常見遺傳性聾致病基因研究進展及基因診斷的臨床應用
2011-06-05 14:36:25周學軍歐陽小梅劉學忠
聽力學及言語疾病雜志 2011年1期
關鍵詞:基因突變
周學軍 歐陽小梅 劉學忠
·綜述·
常見遺傳性聾致病基因研究進展及基因診斷的臨床應用
周學軍1,2歐陽小梅1劉學忠1
耳聾的原因分為遺傳因素和環境因素,也可以是遺傳和環境因素共同作用的結果。遺傳性聾中30%的患者同時合并有外耳畸形或其他器官系統疾病,稱為綜合征型聾(syndromic hearing impairment SHI),其余70%的患者為非綜合征型聾(non-syndromic hearing impairment,NSHI)。根據遺傳模式,遺傳性聾又分為常染色體顯性(DFNB)、隱性(DFNA)、性連鎖和線粒體母系遺傳性聾。在NSHI中,DFNB占80%,DFNA約占15%~20%,性連鎖和線粒體遺傳性聾約占1%[1]。迄今為止,已發現400多個伴有聽覺障礙的綜合征,鑒定出與之相關的遺傳缺陷30多個,而在NSHI中,已定位了100多個致病位點(http://webh01.ua.ac.be/hhh/),克隆了70多個耳聾基因。近年來,遺傳性聾致病的分子生物學研究發展迅速,有關耳聾基因及其功能方面的研究成果令人矚目,本文就常見遺傳性耳聾基因的結構功能、表型特點、人群分布的研究現狀進行綜述。
1 綜合征型聾
綜合征型聾在語前聾患者中占30%,但極少出現語后聾,其正確診斷對于患者及其家族成員的伴發器官或系統(如腎臟、眼部等)病變監控具有重要的意義。臨床上有些綜合征容易診斷,有些則需要對可疑個體進行系統的檢查才能獲得診斷,由于腎臟和眼部是綜合征型聾最常伴發病變的器官,而且臨床表現較為隱蔽,因而需要進行詳細的專科檢查。常見綜合征型聾的突變基因及臨床特點見表1。
2 非綜合征型聾
GJB2基因突變所致非綜合征型遺傳性聾為雙側對稱性語前聾,主要的聽力曲線圖為殘余型、斜坡型和平坦型,極少數為U型,但沒有以低頻下降為主的上升型曲線。聽力損傷程度變異較大,可從輕度到極重度,例如35delG純合子突變患者,大多數出現極重度聾,但仍有部分患者為中度聾,極少數為輕度聾[6]。基因型和表型的關系研究表明,GJB2突變中,出現2個缺失突變的患者的耳聾程度較缺失/錯義雜合突變的患者重,而2個錯義突變的患者耳聾程度最輕[6,7]。據推測,GJB2基因突變的這些表型變化,很可能與修飾基因的作用有關,但迄今尚未得到證實[8]。
由于GJB2基因突變在遺傳性聾中的高發生率,不同種群中均存在某一優勢突變,而且其基因短小,只有2個外顯子,編碼226個氨基酸,因此,GJB2基因突變篩查已經成為耳聾分子檢測的最基本項目。35del G突變篩查在歐美國家、167del T突變篩查在猶太人群以及235delC突變篩查在東亞國家都已經獲得了很好的開展。Liu等[9]對118名中國非綜合征型聾先證者檢測GJB2基因,發現235delC突變的比例最高。Dai等[10]對中國3 004例NSHI患者進行GJB2基因突變篩查,鑒定出488例(16.3%)至少攜帶一個235delC突變位點,其中純合突變233例(7.8%),雜合突變255例(8.5%)。
