干旱和灌溉條件下少花蒺藜草分株生物量分配特征
2011-04-25 09:43:30張志新
草業科學 2011年2期
張志新,田 迅
(1.蘭州大學草地農業科技學院 甘肅草原生態研究所,甘肅 蘭州730020;2.內蒙古民族大學生命科學學院,內蒙古 通遼 028043)
少花蒺藜草(Cenchruspauciflorus)俗名刺蒺藜草、草狗子、草蒺藜、洋蒺藜狗子。禾本科蒺藜草屬,一年生草本植物[1-2]。總狀花序頂生,穗包藏在由多數不育小枝形成的球形刺苞內,每個刺苞含2 粒種子,少數含3粒種子。莖稈扁圓形,基部屈膝或橫臥于地面而于節上生根,且分蘗新枝條[3]。原產于北美洲及熱帶沿海地區,耐旱、抗寒、耐貧瘠、耐修剪、抗病蟲害,具有極強的適應性和競爭力,適于在沙質土壤中生長[4-6]。常生長于低海拔地區的耕地、荒地、牧場、路旁、草原、沙丘、河岸和海濱沙地等。繁殖方式以有性繁殖為主[7-8]。少花蒺藜草繁殖能力強、傳播途徑多。作為生態入侵種,幾年之內便可形成優勢種[5],與其他植物爭奪水分、土壤養分。由于種子的倒刺會對牲畜造成傷害,該草種一般對農牧業生產具有危害作用。但其適口性良好且適于在沙地生長,故有一定的飼用價值,并可用于防風固沙,對沙漠化治理和生態恢復建設具有重要的意義。
植物的表型可塑性是植物適應異質環境的生態表征。張慧榮和楊持[9]研究了羊草(Leymuschinensis)在不同條件下的可塑性變化,發現干旱條件下羊草的根莖和莖節數都有所增加。王靜等[10]、張學勇等[11]通過研究放牧干擾下冷蒿(Artemisiafrigida)和2個生境結縷草(Zoysiajaponica)無性系種群構件特征得出,生物構件的數量特征反映了植物的適應機制,表型可塑性是植物與異質環境長期相互作用的結果。目前,有關少花蒺藜草的研究主要集中在對農牧業危害的調查及防控方面[2,5,12],還沒有很好的方法能徹底防治。有關少花蒺藜草生物構件的數量特征及其對環境的適應機理報道較少。本研究以科爾沁沙地不同水分條件下的少花蒺藜草為研究對象,從構件水平初步探討其生長分配的可塑性變化和規律,為揭示該植物適應環境變化的對策和合理利用及控制提供科學依據。
1 材料與方法
