花生四烯酸與氧化應激的研究進展*
2011-07-31 14:05:48傅淑霞
中國病理生理雜志 2011年11期
陶 磊,傅淑霞
(河北醫科大學第二醫院腎內科,河北 石家莊 050000)
花生四烯酸(arachidonic acid,AA)是一種人體內含量最豐富、性質最活躍、分布最廣泛的多不飽和必需脂肪酸,具有很強的生物活性,其代謝網絡是炎癥代謝網絡的重要組成部分。AA不僅在炎癥中發揮重要作用而且與氧化應激關系密切。對兩者相互作用的研究能夠更好地闡明疾病發生的機制,對病因探究及疾病治療具有重要意義。
1 AA及其代謝
AA屬于n-6系多不飽和必需脂肪酸,簡記為20:4(n-6),由油酸轉化而來。在哺乳動物體內,絕大部分AA與甘油磷脂上的羥基結合,以酯的形式幾乎全部存在于膜磷脂Sn-2的位置。通過以下3條途徑從膜磷脂中釋放:(1)非特異性磷脂酶A2的直接水解;(2)磷脂酰肌醇特異性的磷脂酶C和二酰甘油脂肪酶聯合作用;(3)特異性磷脂酶A2水解作用。其次AA的釋放還受酰基轉移酶的影響,抑制其活性,可以降低再酰化的速率,提高游離 AA的水平[1]。釋放的 AA經3條途徑被代謝:(1)環氧酶(cyclooxygenase,COX)途徑,催化生成前列腺素類和血栓素;(2)脂氧酶(lipoxygenase,LOX)途徑,生成過氧化氫廿碳四烯酸、白三烯類、羥化廿碳四烯酸和脂氧素;(3)經細胞色素 P450酶(cytochrome P450,CYP450)途徑,生成羥化廿碳四烯酸、環氧化二十碳四烯酸及其下級產物二羥基二十碳三烯酸。
2 AA對氧化應激的影響
氧化應激指機體在遭受各種有害刺激時,體內高活性分子如活性氧自由基(reactive oxygen species,ROS)和活性氮自由基(reactive nitrogen species,RNS)產生過多,超出了機體對氧化物的清除能力,氧化系統和抗氧化系統失衡,導致組織損傷。組織和細胞的氧化損傷被認為是許多人類疾病發展的中心環節,正常的氧化代謝會產生活性氧,但濃度非常低,低濃度的活性氧可作為信號轉導通路中的信號分子,或者本身作為細胞的刺激物發揮重要的生理作用,然而高濃度的活性氧是致命的,氧化應激過程中產生的大量活性氧通過脂質過氧化反應,對膜磷脂內的脂肪酸進行修飾,改變了膜的流動性、膜蛋白結構及細胞信號轉導通路等,導致細胞的壞死或凋亡。
2.1 AA誘導氧化應激的產生 AA對氧化應激的誘導主要是通過產生大量活性氧來實現的。AA主要通過以下幾條通路促進活性氧的大量產生:首先,AA能夠激活細胞膜上的鈣通道致鈣內流,使細胞內鈣離子濃度增加[2,3],增加的鈣離子能夠激活 NADPH氧化酶,產生大量活性氧[4]。其次,AA和高糖共同作用能夠激活蛋白激酶C(protein kinase C,PKC)進而激活PKC依賴的NADPH氧化酶,產生大量活性氧[5]。通過使用活性氧(ROS)捕獲劑雙氫羅丹明123,進一步證實了AA和高糖能夠顯著增加ROS的形成,并且隨著AA和高糖的濃度增加,培養液中脂質過氧化物及氧化型谷胱甘肽(L-glutathione oxidized,GSSG)的量也逐漸增加,從而說明氧化應激逐漸加重[6,7]。AA還能誘導人中性粒細胞釋放超氧基[8]。此外,AA通過激活受體交互作用蛋白1(receptor- interacting protein 1,RIP1),產生大量ROS[9]。AA還能通過抑制線粒體呼吸鏈中的復合酶I和III的活性以及增加線粒體膜的流動性,誘導ROS的產生,并且有研究發現在老鼠的心肌細胞線粒體中,ROS的產生量和復合酶III的抑制程度呈線性正相關[10]。除上述途徑外,AA在通過COX、LOX途徑代謝時,也產生大量活性氧,包括脂質過氧化物和超氧陰離子,加重氧化應激[11]。