2.2 SLC26A4基因-DFNB4 SLC26A4基因又名PDS基因,含有21個外顯子,編碼跨膜蛋白pendrin。Pendrin功能主要與碘/氯離子的轉運有關,并在甲狀腺、內耳和腎臟中高表達。SLC26 A4基因突變可導致Pendred綜合征(PS)和非綜合征型聾(DFNB4),均可導致顳骨的發育畸形,包括大前庭水管綜合征(large vestibular aqueduct syndrome,LVAS)和Mondini畸形。研究表明,Pendred綜合征患者完全喪失碘/氯轉運功能,而DFNB4患者碘/氯轉運功能仍存在,但處于較低水平。
迄今為止,在感音神經性聾患者中已發現150多種SLC26A4突變類型(http://www.healthcare.uiowa.edu/labs/pendredandbor/slc Mutations.htm),其中絕大部分是錯義突變,另外還有框移突變和剪接點突變。不同人種中突變的類型和發生頻率有很大差異,但大部分的病例都為散發。Campbell等[11]認為,在白種人中,最常見的SLC26A4突變位點是L236P,其次為T416P和IVS8+1G>A;在西班牙裔中,最常見突變為Q514K[12];而在亞洲,H723R在韓國和日本為頻發突變[13~15],在中國,由LVAS或LVAS伴Mondini畸形導致的聽力障礙中最常見的SLC26 A4基因突變是IVS7-2 A>G[16~18]。有研究顯示IVS7-2A>G突變為始祖效應[19]。
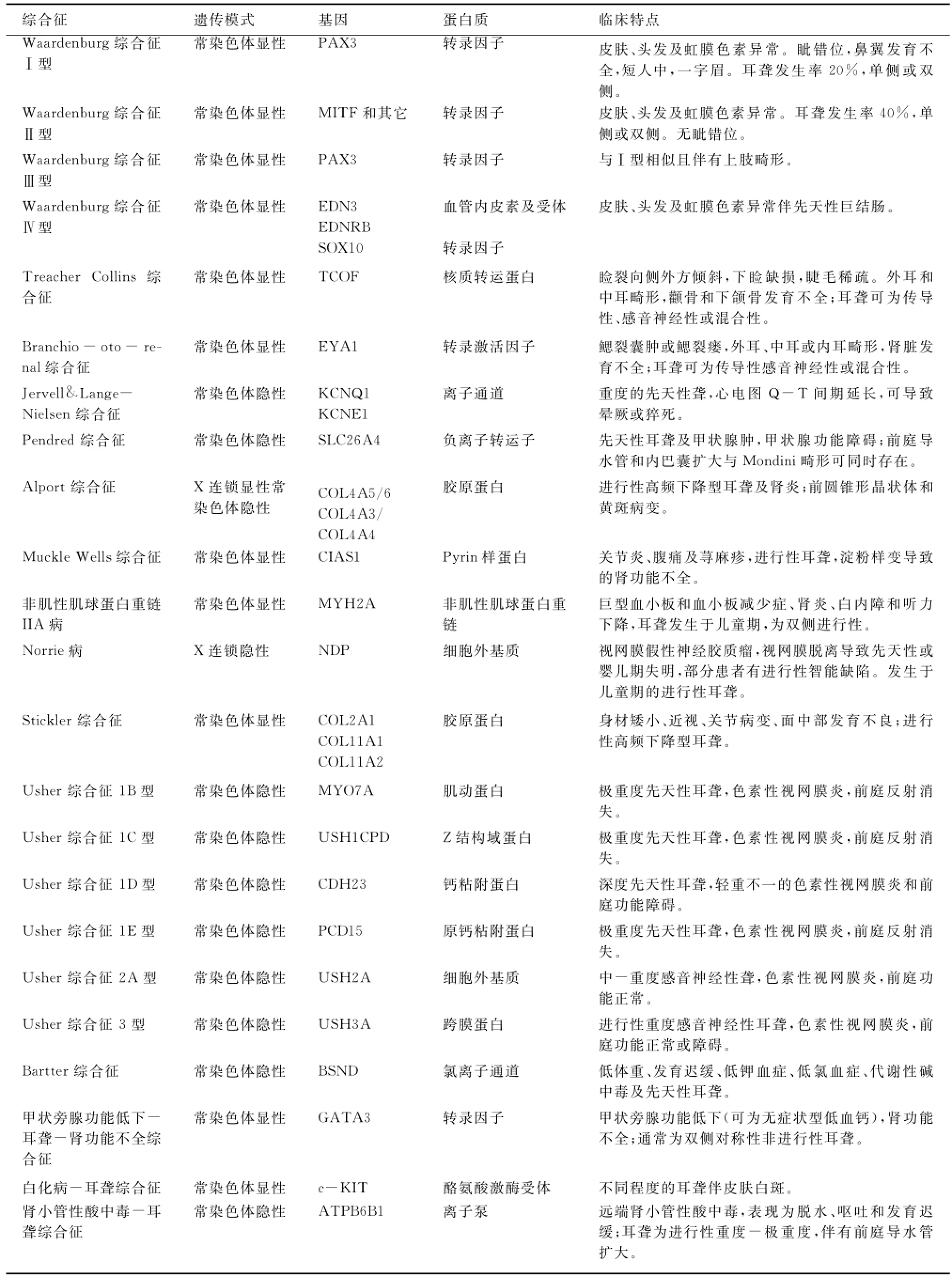
表1 常見綜合征型聾
2.3 GJB6基因-DFNB1,DFNA3 GJB6基因位于染色體13q12上,編碼連接蛋白CX30,與GJB2基因相鄰。CX30與CX26在耳蝸內同一部位表達,并且兩者編碼的氨基酸序列約77%相同。其刪除子從GJB6的5'prime末端延伸到GJB2基因,因此有可能GJB6的刪除子也刪除GJB2的區域。GJB6與GJB2共同構成了導致常染色體隱性感音神經性聾的突變位點DFNB1[20]。GJB2或GJB6的雙等位基因突變或GJB2與GJB6共存的雜合突變均可導致NSHI。GJB6最常見的突變是342kb片段缺失,但隨種群不同變異很大,發生率最高的是西班牙、法國、以色列和英國,占所有DFNB1等位基因突變的5.9%~9.7%,在所有GJB2單等位基因突變患者中,342kb片段缺失發生率達50%[21]。單倍型分析顯示此突變在德系猶太人和西歐的一些國家中有明顯的始祖效應存在[22]。另外2個GJB6突變位點是232kb和309kb片段缺失,不同種群中的發病率差異很大,中國目前尚無342kb、232kb和309kb片段缺失病例報道。
單個導致非綜合征型顯性遺傳性聾的GJB6突變目前僅在少數患者報道。Grifa等通過對198名耳聾患者進行GJB6基因突變篩查,在一個顯性遺傳的意大利家系中發現T5M錯義突變。與其他幾個Connexin蛋白一樣,GJB6基因的錯義突變,還可以導致遺傳性皮膚綜合征(clouston syndrome)。