1.1研究區自然概況 研究區位于通遼市南郊南沙坨子。地理位置43°36′ N, 122°22′ E,海拔178.5 m;屬典型溫帶大陸性季風氣候地帶。春季干旱多風,夏季炎熱,秋季涼爽,冬季干冷;降水多集中在6-8月。年降水量350~450 mm,無霜期150 d,年平均氣溫5.8 ℃。研究樣地地勢起伏,少花蒺藜草呈塊狀和帶狀分布,在個別生境中為單一優勢種。多在車轍中分布,也偶見于道路兩旁,生長狀況各異。研究樣地植物多以旱生為主,多年生植物主要有小葉錦雞兒(Caraganamicrophylla)、麻黃(Ephedraequisetina)、差巴嘎蒿(A.halodendron)、蘆葦(Phragmitescommunis)等[13-14],與少花蒺藜草相伴而生。個別地段有人工林分布。植物蓋度為30%~50%。土壤類型為沙質土壤,部分地段有灌溉。
1.2取樣方法 2007年10月15日,將研究區區分為干旱和灌溉2個微生境,對成熟期少花蒺藜草進行取樣。干旱微生境無灌溉。灌溉微生境地勢較低,并自7月15日起每10 d灌水1次,直至取樣。對于每一生境,設置樣方隨機對少花蒺藜草種群進行取樣。樣方大小為1 m×1 m,共設4次重復。每重復隨機選取9個植株。將所選植株連同根部一并挖出,輕輕抖落根部所附沙土,保持植株的完整,分別放入干凈塑料袋內,標記、封口,帶回實驗室進行測定。
1.3測定與方法 將少花蒺藜草植株樣品的根部剪下,清洗干凈,吸干水分。測定每一樣品地上部分的長度,將樣品依葉片、葉鞘、莖和果穗分開,用分析天平(精確度為0.000 1 g)稱量,分別裝入紙袋中并做好標記。然后將所有構件樣品置于80 ℃烘箱中烘48 h,稱烘干質量。分別計算根、葉片,葉鞘、莖和果穗構件的生物量占植株總生物量的比例。
1.4數據處理與分析 應用Excel 2003軟件進行數據統計與繪圖。采用SPSS 15.0(SPSS Inc.,USA)軟件對2個微生境下各構件生物量之間的差異進行比較和獨立樣本T檢驗;構件生物量與分株生物量之間的關系,株高與各構件、分株生物量之間的關系以及各構件之間的關系均采用相關和回歸分析。
2 結果與分析