2.2 AA抑制氧化應激 AA通過提高抗氧化酶活性及減少線粒體ROS的生成,抑制氧化應激,見圖1。AA通過激活過氧化物酶增殖體激活受體(peroxisome proliferator-activated receptor,PPAR)及促進新蛋白的合成,提高了銅鋅超氧化物歧化酶(copper/zinc superoxide dismutase,Cu/Zn-SOD)、錳超氧化物歧化酶、谷胱甘肽-過氧化物酶(glutathione peroxidase,GPx)及過氧化氫酶(catalase)4種抗氧化酶的活性,對抗谷氨酸鹽及過氧化氫(H2O2)造成的氧化應激損傷,并且對于提高每種抗氧化酶活性所需AA的量及誘導時間各不相同,具有特異性[12]。PPAR包括PPAR-α、PPAR-γ等多種亞型,為進一步明確AA通過激活何種亞型而發揮作用,在實驗中使用了PPAR-α抑制劑MK886,PPAR-γ抑制劑雙酚A二縮水甘油醚(bisphenol A diglycidyl ether,BADGE)以及真核生物蛋白合成抑制劑放線菌酮(cycloheximide),發現其中BADGE和cycloheximide能夠完全阻斷AA的保護作用,MK886不能阻斷AA的保護作用,證明AA對抗氧化應激的神經保護作用主要是通過激活PPAR-γ和合成新蛋白來實現的。盡管MK886不能阻斷AA的保護作用,但是也不能除外AA可能通過PPAR-α途徑發揮保護作用的可能性,因為MK886在阻斷PPAR-α同時,其本身有直接的神經保護作用[12]。AA還能通過激活表皮生長因子受 體 (epidermal growth factor receptor,EGFR)[13],進而激活谷胱甘肽過氧化物酶[14]。除上述通過提高抗氧化酶活性抑制氧化應激外,AA還通過以下途徑減少ROS的生成抑制氧化應激:首先,AA通過解偶聯作用,降低線粒體膜電位,阻止反向電子轉運(reverse electron transport,RET),進而阻止依賴 RET的ROS產生[10]。其次,AA通過降低NADPH氧化酶p47亞基的表達,抑制NADPH氧化酶的活性[15],減少ROS的生成。AA還能通過抑制一氧化氮合酶(nitric oxide synthase,NOS)的活性,抑制具有強氧化性的ROS-過氧亞硝酸鹽(ONOO-)的形成[16]。
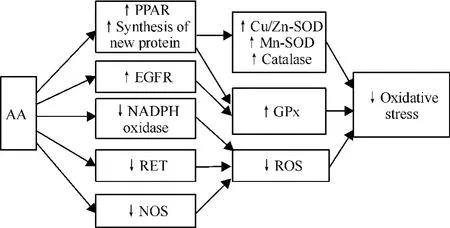
Figure 1.The inhibitory effect of AA on oxidative stress.圖1 花生四烯酸對氧化應激的抑制作用
3 氧化應激對AA的影響
3.1 氧化應激促進AA的釋放
①氧化應激通過增強磷脂酶A2活性,促進AA釋放。磷脂酶A2是一種能水解甘油磷脂分子中第2位酯鍵的酶,分布廣泛,根據其存在部位、氨基酸順序同源性及生化功能不同分為3類:分泌型磷脂酶A2(secreted phospholipase A2,sPLA2);胞質型磷脂酶A2(cytosolic phospholipase A2,cPLA2),這兩者都表現出對Ca2+的高度敏感性;以及Ca2+非依賴型磷脂酶A2(calcium - independent phospholipase A2,iPLA2)。其中cPLA2被認為是氧化應激狀態下激活產生AA的主要酶,cPLA2水平與AA的釋放量呈正相關。cPLA2主要通過以下幾條途徑激活,見圖2。(1)氧化應激能夠增加細胞內游離鈣離子濃度([Ca2+]i),增加的鈣離子,一方面通過激活絲裂原激活的蛋白激酶/細胞外信號調節激酶的激酶1(MAPK/ERK kinase 1,MEK-1),進而激活細胞外信號調節蛋白激酶1/2(extracellular signal-regulated kinase 1/2,ERK1/2),使cPLA2磷酸化激活;另一方面促進cPLA2從胞質向膜的轉運,這兩方面共同促進膜磷脂釋放AA[17]。(2)氧化應激能夠激活 PKC,被激活的PKC一方面通過激活ERK1/2,磷酸化激活cPLA2;另一方面PKC也能直接磷酸化激活cPLA2,促進AA的釋放[17]。