蝦夷扇貝養殖群體的遺傳力估算
2011-03-14 06:06:24鄭懷平李莉張國范
海洋科學 2011年3期
梁 峻,鄭懷平李 莉張國范
(1.中國科學院 海洋研究所,山東 青島 266071;2.中國科學院研究生院 北京 100039)
在數量性狀遺傳研究中,遺傳力的估算具有重要的意義。一方面它指出了在所研究的群體及所處的環境中,在一個性狀的表型變異上遺傳因素和環境因素的相對重要性。另一方面對于育種值估計、選擇指數的制定、選擇反應預測、選擇方法比較以及育種規劃決策等具有重要的作用。
許多研究工作者利用回歸分析、選擇反應和全同胞(或半同胞)對貝類的遺傳力進行了分析。主要包括長牡蠣(Crassostrea gigas)[1]、美洲牡蠣(Crassostrea virginica)[2-3]、歐洲食用牡蠣(Ostrea edulis)[4]、硬殼蛤(Mercenaria mercenaria)[5-6]、紅鮑(Haliotis rufescens)[7]、皺紋盤鮑(Haliotis discus hannai)[8]。
蝦夷扇貝(Patinopecten yessoensisJay)作為優良的海水養殖對象,自 1982年引進中國后,一直是我國北方最重要的海水養殖種類之一[9-10]。特別是近年來隨著蝦夷扇貝遺傳育種工作開展,對其遺傳參數的評估日趨重要。為此,作者以蝦夷扇貝養殖群體為研究對象,在2007年采用平衡巢式設計建立了32個全同胞家系對蝦夷扇貝養殖群體的遺傳力進行了評估。本研究的結果能夠為蝦夷扇貝遺傳育種工作的進一步開展提供科學依據。
1 材料和方法
1.1 實驗親貝及促熟
實驗所用的親貝為來自大連長海哈仙島的浮筏養殖群體,均為形狀規則、健壯的二齡貝,于 2007年2月27日開始在莊河海洋村貝類育苗場進行促熟,整個過程遵循嚴格的生產技術流程。親貝表面清洗干凈后分別置于扇貝籠中 (每籠10層,每層4個)后放在60 m3的水池進行人工控溫促熟。親貝促熟的水溫由開始的2℃逐漸升高,日升溫0.2~0.5℃,在3、5、7℃時分別穩定3~6 d,最后升到9℃恒溫待產。促熟期間,日換水1~2次,日投喂4~8次,投喂親貝的餌料為小硅藻Nitzschia closterium30~60萬個/d,螺旋藻粉 2×106~6×106g/(mL·d),雞蛋 2×106~6×106g/(mL·d),隨水溫升高根據親貝的攝食需要逐漸增加。當性腺發育完全成熟時,可用于催產。
1.2 親貝催產、全同胞家系的建立和幼體培育
