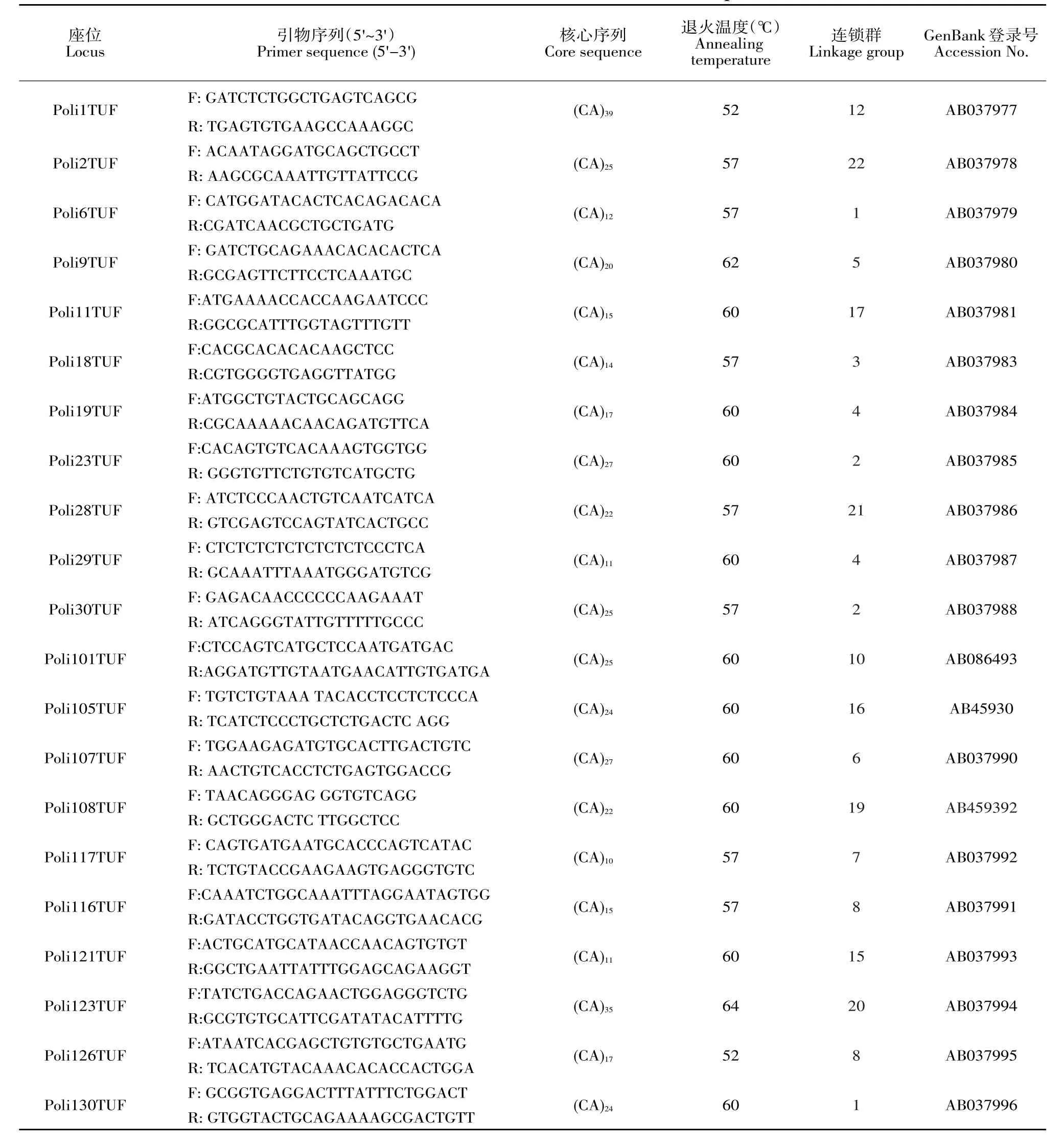
表2 牙鲆微衛星標記信息Table 2 Detailed information on microsatellite markers of Japanese flounder
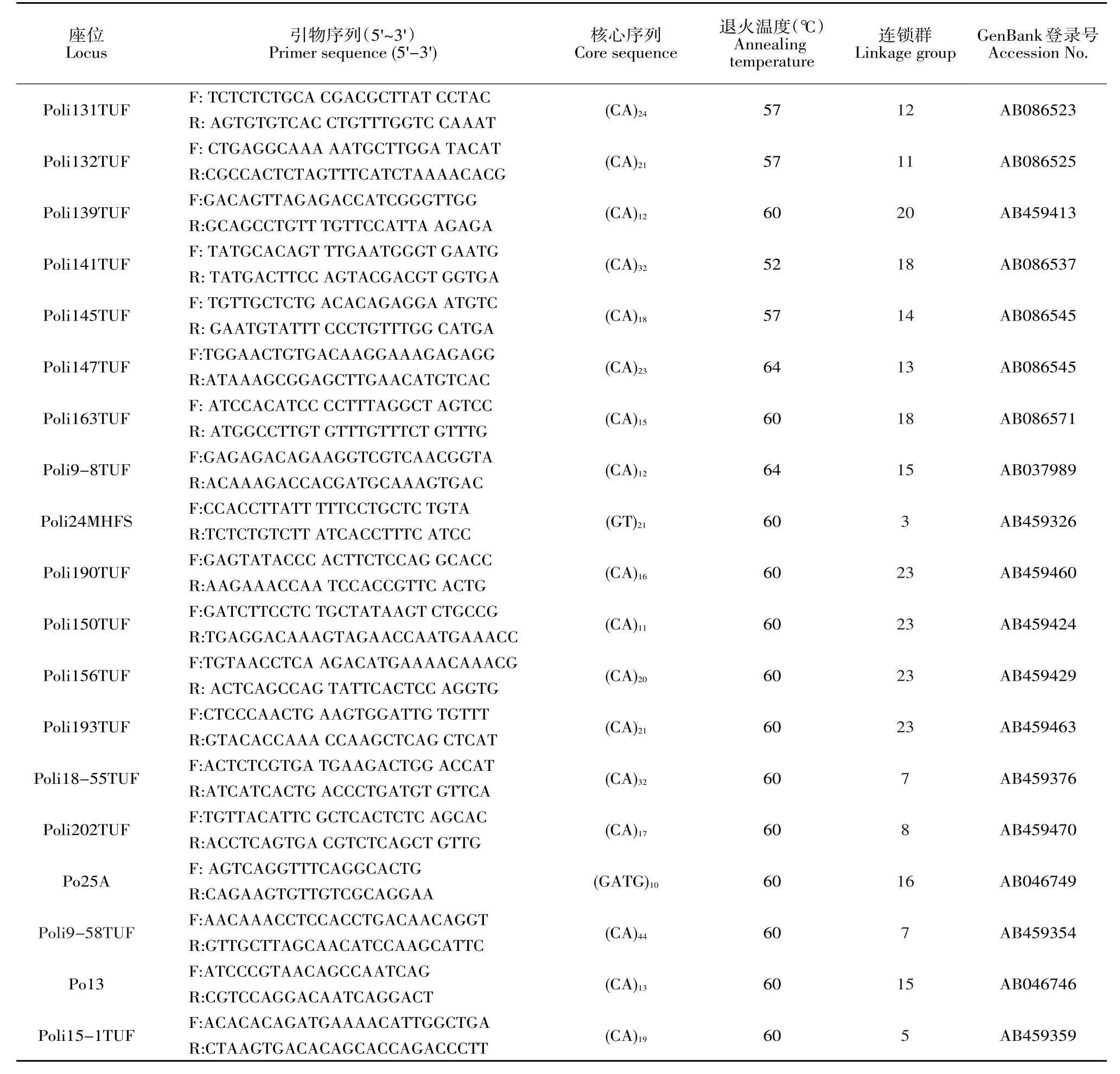
續 表
1.4 PCR反應及電泳
PCR反應體系為25 μL,包括10×的buffer 2.5 μL、Mg2+(25 mmol·L-1)1 μL、dNTPs(各2 mmol·L-1)1 mL、上下游引物(10 mmol·L-1)各1 μL、模板 1 μL(30~50 ng)、Taq DNA 聚合酶 1 U,加適量ddH2O。PCR反應程序包括:94℃30 s,退火30 s,72℃ 30 s,28循環;最后72℃延伸10 min。PCR產物經8%非變性聚丙烯酰胺凝膠電泳檢測,電泳后經1%的硝酸銀染色10 min,顯色液(1%甲醛,2%氫氧化鈉,0.04%無水碳酸鈉)顯色10 min。凝膠在掃描儀上成像,Gel-Pro Analyzer 4.5軟件對電泳譜帶進行分析。
1.5 統計分析
通過Gel-Pro Analyzer 4.5軟件分析電泳圖片計算等位基因片段大小,利用SAS8.2軟件分析生長性狀與微衛星標記的相關性。
2 結果與分析
2.1 微衛星位點差異性等位基因片段篩選
表1中個體主要生長性狀表型值范圍分別為:FGG,體重(2 043.71~2 384.92)、全長(53.44~62.32)、體高(17.42~19.85);SGG,體重(808.81~976.22)、 全 長(41.74~46.78)、 體 高(12.58~15.29),表明兩組個體生長性狀存在顯著差異。通過Gel-Pro Analyzer 4.5軟件分析電泳圖片,獲得FGG和SGG個體在不同位點的等位基因片段大小。