(3)氧化應激能夠直接誘導ERK1/2和p38絲裂原激活蛋白激酶(p38 mitogen-activated protein kinase,p38 MAPK)的磷酸化激活[18],激活的這兩種酶能夠分別誘導cPLA2磷酸化,激活cPLA2,釋放 AA[19,20]。(4)氧化應激還能通過 Raf- MEK -p42/44(MAPK)通路,致 cPLA2磷酸化,激活 cPLA2,釋放AA[21]。(5)氧化應激狀態下膜磷脂發生脂質過氧化,cPLA2對氧化的膜磷脂具有高度親和力和選擇性的降解作用,進一步增強cPLA2活性,增加AA釋放[21]。(6)氧化應激下存在sPLA2的激活,激活的sPLA2通過增強PKC和ERK的活性,進而增加cPLA2的激活,促進 AA釋放[17]。除上述 cPLA2激活外,氧化的膜磷脂能夠顯著增加sPLA2的活性[22],并且氧化應激下激活的cPLA2也能進一步增強sPLA2的活性[17]。由此可見,氧化應激狀態下存在cPLA2和sPLA2兩種酶的激活,并且兩者之間存在著復雜相互關系,共同作用促進AA釋放。

Figure 2.The activation of cPLA2and the release of AA under oxidative stress.圖2 氧化應激下cPLA2的激活及AA的釋放
對于iPLA2是否參與氧化應激下AA的釋放,存在爭議,在星形膠質細胞研究中發現,氧化應激狀態下抑制 iPLA2的活性,對 AA的釋放沒有影響[18]。然而在U937吞噬細胞研究中卻發現,iPLA2是氧化應激下釋放AA的主要酶[23]。
②氧化應激抑制AA的再酰化,增加游離的AA濃度。AA參與了膜磷脂的脫酰基和再酰基作用循環,AA被PLA2從磷脂膜上水解下來,另1個脂肪酸通過酰基輔酶A合成酶和溶血磷脂酯酰轉移酶結合到膜磷脂上,這個循環是構成AA和其它脂肪酸結合到磷脂上的主要通路,氧化應激狀態時AA的再酰化被抑制。氧化應激通過損傷線粒體,抑制三磷酸腺苷(adenosine triphosphate,ATP)的合成[24],而 ATP 是合成酰化作用中間產物脂肪酰輔酶A所必須的,通過抑制ATP的合成,從而抑制了AA的酯化作用,增加了AA的釋放。
3.2 氧化應激抑制血小板 AA的釋放 Sorrell等[25]利用吩嗪衍生物綠膿菌素的氧化還原特性產生了少量的活性氧自由基,來研究氧化應激對人血小板內AA代謝的影響,血小板內的AA預先用[3H]標記,并且向培養液中加入鈣離子載體A23187,使鈣離子快速進入細胞內,保證了細胞質的游離鈣離子濃度,結果發現用綠膿菌素處理的血小板游離AA標記量減少,相對應的是膜磷脂上的標記增加。在向培養液中加入綠膿菌素前用抗氧化劑,超氧化物歧化酶,過氧化氫酶或者羥自由基清除劑,甘露醇等預處理,能夠恢復AA的釋放和類花生四烯酸的量,從而說明氧化應激抑制血小板膜磷脂AA釋放,然而,綠膿菌素產生的活性氧對加入外源性AA的代謝沒有影響。
4 小結
綜上所述,AA既能誘導氧化應激產生,也能抑制氧化應激,同時氧化應激對AA的釋放也有正反兩方面作用。AA作為一種重要的炎癥因子,與氧化應激之間存在著復雜的相互關系,不同細胞間二者的相互關系也不盡相同,通過AA的橋梁作用,不僅能夠將炎癥和氧化應激有機結合起來,從炎癥和氧化應激兩方面系統闡述疾病發生發展全過程,而且還能從這兩個方面對疾病進行治療,對臨床疾病的診治具有重大意義。
[1]陳 瑗,周 玫.自由基醫學基礎與病理生理[M].第1版.北京:人民衛生出版社,2002.175 -179.
[2]Fiorio Pla A,Genova T,Pupo E,et al.Multiple roles of protein kinase A in arachidonic acid-mediated Ca2+entry and tumor- derived human endothelial cell migration[J].Mol Cancer Res,2010,8(11):1466 -1476.