隨機選取性腺發育完全成熟的8個雄貝和32個雌貝,陰干2 h后,親貝被單獨分別放入80 L塑料桶中水溫由9℃提至13℃進行誘導產卵、排精。約30 min后親貝開始陸續排放精子和卵。每個親貝排出的卵被平均放在3個80 L的塑料桶中 (桶已編號)。之后,通過巢式平衡實驗設計,每個作為雄親親貝產生的精子與 4個作為雌親親貝產生的卵分別交配,產生了8個父本半同胞家系群和32個全同胞家系。每個家系的受精卵被被單獨地放在 80L的塑料桶中進行孵化,孵化水溫13℃、鹽度30。約72 h 后,D 型幼蟲孵出。
每個家系的幼蟲被定量后單獨地放入80 L的塑料桶中培養,幼蟲的起始培養密度被設置為 10個/mL。幼蟲培育水溫由選育時的14℃隨換水逐漸升高,直至 16℃開始恒溫培養,各家系的幼蟲被培養在相同的環境條件下,包括密度、餌料、水溫、鹽度等。幼蟲的前期餌料為金藻,后期為金藻+小硅藻,投餌量和投餌次數隨幼蟲年齡的增大而增加。每天全換水一次,換水時,桶被迅速徹底清洗。注意操作,保證家系間的隔離。
1.3 稚貝的保苗和養成
20d時幼蟲開始陸續出現眼點,當 30%左右的幼蟲出現眼點時,開始投放附著基(聚乙烯網片 2片)。在60 d時,網片被裝入60目的聚乙烯網袋中,到蝦圈進行稚貝保苗,100 d時將聚乙烯網片上的稚貝全部涮洗下來,裝入40目聚乙烯網袋中到海上掛養進入幼貝養成期。當稚貝的殼高達到 10mm以上時,被從網袋中移入扇貝暫養籠(網徑 0.5cm),每層250個;隨著扇貝的生長,每層降至 100個、至 50個。而后進入養成籠(帶有標記牌),每層30個,期間更換扇貝籠 2~3次;每次換籠時,死亡的個體被記錄、剔除,重新分層,使每層30個,從而把環境效應降至最小。
幼蟲培育、稚貝的保苗和養成過程中,任何個體沒有被隨意地丟棄。
幼蟲培育和稚貝保苗在莊河海洋村貝類育苗場的扇貝育苗車間和室外蝦池中完成;在大連長海縣哈仙島海域養成。
1.4 取樣與測量
幼蟲期,在1,5,10,15和20 d時取樣,每個家系每次取樣 50個,用4%的福爾馬林殺死,在 100×的顯微鏡下用目微尺測量殼長、殼高;稚貝期,在日齡40和60 d時取樣,每個家系每次取樣30個,在40×的顯微鏡下目微尺測量殼長、殼高;在養成期,當日齡 500 d時已達到商品規格,每個家系每次取樣 30個,用游標卡尺(精度 0.02 mm)測量殼長、殼高,電子天平稱全濕質量(精度0.1g)。
1.5 數據分析
全同胞和半同胞的方差是表型變量剖分為遺傳組分和環境組分的基礎,全同胞和半同胞之間的方差分析見表1。