2.1分株高度、根和穗生物量特征 結果顯示,2個生境條件下少花蒺藜草的分株高度和穗生物量均存在顯著差異(P<0.05) (表1)。灌溉條件下,植株高度為41.20 cm,明顯高于干旱條件的33.70 cm;果穗生物量是干旱條件下的1.8倍。表明少花蒺藜草對環境差異具有顯著的適應反應。
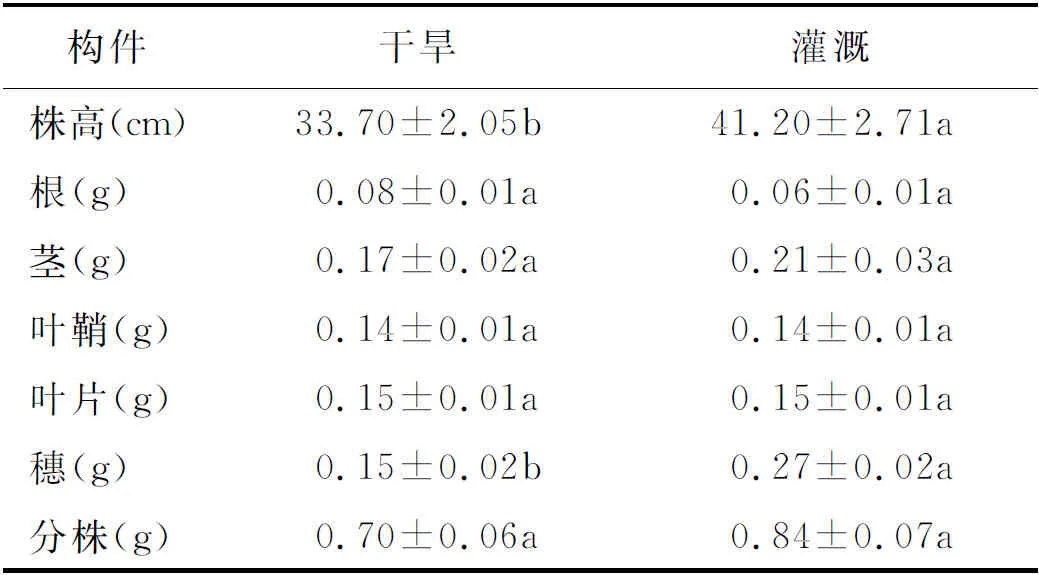
表1 不同條件下少花蒺藜草種群分株的數量特征
2.2構件生物量與分株生物量的關系 相關分析結果表明,2種條件下,根、莖、葉片、葉鞘生物量均與分株生物量呈極顯著的正相關(P<0.01),灌溉條件中相關系數(r)依次為:0.777、0.931、0.928和0.921;干旱條件中相關系數(r)依次為0.561、0.908、0.835和0.947。
回歸分析及曲線擬合的結果表明,根、莖、葉片、葉鞘生物量與分株生物量的關系符合y=axb冪函數的變化規律。在灌溉條件下,各方程的冪值(b)均高于干旱條件下相應冪值,葉片生物量的b值差異最大。表明隨著分株生物量的增加,灌溉條件下各構件的生物量增長速度均高于干旱條件(圖1)。
2.3構件生物量與植株高度的關系 相關分析的結果表明,少花蒺藜草在不同生境條件下,根、莖、葉片、葉鞘生物量和分株生物量5個指標與分株高度均呈極顯著的正相關(P<0.01)關系,且符合y=axb冪函數的變化規律。在灌溉條件下,方程的冪值(b)均高于干旱條件中相應冪值。表明隨著分株高度的增加,灌溉條件下各構件的生物量增長速度均高于干旱條件(表2)。
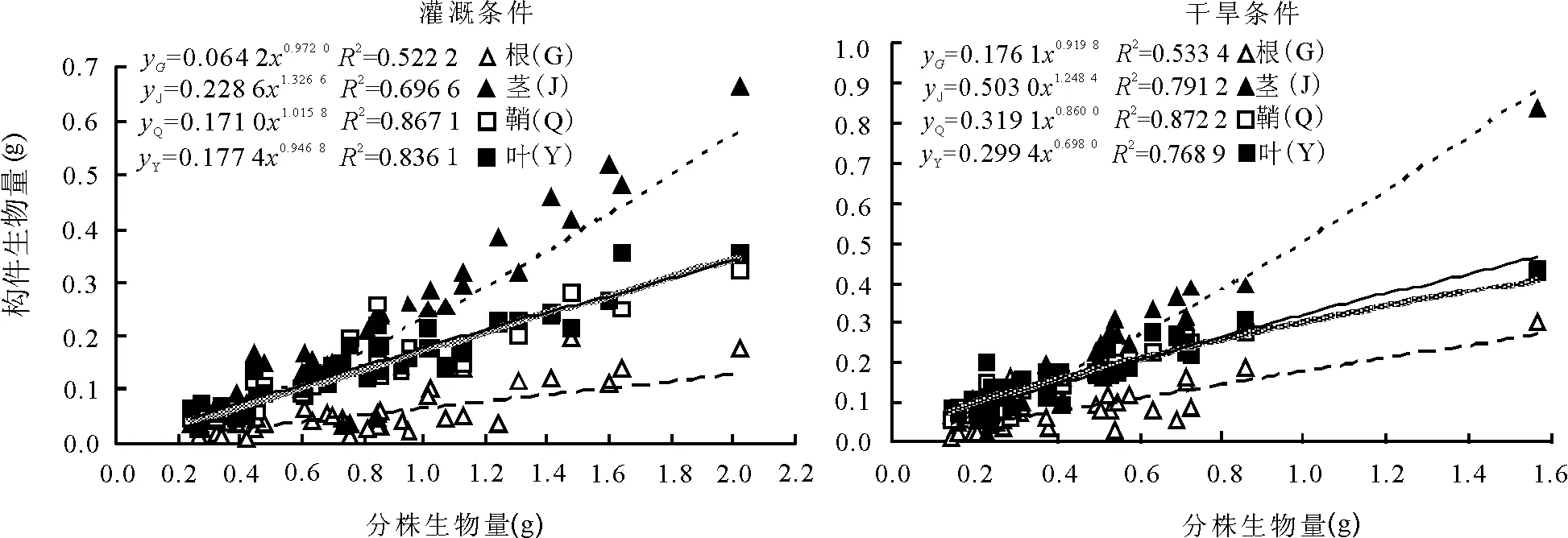
圖1 2種條件下少花蒺藜草分株生物量與各構件生物量的關系
2.4構件生物量之間的關系 相關分析結果顯示,灌溉條件下,根、莖、葉片和葉鞘生物量之間均呈極顯著的正相關(P<0.01);干旱條件下,根與葉鞘生物量、莖生物量、葉片生物量;莖與葉片生物量、葉鞘生物量;葉片與葉鞘生物量之間呈顯著的正相關(P<0.01)。以上結果表明,構件生長是相互關聯的。
回歸分析以及曲線擬合的結果表明,各構件之間的關系符合線性函數模型y=ax+b。擬合的線性方程參數見表3。
表3數據顯示,干旱條件下葉鞘、莖、葉片生物量基于根生物量的增長速率小于灌溉條件;而葉鞘、葉片生物量基于莖生物量的變化大于灌溉條件,葉片生物量基于葉鞘生物量的增長速率小于灌溉條件。