2.4 MYO7A基因-DFNB2,DFNA11 MYO7A基因定位于染色體11q13.5,有48個外顯子,編碼肌球蛋白myosinⅦA。myosinⅦA屬于非常規肌球蛋白,在人胎兒內耳的內外毛細胞、前庭半規管的Ⅰ、Ⅱ型毛細胞均有表達,其功能目前尚不清楚,但認為其與細胞內膜的交通調控有關。MYO7A基因突變可導致DFNB2和DFNA11以及USH1B。目前已報告的突變位點在DFNB2有4個,DFNA11有5個,分別為在日本家系中發現的p.del A886-K887-K888、在美國家系中發現的p.G722R[23]、在荷蘭家系中發現的p.N458I[24]、在德國家系中發現的p.R853C[25]和在意大利家系中發現的p.A230V[26];USH1B有126個(http://www.hgmd.cf.ac.uk/ac/gene.php?gene=MYO7A)突變分布于整個MYO7A基因。
北朝中后期,羈旅生涯、流亡生活、婚姻愛情等題材在表達方式上已經發生了潛在的變化,由直爽轉變為委婉。如《紫翎馬歌辭》:“高高山頭樹,風吹葉落去,一去數千里,何當還故處?”詩歌把動亂時期背井離鄉的人,比作被山風吹落飄向遠方的樹葉,找不到自己的故里,委婉含蓄,給人很貼切的感覺。又如北魏胡太后所作《楊白花歌》:
DFNB2臨床表現為非特異性的語前或語后聾,而DFNA11的表型特點為進行性語后聾,無或僅有輕微的前庭功能障礙,聽力損失的程度、聽力曲線圖較為多變,與突變的位點有關,p.A230V、p.R853C和p.del A886-K887-K888的聽力曲線為平坦型或高頻下降型,p.N458I和p.G722R則為上升型曲線[23~26]。
2.5 GJB3基因-DFNA2 GJB3基因定位于人類染色體1p33-p35,編碼有270個氨基酸的縫隙連接蛋白Connexin31。最早在中國的2個DFNA小家系中發現該基因的錯義突變和無義突變,從而定位并成功克隆了GJB3基因。同時,動物實驗證明GJB3基因在大鼠耳蝸中有表達。Liu等在25個中國DFNB家系中檢測GJB3,發現其中2個家系成員攜帶GJB3復合雜合突變,其中之一為423-425del ATT的3bp缺失,另一為423A>G突變,其結果證明了GJB3突變不僅能導致常染色體顯性非綜合征型聾,也能導致常染色體隱性非綜合征型聾。DFNA2臨床多表現為高頻聽力受損的進行性語后聾。
2.6 WFS1基因-DFNA6/14/38 WFS1基因定位于4p16.1,編碼跨膜結構蛋白wolframin,主要分布在內質網,可能參與蛋白質轉運及調節鈣離子的動態平衡,其在外周聽覺系統的表達目前尚未清楚。WFS1基因突變可以引起常染色體顯性感音神經性聾(DFNA6/14/38)和常染色體隱性Wolframin綜合征。目前已經至少發現了該基因的30多種導致DFNA6/14/38的錯義突變[27,28]和200多種導致Wolframin綜合征的突變(http://www.khri.med.umich.edu/research/lesperance_lab/low_freq.php)。研究顯示當WFS1基因發生失活突變時導致wolframin綜合征,而在編碼C-末端第8外顯子的非失活性雜合錯義突變則導致低頻下降為主的感音神經性聾[29]。