40個微衛星標記掃描FGG和SGG個體基因組結果中,有Poli19TUF、Poli24MHFS、Poli150 TUF、Poli190TUF等4個位點在兩組內出現了差異性等位基因片段。
2.2 微衛星位點差異性等位基因片段統計
統計 Poli19TUF、Poli24MHFS、Poli150TUF、Poli190TUF位點在FGG和SGG中個體的擴增情況。表3中,同一標記在不同組別中獲得了特異性的等位基因片段。Poli19TUF、Poli24MHFS、Poli190TUF位點在FGG中均有25個左右個體擴增出相應的特異性片段,其出現率為83.33%~86.67%;而在Poli150TUF位點的出現率略低,為66.67%。SGG中Poli150TUF、Poli190TUF位點有23個左右個體擴增出相應的特異性片段,其出現率為73.33%~80.00%;而Poli24MHFS、Poli19TUF位點出現率相對較低,為40.00%~60.00%。
2.3 微衛星位點差異性等位基因片段與生長性狀的連鎖分析
利用SAS8.2軟件的GLM過程進行牙鲆主要生長性狀與微衛星標記的相關性分析,統計4個位點差異性等位基因片段在FGG和SGG內個體主要生長性狀之間的差異,尋找與性狀緊密連鎖的標記。同一標記具有特定差異性片段的兩組個體,在體重、全長、體高三個性狀上表現出顯著差異(見表4),表明Poli19TUF、Poli24MHFS、Poli 150TUF、Poli190TUF位點與生長性狀具有緊密的連鎖性。
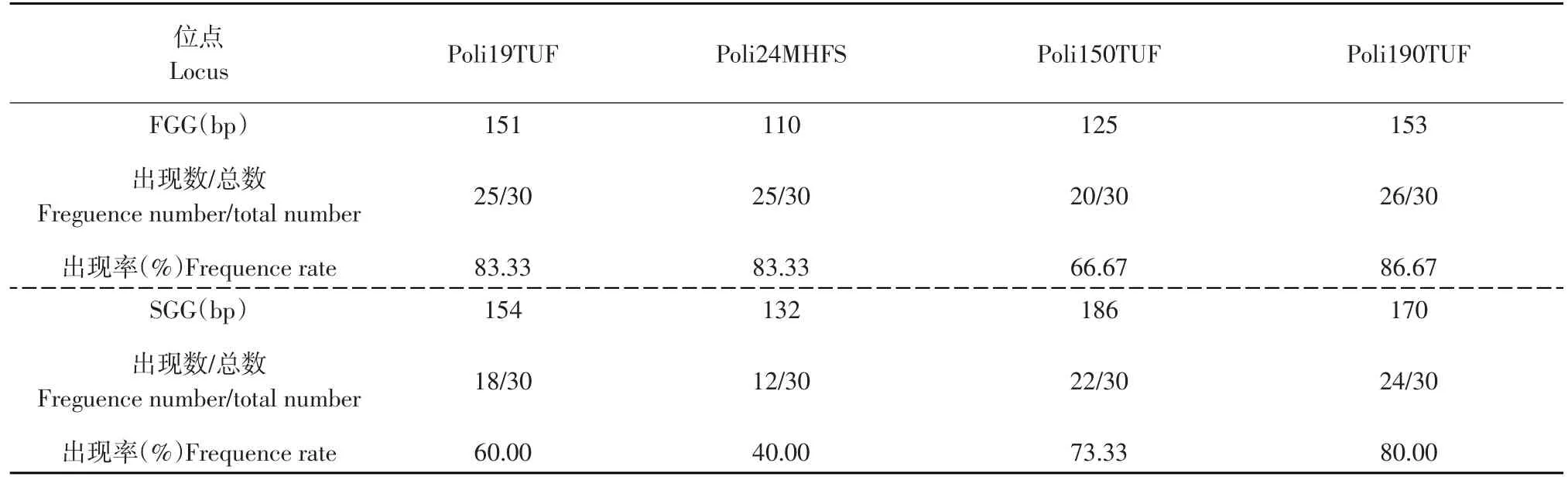
表3 FGG和SGG差異性等位基因片段統計Table 3 Statistics of different banding patterns in FGG and SGG