[3]Thompson J,Mignen O,Shuttleworth TJ.The N - terminal domain of Orai3 determines selectivity for activation of the store - independent ARC channel by arachidonic acid[J].Channels(Austin),2010,4(5):398 -410.
[4]Jagnandan D,Church JE,Banfi B,et al.Novel mechanism of activation of NADPH oxidase 5.Calcium sensitization via phosphorylation[J].J Biol Chem,2007,282(9):6494-6507.
[5]Inoguchi T,Li P,Umeda F,et al.High glucose level and free fatty acid stimulate reactive oxygen species production through protein kinase C-dependent activation of NAD(P)H oxidase in cultured vascular cells[J].Diabetes,2000,49(11):1939 -1945.
[6]Lee SJ,Mun GI,An SM,et al.Evidence for the association of peroxidases with the antioxidant effect of p-coumaric acid in endothelial cells exposed to high glucose plus arachidonic acid[J].BMP Rep,2009,42(9):561 -567.
[7]Mun GI,An SM,Park H,et al.Laminar shear stress inhibits lipid peroxidation induced by high glucose plus arachidonic acid in endothelial cells[J].Am J Physiol Heart Circ Physiol,2008,295(5):H1966 - H1973.
[8]黃鐵生.花生四烯酸及其衍生物對腫瘤壞死因子誘導的中性粒細胞超氧基釋放的影響[J].中國病理生理雜志,2000,16(4):347 -350.
[9]Kim S,Dayani L,Rosenberg PA,et al.RIP1 kinase mediates arachidonic acid-induced oxidative death of oligodendrocyte precursors[J].Int J Physiol Pathophysiol Pharmacol,2010,2(2):137 -147.
[10]Sch?nfeld P,Wojtczak L.Fatty acids decrease mitochondrial generation of reactive oxygen species at the reverse electron transport but increase it at the forward transport[J].Biochim Biophys Acta,2007,1767(8):1032 -1040.
[11]陳 瑗,周 玫.自由基-炎癥與衰老性疾病[M].第1 版.北京:科學出版社,2007.11 -16.
[12]Wang ZJ,Liang CL,Li GM,et al.Neuroprotective effects of arachidonic acid against oxidative stress on rat hippocampal slices[J].Chem Biol Interact,2006,163(3):207-217.