表1 半同胞家系和全同胞家系遺傳力估計方差分析Tab.1 Variance analysis of esitimation of heritability for half-sib families and full-sib families
每個雄親與相同數目的雌親交配,每個雌親產生相同數目的子女時,K1=K2為每個雌親的子女數目,k3為每個雄親的子女數目。
方差分量σ2S是由于雄親組不同而引起的,這些組是由同父異母的半同胞構成,因此σ2S是半同胞協方差;方差分量σ2D是由于雌親組不同而引起的,這些組是由同父同母的全同胞構成,因此σ2D是全同胞協方差;方差分量σ2W包含了剩余的遺傳方差和環境方差。它們分別由下面的公式求得:
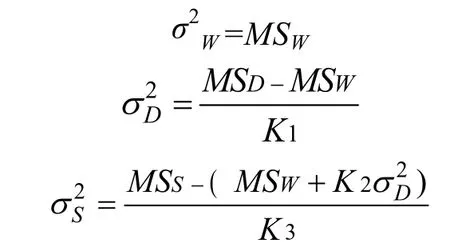
由全同胞分析可得到三個遺傳力的估計值。根據原因方差組分與協方差之間的關系,估計遺傳力的公式如下:
父本半同胞估計的狹義遺傳力為半同胞組內相關系數的4倍
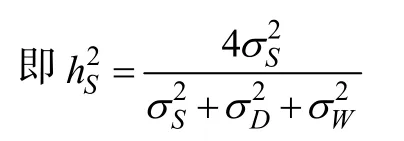
母本半同胞估計的狹義遺傳力為半同胞組內相關系數的4倍
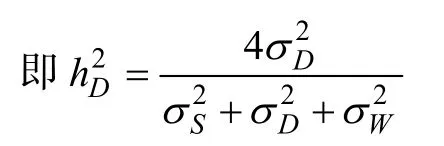
全同胞估計的狹義遺傳力為全同胞組內相關系數的4倍
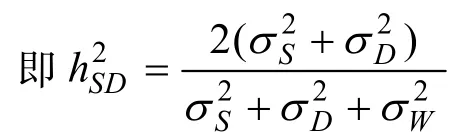
是半同胞遺傳力、h2D是全同胞遺傳力、h2SD是半同胞與全同胞的平均遺傳力。理論上,這3個遺傳力的估計值應該相等。然而,實際估計時三者間總是存在著一定的偏差。在滿足全同胞資料估計遺傳力的條件下,一般以h2SD為準,因為它是雄親組分估計和雌親組分估計的平均值。
巢式方差分析用SAS軟件基于GLM模型的限制性最大似然方法進行(SAS Institute Inc,1990)。應用統計模型為:

其中,Yijk是第i雄親與第j雌親交配而得的第k個子女的觀察值,μ為共同均數,αi為第i雄親的效應,βij為與第i雄親交配的第j雌親的效應,eijk是歸因于個體的不可控環境和遺傳離差。
所有的統計分析均用SAS8.0軟件進行,差異的顯著性都設置為P< 0.05,顯著性檢驗采用t檢驗(P< 0.05)[11]。
2 結果
2.1 表型性狀的變異系數
表2 和表3列出了在幼蟲期(1日齡、5日齡、10日齡、15日齡、20日齡)和稚貝期(40日齡、60日齡)這兩個早期發育階段表型性狀(殼長、殼高)的平均值、標準差和變異系數。變異系數值隨日齡的增長而加大,在殼長與殼高上的結果一致。這樣的結果表明隨日齡的增長,個體間差異增加。