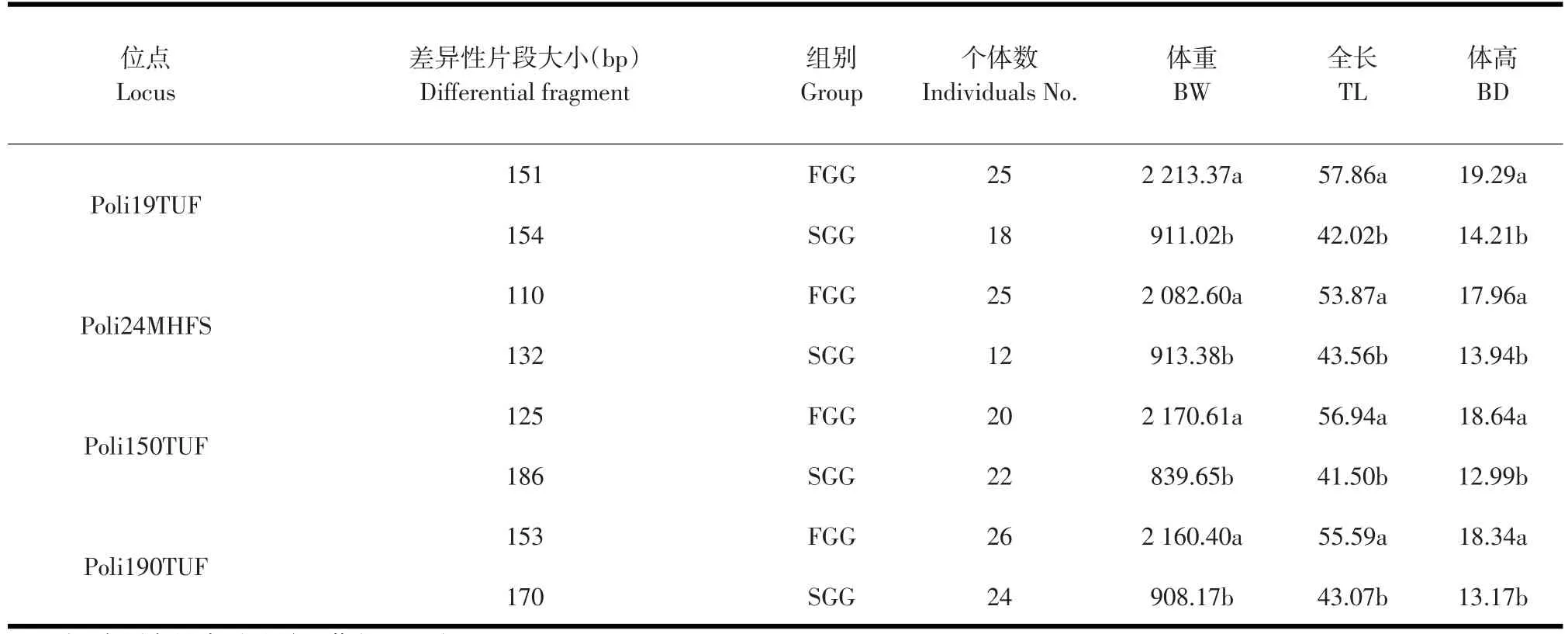
表4 FGG和SGG差異性等位基因片段個體生長性狀比較Table 4 Comparison of different banding patterns in FGG and SGG individuals
3 討論與結論
魚類的生長性狀受多基因控制,遺傳基礎復雜;且由于生存在海淡水中,較畜牧業品種更加容易受到環境效應的影響。目前,對于主要養殖魚類經濟性狀的遺傳改良多采取家系選育、人工誘導雌核發育等技術。利用這些方法,近些年先后培育出“夏奧1號”奧里亞羅非魚、吉鱺羅非魚、異育銀鯽“中科3號”、松浦鏡鯉、鰱“鄂豐1號”、加州鱸等優良新品種(系)12個。其中,雌核發育技術在水產動物育種過程中發揮了越來越重要的作用。但是,所利用的多為減數分裂雌核發育技術,以固定母本性狀和建立高遺傳相似度家系為主要手段。很少以誘導有絲分裂雌核發育獲得的雙單倍體為親本培育新品種,用此材料進行分子標記與經濟性狀的連鎖分析也未見報道。
本研究利用已經制備的雙單倍體牙鲆按照體重等主要生長性狀表型值分成FGG和SGG探索標記與生長性狀的相關性,這種分群分離分析法能夠快速鑒定與特定基因相連鎖的標記[26]。類似方法在其它魚類中也有報道,如:羅非魚[27]、鯉魚[28]、大菱鲆[29]。本研究從Coimbra等發表的遺傳連鎖圖譜上選擇了40個高多態性微衛星標記,覆蓋了牙鲆24個連鎖群中的22個,分析結果表明:4個標記與生長性狀顯著相關[25]。王桂興等初步報道了牙鲆與生長性狀相關的微衛星標記,找到了8個與生長性狀相關標記。其使用材料為隨機減數分裂雌核發育家系,在同一位點出現了多種個體基因型,且為雜合態;如Poli123TUF位點出現了6種基因型;這一點不利于標記與性狀的連鎖分析[30]。