DFNA6/14/38患者臨床表現為發展緩慢、雙側對稱的低頻聽力下降[30,31],此不同于DIAPHI基因突變引起的DFNA1。言語辨別率良好,如果僅在2 k Hz及以下頻率受累時,常不自覺耳聾的存在,隨著年齡增大,高頻區聽力也逐漸受累,聽力曲線變為平坦型。Wolframin綜合征表現為典型的發展迅速的高頻聽力下降,在人群中的雜合性攜帶率為0.3%~1%。因此,對于有陽性家族史的低頻感音性聾患者進行WFS1基因的篩查是有價值的。
2.7 COCH基因-DFNA9 COCH基因定位于14q12-q13,它編碼cochlin蛋白,在耳蝸基蝸螺旋緣和螺旋韌帶的纖維細胞以及前庭迷路內的半規管壺腹脊感覺上皮的間質細胞內高表達。目前在10多個DFNA9家系中共發現了12種COCH基因突變-V66G、G88E、W117R、P51S、I109N、A119T、I109T、C542F、G87W、M512T、C542Y和V104del[32]。
DFNA9是目前發現的伴有前庭功能障礙的常染色體顯性非綜合征型聾之一,其表型在不同人群中較一致,大部分家系表現為40~60歲之間的進行性感音神經性聾[33],個別家系耳聾進展于20~40歲之間。初期為高頻聽力受損,繼之所有頻率受累,直至60~80歲發展為重度-極重度聾,部分患者出現類似于梅尼埃病的前庭功能障礙。DFNA9耳聾的外顯率幾乎為100%,而在某些家系,前庭功能障礙也表現為完全的外顯率[33],一般伴有前庭功能障礙的患者聽力下降比較嚴重。
2.8 母系遺傳線粒體基因突變 線粒體DNA(mt DNA)是除核DNA外唯一存在于細胞內的遺傳物質。共有37個編碼基因,編碼13種m RNAs、2種r RNA和22種t RNA。在有性生殖中,只有卵子才能將線粒體遺傳給受精卵,因此,線粒體基因突變只能遺傳自母系。
線粒體基因突變性耳聾在不同人群中發病率不同,有相當一部分的語后聾患者源自于線粒體基因突變[34],但很少引起語前聾。線粒體基因突變性耳聾分為綜合征型和非綜合征型耳聾,導致的綜合征有MERRF綜合征、MELAS綜合征、Pearson綜合征、Kearns-Sayre綜合征和母系遺傳性糖尿病-耳聾綜合征。對耳蝸的影響為外毛細胞功能喪失,而耳蝸移植治療有效表明耳蝸神經未受影響。
多個線粒體基因突變可導致非綜合征型聾,其中最常見的是12Sr RNA基因上1555A>G的突變。此突變最早報告于一個大的阿拉伯-以色列家族,大部分受累患者表現為嬰兒期發病的重度-極重度聾,部分為成年發病,而個別成員聽力正常,繼而發現該家系耳聾的表型可能與8號染色體上的一個未知的顯性基因有關。目前,大量研究表明1555A>G突變與氨基糖苷類抗生素(AmAn)所致的藥物性聾有關[34]。據分析,當線粒體DNA存在1555 A>G突變時,在AmAn存在的條件下,線粒體ATP產生不足,引起細胞內外Na+、K+、Ca+等離子濃度失衡,導致耳蝸和前庭細胞損傷或死亡。攜帶1555A>G突變的個體對AmAn特別敏感,即使小劑量使用也可能致聾,由于不同修飾基因的作用,1555 A>G突變的表型變化也非常大[35]。1555A>G突變率在西班牙人中為27%。在日本,無氨基糖苷類藥物接觸史的極重度耳聾個體中1555A>G突變攜帶率為1%。其他導致非綜合征型聾的線粒體基因突變包括t RNAser(UCN)上的7445A>G、7472insC、7510T>C和7511T>C突變。