表2 幼蟲期、稚貝期殼長的平均值、標準差(±S.D.)及變異系數(CV)Tab.2 Mean,S.D.and CV of shell lengths at larve or spat stage
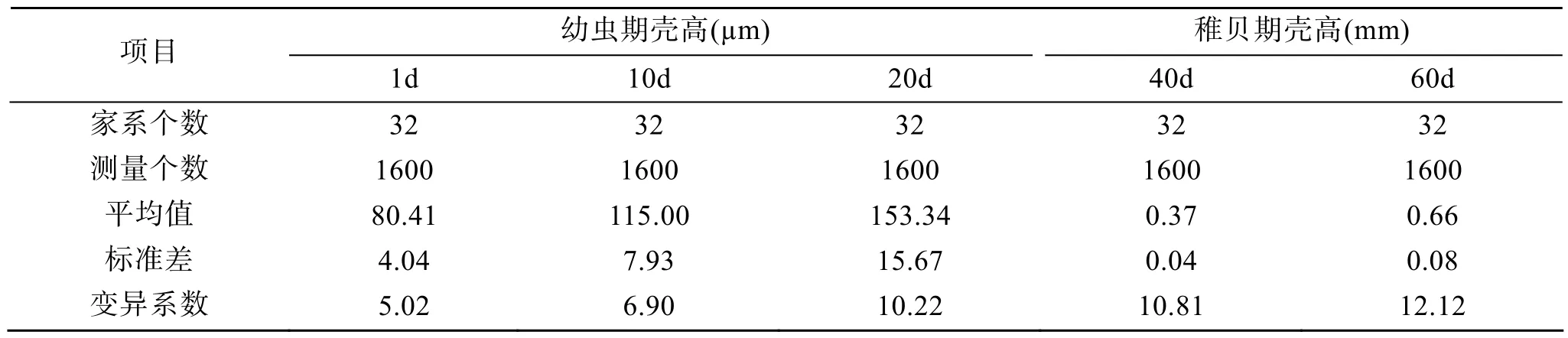
表3 幼蟲期、稚貝期殼高的平均值、標準差(±S.D.)及變異系數(CV)Tab.3 Mean,S.D.and CV of shell heights at larve or spat stage
表4列出達商品規格時測量性狀(殼長、殼高、殼寬)與稱量性狀(全濕質量)的平均值、標準差和變異系數。變異系數在測量性狀(殼長、殼高、殼寬)與稱量性狀(全濕質量)間明顯不同,測量性狀的變異系數小于稱量性狀的變異系數。這樣的結果說明蝦夷扇貝在稱量性狀(全濕質量)個體間差異比殼長、殼高、殼寬等測量性狀個體間差異更大。
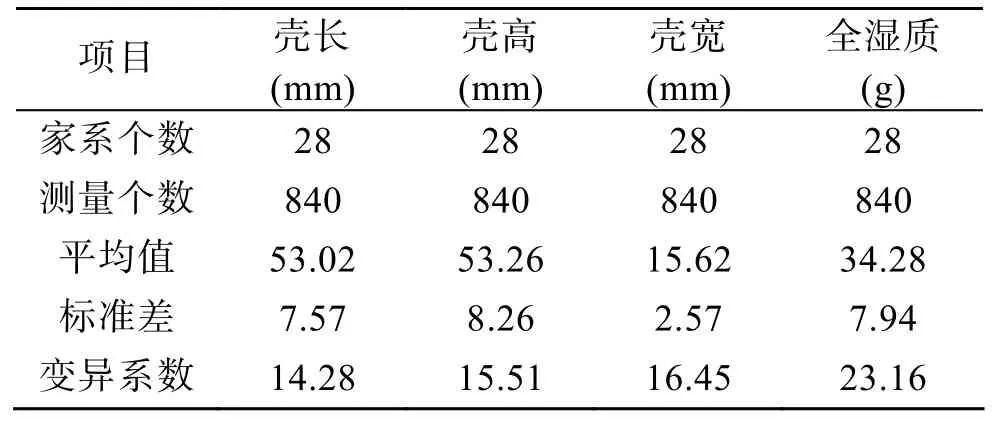
表4 達商品規格時生產性狀的平均值、標準差、變異系數Tab.4 Mean,S.D.and CV of yield traits of market standard
2.2 親本對后代的影響
方差分析表明,雄親和雌親對不同發育階段(幼蟲期、稚貝期、養成期)的測量性狀(殼長、殼高)都能夠產生顯著的影響(表5)。雄親和雌親對達商品規格時測量性狀(殼長、殼高、殼寬)及稱量性狀(全濕質量)也都能夠產生顯著的影響(表6)。

表5 不同發育階段殼長、殼高的方差組分分析Tab.5 Variance analysis of shell lengths and shell heights at different stage
2.3 表型性狀的遺傳力估計
根據表5中的方差組分估計的蝦夷扇貝在雄親和雌親對不同發育階段(幼蟲期、稚貝期、養成期)的測量性狀(殼長、殼高)的半同胞遺傳力h2S、全同胞遺傳力h2D以及半同胞與全同胞的平均遺傳力h2SD等被列在表7、表8中。由表可知,殼長的半同胞遺傳力h2S從0.186到0.364,全同胞遺傳力h2D從0.383到0.838,半同胞與全同胞的平均遺傳力h2SD從0.307到0.521,t檢驗表明殼長的半同胞遺傳力均未達到顯著水平,但全同胞遺傳力和平均遺傳力都達到顯著水平(P< 0.05);殼高的半同胞遺傳力h2S從0.171到0.381,全同胞遺傳力h2D從0.346到0.464,半同胞與全同胞的平均遺傳力h2SD從0.280到0.423,t檢驗表明殼高的半同胞遺傳力均未達到顯著水平,但全同胞遺傳力和平均遺傳力都達到了顯著水平(P<0.05)。