在此基礎上,本研究首次采用雙單倍體牙鲆為材料,其遺傳背景完全純合,在一個位點只有兩種基因型,便于更加準確的分析標記與性狀的相關性。同時,分析的標記數量也增加到40個;其中,3個相關標記Poli24MHFS、Poli150TUF、Poli190TUF即為新增標記。本試驗檢測出的4個相關標記分布于3個連鎖群,表明控制牙鲆生長性狀的基因有多個且位于不同連鎖群,這一點與王桂興等[30]結果相同。此外,本研究分析的3個主要生長性狀:體重、全長、體高,三者之間存在高度的表型相關和遺傳相關[31-32],若標記與其中1個性狀相關,與其余2個性狀也應出現相關性,這一推斷在本研究結果中得以輔證(4個標記與3個性狀均呈顯著相關)。當然魚類品種眾多、形態各異,處于不同的發育時期和階段使得不同生長性狀之間的相關性也可能存在不同規律。借助分子標記技術挖掘出與性狀緊密連鎖的標記,如果將個體標記效應作為隨機效應加入到動物模型等遺傳評估模型中,能夠讓性狀遺傳力和個體育種值估計更加精確,推動具有特定經濟性狀的新品種或新品系的選育進程,縮短選育周期,提高遺傳進展。
分子生物學技術的發展日新月異,從過去的DNA指紋圖譜技術、同工酶標記到微衛星、SNP標記也僅僅用了短暫的時間。伴隨著鯉魚全基因組測序的啟動,全基因組測序技術在水產動物的應用已經展開,牙鲆、半滑舌鰨、大黃魚等品種都已經完成測序,處于組裝、拼接和注釋階段。未來的品種選育與性狀遺傳改良必將借助于這些新興的技術,深入到基因組水平的層次上詮釋個體的生長發育過程。因此,在完成挖掘生長相關標記的基礎上,本研究將繼續利用牙鲆遺傳圖譜和基因組信息定位目的基因,真正發揮出分子標記技術在育種中的有效性和實用性。