此外,12Sr RNA的961位點突變和1095T>C突變可能與藥物性聾和非綜合征型聾有關[36~38]。在中國,由于Am An的大量應用,線粒體基因突變性耳聾發病率顯著不同,Li等[36]對中國散發的氨基糖苷類抗生素致聾和非綜合征型聾的兒童進行了線粒體12SrRNA突變的系統分析,發現在這一人群中48%為氨基糖苷類藥物性聾,1555A>G突變占氨基糖苷類藥物性聾和非綜合征型聾的13%和2.9%,961位點突變分別占1.7%和4.4%。
3 常見遺傳性聾致病基因的診斷意義及臨床應用
目前,在大部分發達國家,新生兒聽力篩查已常規化,經過聽力篩查發現的重度-極重度聾兒童,即可進行耳聾基因篩查。眾多非綜合征型聾基因中,GJB2、SLC26A4突變在不同耳聾人群中均占有極高的比率,個別基因突變又與患者某些臨床特征相關聯,這些基因又存在著外顯子不大、有突變熱點或區域的特點,適合采用測序、限制性酶切、斑點雜交或基因掃描技術進行篩查,這使得臨床上采用正確的基因篩查策略,在短期內完成耳聾遺傳學診斷成為可行。
3.1 在中國,GJB2、SLC26A4和線粒體基因突變導致的耳聾占遺傳性聾的30%~50%[39,40],因此,新生兒出生后即針對頻發的GJB2、SLC26A4、線粒體1555A>G突變進行篩查,可以快速簡便的早期發現相當一部分遺傳性聾患兒。
3.2 耳聾如與氨基糖苷類抗生素使用相關,或者有明確的母系遺傳特征,須進行線粒體基因突變篩查,其中12SrRNA基因上的1555A>G突變為篩查重點。對攜帶mt DNA 1555A>G突變人群明確告知終生禁用氨基糖苷類藥物,可有效避免藥物性聾的發生。
3.3 SLC26A4基因突變發病率在ARNSHI中僅次于GJB2突變,臨床上表現為進行性的高頻下降型耳聾或同時伴有甲狀腺腫、前庭導水管擴大或Mondini畸形的患者,須重點篩查SLC26A4基因,篩查位點在歐美國家主要為L236P、T416P、IVS8+1G>A,在亞洲則主要為IVS7-2A>G和H723R。在大前庭水管綜合征患者,鑒定出SLC26A4基因突變后,相當一部分尚無聽力障礙或仍具有較好的殘余聽力,對此類患者早期診斷,可以為其制訂一整套的生活指導方案,預防耳聾的發生或進一步加重。由于SLC26A4基因突變篩查能夠先于CT檢查之前發現和確診LVAS,而且檢出率大于80%,在某些地區更容易進行批量檢測,因而已經成為顳骨CT篩查的重要補充方法。
3.4 臨床表現為發展緩慢的、雙側對稱的2 k Hz以下低頻聽力下降,須考慮篩查WFS1基因和MYO7A基因。
3.5 臨床表現為遲發的進行性聽力下降伴進行性前庭功能障礙患者,提示重點篩查COCH基因。
3.6 聽力篩查中出現ABR反應陰性而OAE反應陽性的兒童,提示篩查OTOF基因。
3.7 耳聾遺傳模式為X-連鎖且伴有Mondini畸形等骨迷路的發育不良患者,應考慮篩查POU3F4基因。如X-連鎖中-重度進行性感音神經性聾,應考慮篩查PRPS1基因[41]。
耳聾基因診斷明確后,根據先證者及其父母親攜帶基因類型,結合遺傳規律,即可提供準確的遺傳咨詢,評估再生育聾兒的風險進行耳聾產前診斷和耳聾基因遠程診斷。
1 Cryns K,Van Camp G.Deafness genes and their diagnostic applications[J].Audiol Neurootol,2004,9:2.