殼長、殼高的全同胞遺傳力h2D均高于半同胞遺傳力h2S和半同胞與全同胞的平均遺傳力h2SD。在不同生長期,殼長、殼高根據父系半同胞估計的遺傳力水平逐漸增加,變化范圍分別為 0.186~0.364、0.171~0.381;根據母系半同胞家系估計的遺傳力水平逐漸變小,變化范圍分別為0.856~0.383、0.726~0.346。
根據表6中的方差組分估計的蝦夷扇貝在收獲時的殼長、殼高、殼寬、全濕質量等四個生產性狀的半同胞遺傳力h2S、全同胞遺傳力h2D以及半同胞與全同胞的平均遺傳力h2SD等被列在表9中。4個生產性狀的半同胞遺傳力h2S從0.257到0.384,t檢驗表明4個生產性狀的半同胞遺傳力均未達到顯著水平;4個生產性狀的全同胞遺傳力h2D從0.369到0.769,t檢驗表明 4個生產性狀的全同胞遺傳力均達到了顯著水平(P< 0.05);4個生產性狀的半同胞與全同胞的平均遺傳力h2SD從0.358到0.513,t檢驗表明它們都達到了顯著水平(P< 0.05)。
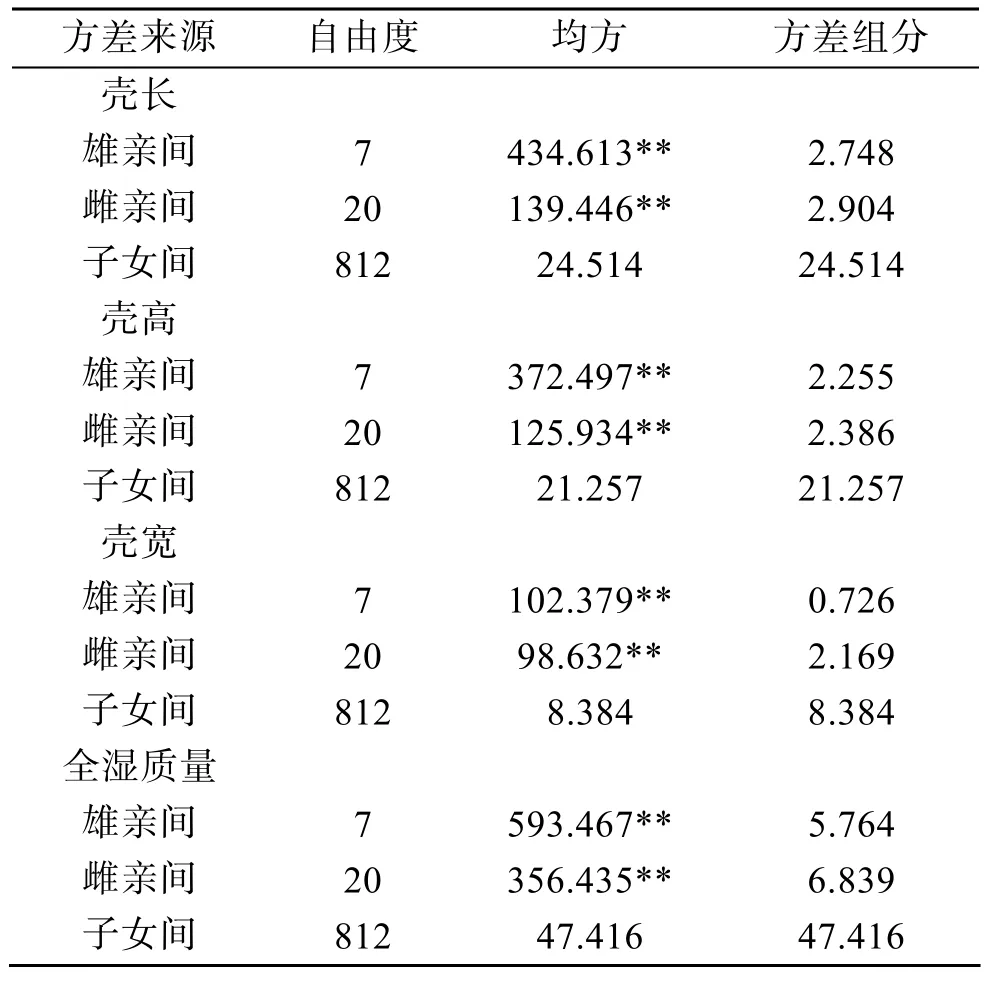
表6 成體期測量性狀與全濕質量的方差組分分析Tab.6 Variance analysis of traits and total weight at adult stage
3 討論
遺傳力是數量遺傳學中最重要、最常用的基本遺傳參數,也是數量遺傳學的最重要的特征值之一[12],在整個數量遺傳學中起著十分重要的作用。同時,估計遺傳力對遺傳育種具有重要的指導意義,它有助育種方案制定的合理化。遺傳力的大小直接影響選擇反應的效果。一般而言,基于加性方差估算的遺傳力越大,選擇反應的效果越明顯。而對于遺傳力低的性狀,選擇反應的效果不明顯。對于遺傳力較低的性狀,育種的策略為高強度選擇、家系選擇和群體(或品系)雜交。

表7 不同階段殼長的遺傳力、標準誤(±S.E.)及顯著性檢驗的t值Tab.7 Heritability,S.E.of heritability,and significance test of shell lengths at different stages
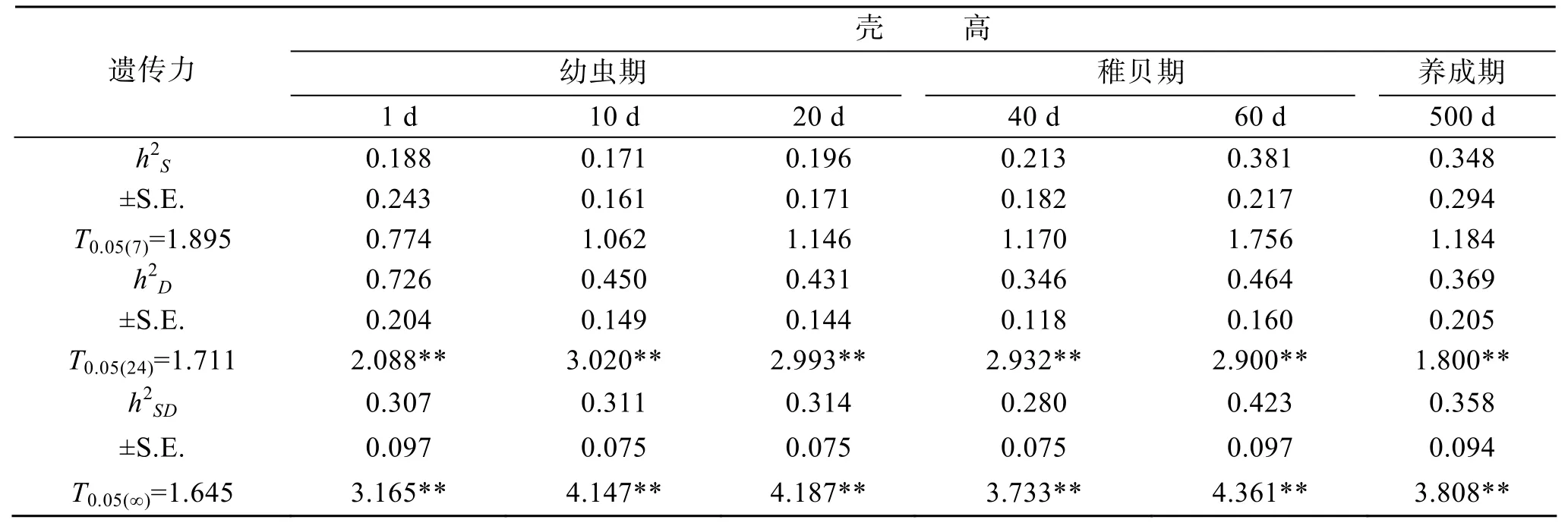