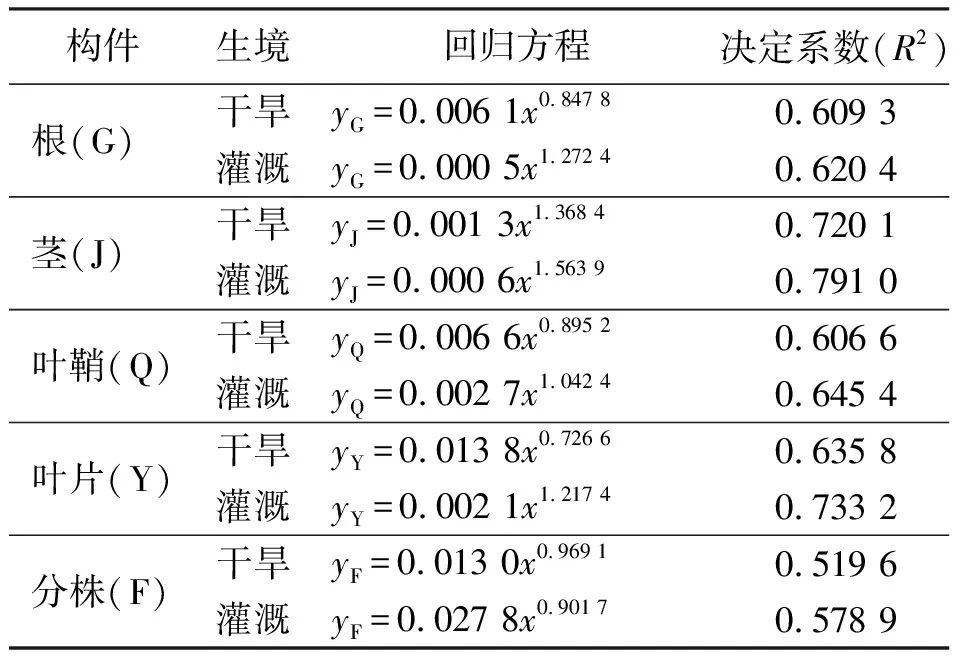
表2 不同條件下少花蒺藜草種群分株高度(y)與構件(x)生物量的關系
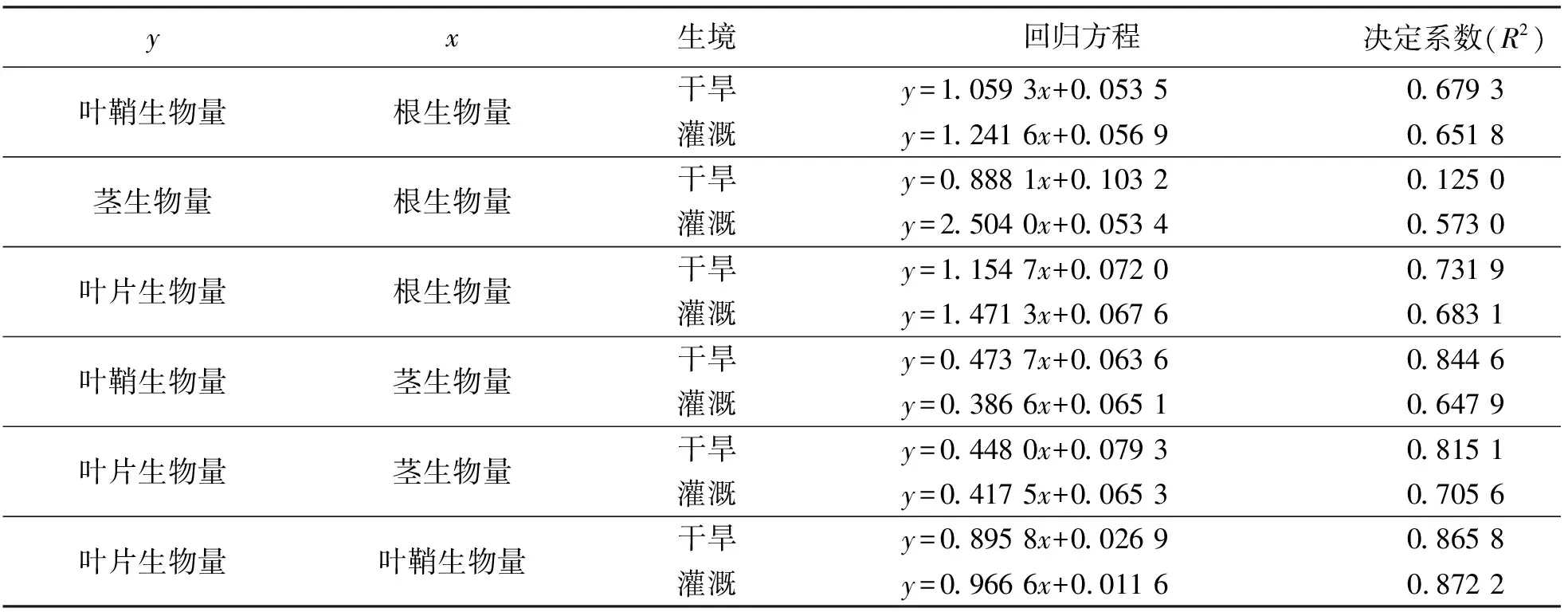
表3 不同條件下少花蒺藜草種群構件生物量之間的關系
3 討論
結果顯示,少花蒺藜草的生長和繁殖性能在灌溉和干旱條件下表現出一定的差異。干旱條件下,少花蒺藜草分株高度降低、穗生物量變小,在物質分配上表現出的是存活對策。灌溉條件下,少花蒺藜草分株高度增加、穗生物量變大,在物質分配上表現出的是競爭和繁殖生長[15]。這與王齊等[16]對水分脅迫下結縷草種群進行研究后得出的株高與生物量隨水分脅迫的增加而降低的結論一致。少花蒺藜草在2個生境條件下所表現的差異,體現出少花蒺藜草應對不同環境條件的生態適應對策特征。
少花蒺藜草根、莖、葉片、葉鞘生物量與分株生物量緊密關聯。在灌溉條件下,隨著分株生物量的增加,各構件的生物量增長速度均高于干旱條件,且以葉片的增長速度為最大;隨著分株高度的增加,各構件的生物量增長速度均高于干旱條件。這一結果表明少花蒺藜草在灌溉條件下的物質分配原則是優先建造葉器官與其他植物進行競爭。2種生境條件下少花蒺藜草各構件生物量之間也是緊密關聯的。有所不同的是,基于根的生長,在干旱條件下少花蒺藜草向葉鞘、莖、葉片生長的物質分配要小于灌溉條件;基于葉鞘的生長向葉片生長的物質分配要小于灌溉條件,基于莖的生長向葉片和葉鞘生長的物質分配要大于灌溉條件。這些物質分配特征從不同側面反映了少花蒺藜草適應干旱環境的生態響應機制。
通過本研究的結果和分析,可以看出在物質分配上,干旱條件下少花蒺藜草表現出的是存活對策;灌溉條件下表現出的是競爭和繁殖生長。
[1] Umberto Q.CRC World Dictionary of Grasses[M].Boca Raton,FL:Taylor & Francis,2006:442-443.
[2] 可欣,張秀玲,劉柏,等.彰武縣少花蒺藜草發生情況及防除技術[J].雜糧作物,2006,26(1):39-40.