2 Kenneson A,Van Naarden Braun K,Boyle C.GJB2(connexin 26)variants and nonsyndromic Sensorineural hearing loss:A Huge review[J].Genet Med,2002,4:258.
3 Van Laer L,Coucke P,Mueller RF,et al.A common founder for the 35delG GJB2 gene mutation in connexin 26 hearing impairment[J].J Med Genet,2001,38:515.
4 Ohtsuka A,Yuge I,Kimura S,et al.GJB2 deafness gene shows a specific spectrum of mutations in Japan,including a frequent founder mutation[J].Hum Genet,2003,112:329.
5 Yan D,Park HJ,Ouyang XM,et al.Evidence of a founder effect for the 235delC mutation of GJB2(connexin 26)in east Asians[J].Hum Genet,2003,114:44.
6 Snoeckx RL,Huygen PLM,Feldmann D,et al.GJB2 mutations and degree of hearing loss:A multicenter study[J].Am J Hum Genet,2005,77:945.
7 Cryns K,Orzan E,Murgia A,et al.A genotype-phenotype correlation for GJB2(connexin 26)deafness[J].J Med Genet,2004,41:147.
8 Hilgert N,Huentelman MJ,Thorburn AQ,et al.Phenotypic variability of patients homozygous for the GJB2 mutation 35delG cannot be explained by the influence of one major modifier gene[J].Eur J Hum Genet,2009,17:517.
9 Liu XZ,Xia XJ,Ke XM,et al.The prevalence of connexin 26(GJB2)mutations in the Chinese population[J].Hum Genet,2002,111:394.
10 Dai P,Yu F,Han B,et al.The prevalence of the 235delC GJB2 Mutation in a Chinese deaf population[J].Genet Med,2007,9:283.