[1] Thorgaard G H.Chromosome set manipulation and sex control in fish,in"Fish Physiology"(ed.by W.S.Hoar,D.J.Randall,and E.M.Donaldson)[M],Academic Press,New York,1983,9:405-434.
[2] Arai K.Genetic improvement of aquaculture finfish species by chromosome manipulation techniques in Japan[J].Aquaculture,2001,197:205-228.
[3] Komen H,Thorgaard G H.Androgenesis,gynogenesis and the production of clones in fishes:A review[J].Aquaculture,2007,269(1-4):150-173.
[4] 朱曉琛,劉海金,孫效文,等.微衛星評價牙鲆雌核發育二倍體純合性[J].動物學研究,2006,27(1):63-67.
[5] 劉海金,劉永新,侯吉倫,等.牙鲆減數分裂與有絲分裂雌核發育的遺傳差異[J].水產學報,2010,34(6):718-724.
[6] 劉海金,陸桂,王曉梅,等.有絲分裂雌核發育牙鲆的微衛星鑒定[J].中國水產科學,2010,17(5):889-894.
[7] Galbusera P,Volckaert F A M,Ollevier F.Gynogenesis in the African catfish Clarias gariepinus(Burchell,1822)Ⅲ.Inductionof endomitosis and the presence of residual genetic variation[J].Aquaculture,2000,185:25-42.
[8] Bertotto D,Cepollaro F,Libertini A,et al.Production of clonal founders in the European sea bass,Dicentrarchus labrax L,by mitotic gynogenesis[J].Aquaculture,2005,246:115-124.
[9] Geoffrey C W,Brian G B,Sylvie M A Q.Production of viable homozygous,doubled haploid channel catfish(Ictalurus punctatus)[J].Marine Biotechnology,2009,12:380-385.
[10] Streisinger G,Walker C,Dower N,et al.Production of clones of homozigous diploid zebra fish(Brachydanio rerio)[J].Nature,1981,291:293-296.
[11] Naruse K,Ijiri K,Shima A,et al.The Production of Cloned Fish in the Medaka(Oryzias ktipes)[J].Experimental Zoology,1985,236:335-341.
[12] Han H S,Taniguchi N,Tsujimura A.Production of clonal ayu by chromosome manipulation and confirmation by isozyme marker and tissue grafting[J].Nippon Suisan Gakkaishi,1991,57:825-832.