[3] 唐昆.外來入侵生物少花蒺藜草[J].湖南農業,2006(5):16.
[4] Kucera C L.The Grasses of Missouri[M].Missouri:University of Missouri Press,1961:61-62.
[5] 杜廣明,曹鳳芹,劉文斌,等. 遼寧省的少花蒺藜草及其危害[J].中國草地,1995(3):71-73.
[6] 邱月,莊武, 曲波,等.少花蒺藜草遼寧省分布現狀、存在問題及防控建議[J].農業環境與發展, 2009(3):56-57.
[7] 王巍,韓志松.外來入侵生物——少花蒺藜草在遼寧地區的危害與分布[J].草業科學,2005,22(7):63-64.
[8] McCarty L B,Everest J W,Hall D W,etal.Color Atlas of Turfgrass Weeds[M].Chelsea:AnnArbor Press,2001:269-270.
[9] 張慧榮,楊持.不同生境條件下羊草種群構件的可塑性變化[J].內蒙古大學學報(自然科學版),2008,39(5):321-324.
[10] 王靜,楊持,王鐵娟,等.冷蒿(Artemisiafrigida)種群在放牧干擾下構件的變化[J].生態學報,2006,26(3):960-965.
[11] 張學勇,楊允菲,邵奎龍,等.遼東半島不同生境結縷草無性系種群構件生物量結構[J].草業科學,2006,23(4):78-81.
[12] 趙艷,呂林有,王巍,等.苜蓿不同播種行距對防控少花蒺藜草的效果[J].草業科學,2010,27(4):78-81.
[13] 李明,蔣德明,押田敏雄,等.科爾沁沙地人工固沙群落草本植物種群生態位特征[J].草業科學,2009,26(8):10-16.
[14] 左小安,趙哈林,趙學勇,等.科爾沁沙地不同恢復年限退化植被的物種多樣性[J].草業學報, 2009,18(4):9-16.
[15] 周嬋,楊允菲.松嫩平原兩個生態型羊草葉構件異速生長規律[J].草業學報,2006,15(5):77-78.
[16] 王齊,孫吉雄,安淵,等.水分脅迫對結縷草種群特征和生理特性的影響[J].草業學報, 2009,18(2):33-38.