11 Campbell C,Cucci RA,Prasad S,et al.Pendred syndrome,DFNB4,and PDS/SLC26A4 identification of eight novel mutations and possible genotype-phenotype correlations[J].Hum Mutat,2001,17:403.
12 Pera A,Dossena S,Rodighiero S,et al.Functional assessment of allelic variants in the SLC26A4 gene involved in Pendred syndrome and nonsyndromic EVA[J].Procn Natl Acad Sci USA,2008,105:18 608.
13 Park HJ,Shaukat S,Liu XZ,et al.Origins and frequencies of SLC26A4(PDS)mutations in east and south Asians:Global implications for the epidemiology of deafness[J].J Med Genet,2003,40:242.
14 Park HJ,Lee SJ,Jin HS,et al.Genetic basis of hearing loss associated with enlarged vestibular aqueducts in Koreans[J].Clin Genet,2005,67:160.
15 Tsukamoto K,Suzuki H,Harada D,et al.Distribution and frequencies of PDS(SLC26A4)mutations in Pendred syndrome and nonsyndromic hearing loss associated with enlarged vestibular aqueduct:A unique spectrum of mutations in Japanese[J].Eur J Hum Genet,2003,11:916.
16 Reyes S,Wang G,Ouyang X,et al.Mutation analysis of SLC26A4 in mainland Chinese patients with enlarged vestibular aqueduct[J].Otolaryngol Head Neck Surg,2009,141:502.
17 Wang QJ,Zhao YL,Rao SQ,et al.A distinct spectrum of SLC26A4 mutations in patients with enlarged vestibular aqueduct in China[J].Clin.Genet,2007,72:245.
18 Dai P,Li Q,Huang D,et al.SLC26A4 c.919-2A4G varies among Chinese ethnic groups as a cause of hearing loss[J].Genet Med,2008,10:586.
19 Yang JJ,Tsai CC,Hsu H M,et al.Hearing loss associated with enlarged vestibular aqueduct and Mondini dysplasia is caused by splice-site mutationin the PDS gene[J].Hear Res,2005,199:22.
20 Common JE,Bitner-Glindzicz M,O'Toole EA,et al.Specific loss of connexin 26 expression in ductal sweat gland epithelium associated with the deletion mutation del(GJB6-D13S1830[J].Clin Exp Dermatol,2005,30:688.
21 Del Castillo I.A deletion involving the connexin 30 gene in nonsyndromic hearing impairment[J].N Engl J Med,2002,346:243.
22 Del Castillo I,Moreno-Pelayo MA,Del Castillo FJ,et al.Prevalence and evolutionary origins of the del(GJB6-D13S1830)mutation in the DFNB1 locus in hearingimpaired subjects:A multicenter study[J].Am J Hum Genet,2003,73:1 452.
23 Street VA,Kallman JC,Kiemele KL.Modifier controls severity of a novel dominant low frequency Myosin VIIA(MYO7A)auditory mutation[J].J Med Genet,2004,41:e62.
24 Luijendijk MW,Van Wijk E,Bischoff AM,et al.Identification and molecular modelling of a mutation in the motor head domain of myosin VIIA in a family with autosomal dominant hearing impairment(DFNA11)[J].Hum Genet,2004,115:149.
25 Bolz H,Bolz SS,Schade G,et al.Impaired calmodulin binding of myosin-7A causes autosomal dominant hearing loss(DFNA11)[J].Hum Mutat,2004,24:274.
26 Di Leva F,D'Adamo P,Cubellis MV,et al.Identification of a novel mutation in the myosin VIIA motor domain in a family with autosomal dominant hearing loss(DFNA11)[J].Audiol Neurootol,2006,11:157.
27 Fukuoka H,Kanda Y,Ohta S,et al.Mutations in the WFS1 gene are a frequent ause of autosomal dominant nonsyndromic low-frequency hearing loss in Japanese[J].Jhum Genet, 2007,52:510.