[13] Komen J,Bongers A B J,Richter C J J,et al.Gynogenesis in common carp(Cyprinus carpio L.)II.The production of homozygous gynogenetic clones and Flhybrids[J].Aquaculture,1991,92:127-142.
[14] Quillet E,Garcia P,Guyomard R.Analysis of the production of all homozygous lines of rainbow trout by gynogenesis[J].Experimental Zoology,1991,257:367-374.
[15] Kobayashi T,Ide A,Hiasa T,et al.Production of cloned amago salmon Oncorhynchus rhodurus[J].Fishery Science,1994,60:275-281.
[16] Mu¨ller-Belecke A,Ho¨rstgen-Schwark G.Sex determination in tilapia(Oreochromis niloticus)sex ratios in homozygous gynogenetic progeny and their offspring[J].Aquaculture,1995,137:57-65.
[17] Mu¨ller-Belecke A,Ho¨rstgen-Schwark G.Performance testing of clonal Oreochromis niloticus lines[J].Aquaculture,2000,184:67-76.
[18] Yamamoto E.Studies on sex-manipulation and production of cloned populations in hirame,Paralichthys olivaceus(Temminck et Schlegel)[J].Aquaculture,1999,173:235-246.
[19] Kato K,Hayashi R,Yuasa D,et al.Production of cloned red sea bream,Pagrus major,by chromosome manipulation[J].Aquaculture,2002,207:19-27.
[20] 魯紹雄,吳常信.動物遺傳標記輔助選擇研究及其應用[J].遺傳,2002,24(3):359-362.
[21] 陳曉漢,曾地剛,李詠梅,等.凡納濱對蝦生長性狀的微衛星DNA標記分析[J].武漢大學學報,2006,52(4):498-502.
[22] 張天時,劉萍,李健,等.中國對蝦與生長性狀相關微衛星DNA分子標記的初步研究[J].海洋水產研究,2006,27(5):34-38.
[23] 潘賢,梁利群,雷清泉.篩選與鯉魚抗寒性狀相關的微衛星分子標記[J].哈爾濱工業大學學報,2008(8):915-918.
[24] 盧鐘磊,池信才,王義權.褐牙鲆耐熱性狀相關的微衛星分子標記的篩選[J].廈門大學學報:自然科學版,2007(3):396-402.
[25] Coimbra M R M,Kobayashi K,Koretsugu S,et al.A genetic linkage map of the Japanese flounder,Paralichthys olivaceus[J].Aquaculture,2003,220:203-218.
[26] Michelmore R W.Identification of marker linked to diseaseresistance gene by bulked segregant analysis:a rapid method detect marker in specific genomic regions by using segregating populations[J].Proc Natl Acad Sci USA,1991,88:9828-9832.
[27] Cnani A,Halerman E M,Ron M,et al.Detection of a chromosomal region with two quantitative trait loci,affecting cold tolerance and fish size,in an F2tilapia hybrid[J].Aquaculture,2003,223:ll7-l28.
[28] 張義鳳,張研,魯翠云,等.鯉魚微衛星標記與體重、體長和體高性狀的相關性分析[J].遺傳,2008,30(5):613-615.
[29] 許可,馬愛軍,王新安,等.大菱鲆(Scophthalmus maximus)生長性狀相關的微衛星標記篩選[J].海洋與湖沼,2008,40(5):577-583.
[30] 王桂興,劉永新,孫效文,等.牙鲆微衛星分子標記與生長性狀的相關性分析[J].東北農業大學學報,2009,40(7):77-84.
[31] 顧憲明,劉永新,王凱,等.應用DF-REML法估計牙鲆性狀遺傳參數[J].東北農業大學學報,2008,39(11):71-75.
[32] 王凱,劉海金,劉永新,等.牙鲆形態性狀對體重的影響效果分析[J].上海水產大學學報,2008,17(6):655-660.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