[13]Freeman EJ,Sheakley ML,Clements RJ.Angiotensin II-dependent growth of vascular smooth muscle cells requires transactivation of the epidermal growth factor receptor via a cytosolic phospholipase A(2)-mediated release of arachidonic acid[J].Arch Biochem Biophys,2010,498(1):50-56.
[14]Duval C,Augé N,Frisach MF,et al.Mitochondrial oxidative stress is modulated by oleic acid via an epidermal growth factor receptor-dependent activation of glutathione peroxidase[J].Biochem J,2002,367(Pt3):889 -894.
[15]Keane DC,Takahashi HK,Dhayal S,et al.Arachidonic acid actions on functional integrity and attenuation of the negative effects of palmitic acid in a clonal pancreatic βcell line[J].Clin Sci(Lond),2011,120(5):195 -206.
[16]Palomba L,Cerioni L,Cantoni O.Arachidonic acid inhibits neuronal nitric oxide synthase elicited by proinflammatory stimuli and promotes astrocyte survival with both exogenous and endogenous peroxynitrite via different mechanisms[J].J Neurosci Res,2010,88(11):2459 -2468.
[17]Han WK,Sapirstein A,Hung CC,et al.Cross-talk between cytosolic phospholipase A2α(cPLA2α)and secretory phospholipase A2(sPLA2)in hydrogen peroxide-induced arachidonic acid release in murine mesangial cells:sPLA2regulates cPLA2alpha activity that is responsible for arachidonic acid release[J].J Biol Chem,2003,278(26):24153-24163.
[18]Zhu D,Hu C,Sheng W,et al.NAD(P)H oxidase-mediated reactive oxygen species production alters astrocyte membrane molecular order via phospholipase A2[J].Biochem J,2009,421(2):201 -210.
[19]Shelat PB,Chalimoniuk M,Wang JH,et al.Amyloid beta peptide and NMDA induce ROS from NADPH oxidase and AA release from cytosolic phospholipase A2in cortical neurons[J].J Neurochem,2008,106(1):45 -55.
[20]Nito C,Kamada H,Endo H,et al.Role of the p38 mitogen-activated protein kinase/cytosolic phospholipase A2signaling pathway in blood-brain barrier disruption after focal cerebral ischemia and reperfusion[J].J Cereb Blood Flow Metab,2008,28(10):1686 -1696.
[21]van Rossum GS,Drummen GP,Verkleij AJ,et al.Activation of cytosolic phospholipase A2in Her14 fibroblasts by hydrogen peroxide:a p42/44MAPK-dependent and phosphorylation- independent mechanism[J].Biochim Biophys Acta,2004,1636(2 -3):183 -195.
[22]Korotaeva AA,Samoilova EV,Piksina GF,et al.Oxidized phosphatidylcholine stimulates activity of secretory phospholipase A2group IIA and abolishes sphingomyelininduced inhibition of the enzyme[J].Prostaglandins Other Lipid Mediat,2010,91(1 -2):38 -41.
[23]Pérez R,Balboa MA,Balsinde J.Involvement of group VIA calcium-independent phospholipase A2in macrophage engulfment of hydrogen peroxide-treated U937 cells[J].J Immunol,2006,176(4):2555 - 2561.
[24]Maenpaa CJ,Shames BD,Van Why SK,et al.Oxidant-mediated apoptosis in proximal tubular epithelial cells following ATP depletion and recovery[J].Free Radic Biol Med,2008,44(4):518 -526.
[25]Muller M,Sorrell TC.Oxidative stress and the mobilisation of arachidonic acid in stimulated human platelets:role of hydroxyl radical[J].Prostaglandins,1997,54(1):493-509.
猜你喜歡
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛生(2016年3期)2016-11-12 13:23:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
西南軍醫(2015年2期)2015-01-22 09:09:37
中國衛生(2014年12期)2014-11-12 13:12:52