28 Khanim F,Kirk J,Latif F,et al.WFS1/wolframin mutations,Wolfram syndrome,and associated diseases[J].Hum Mutat,2001,17:357.
29 Cryns K,Pfister M,Pennings RJ,et al.Mutations in the WFS1 gene that cause low frequency sensorineural hearing loss are small non-inactivating mutations[J].Hum Genet,2002,110:389.
30 Young TL,Ives E,Lynch E,et al.Non-syndromic progressive hearing loss DFNA38 is caused by heterozygous missense mutation in the Wolfram syndrome gene WFS1[J].Hum Mol Genet,2001,10:2 509.
31 Bespalova IN,Van Camp G,Bom SJ,et al.Mutations in the Wolfram syndrome 1 gene(WFS1)are a common cause of low frequency sensorineural hearing loss[J].Hum Mol Genet,2001,10:2501.
32 Hildebrand MS,Tack D,Deluca A,et al.Mutation in the COCH gene is associated with superior semicircular canal dehiscence[J].Am J Med Genet A,2009,149:280.
33 Kamarinos M,McGill J,Lynch M,et al.Identification of a novel COCH mutation,I109N,highlights the similar clinical features observed in DFNA9 families[J].Hum Mutat,2001,17:351.
34 Hutchin TP,Thompson KR,Parker M,et al.Prevalence of mitochondrial DNA mutations in childhood/congenital onset non-syndromal sensorineural hearing impairment[J].J Med Genet,2001,38:229.
35 Kokotas H,Petersen MB,Willems PJ.Mitochondrial deafness[J].Clin Genet,2007,71:379.
36 Li R,Greinwald JH,Yang L,et al.Molecular analysis of mitochondrial 12S rRNA and t RNAser(UCN)gene in paediatric subjects with nonsyndromic hearing loss[J].J Med Genet,2004,41:615.
37 Li Z,Li R,Chen J,et al.Mutational analysis of the mitochondrial 12S r RNA gene in Chinese paediatric subjects with aminoglycoside-induced and non-syndromic hearing loss[J].Hum Genet,2005,117:9.
38 Wang Q,Li R,Zhao H,et al.Clinical and molecular characterization of a Chinese patient with auditory neuropathy associated with mitochondrial 12S r RNA T1095C mutation[J].Am J Med Genet A,2005,133:27.
39 戴樸,于飛,康東洋,等.線粒體基因1555位點和GJB21基因及SLC26A4基因的診斷方法及臨床應用[J].中華耳鼻咽喉頭頸外科雜志,2005,40:769.
40 戴樸,劉新,于飛,等.18個省市聾校學生非綜合征型聾病分子流行病學研究(1)-GJB2 235delC和線粒體DNA12Sr RNA A1555G突變篩查報告[J].中華耳科學雜志2006,4:1.
41 Liu XZ,Han DY,Li JZ,et al.Loss-of-Function Mutations in the PRPS1 gene cause non-syndromic X-linked sensorineural deafness,DFN2[J].Am J Hum Genet,2010,86:65.
(2009-12-07收稿)
(本文編輯 周濤)
10.3969/j.issn.1006-7299.2011.01.024
R764.44
A
1006-7299(2011)01-0073-05
1 邁阿密大學醫學院耳鼻咽喉科(佛羅里達 33136); 2 海南醫學院附屬醫院耳鼻咽喉科
劉學忠(Email:xliu@med.miami.edu)
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
中國醫學影像學雜志(2021年6期)2021-08-13 08:43:36
中國生殖健康(2020年2期)2021-01-18 02:51:26
小學生導刊(2018年13期)2018-06-29 03:49:00
中國生殖健康(2018年2期)2018-01-12 13:57:51
現代檢驗醫學雜志(2016年4期)2016-11-15 02:01:14
中國現代醫學雜志(2015年26期)2015-12-23 11:04:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:44
中華皮膚科雜志(2014年4期)2014-12-19 12:55:49
中國神經精神疾病雜志(2014年1期)2014-03-01 03:23:22