表8 不同階段殼高的遺傳力、標準誤(±S.E.)及顯著性檢驗的t值Tab.8 Heritability,S.E.of heritability,and significance test of shell heights at different stages

表9 成貝期殼長、殼高、殼寬與全濕質量的遺傳力估計Tab.9 Estimation of heritability of shell length,shell height,shell width,and total weight at adult stage
進行遺傳力的估計有助育種方案制定的合理化。估計遺傳力的方法回歸法、全同胞(或半同胞)相關法和利用選擇反應進行遺傳力的估計。有關貝類遺傳力的報道較多。國內外利用同胞家系估計貝類遺傳力的研究一直都有報道。Lannan[1]利用同胞家系估計長牡蠣C.gigas在 18月齡的體重和肉體重的遺傳力分別為 0.33和 0.37,幼體存活率和附著率的遺傳力分別為0.31和0.09。Newkirk[2]估計6和16日齡美洲牡蠣C.virginica幼蟲的半同胞遺傳力分別為 0.39和 0.50,全同胞遺傳力分別為 0.51和 0.60;Davis[13]估計美洲牡蠣C.virginica幼蟲殼長的半同胞遺傳力為0.44(選擇群體)和0.14(未選擇群體),7月齡體重與殼長的半同胞遺傳力分別為 0.51和 0.10;Rawson 等[5]利用同胞分析法估計硬殼蛤M.mercenaria在稚貝 9月齡時殼長的半同胞遺傳力為0.91,全同胞遺傳力為 0.57,半同胞與全同胞的平均遺傳力為0.74;Hilbish等[6]利用巢式設計估計硬殼蛤M.mercenaria殼長在受精后第2、10和270天的半同胞遺傳力為 1.08、0.82和 0.85,全同胞遺傳力為0.67、1.28和0.91。此外,Jonasson[7]采用不平衡設計估計紅鮑Haliotis rufescens四月齡成活率的遺傳力為0.11,8月齡和17月齡殼長的遺傳力分別為 0.08和 0.30。鄧岳文[8]利用巢式平衡設計估計皺紋盤鮑Haliotis discus hannai10日齡、20日齡、30日齡殼長的半同胞遺傳力從0.232到0.365、全同胞遺傳力從0.175 到0.747、平均遺傳力從0.202到0.489,殼寬的半同胞遺傳力從0.181到0.316、全同胞遺傳力從0.104到0.604、平均遺傳力從0.142到0.428。從這些報道中不難發現,貝類遺傳力的變化幅度比較大。本研究結果同樣發現蝦夷扇貝的遺傳力有較大的變動范圍,如達到商品規格時,4個生產性狀(殼長、殼高、殼寬、全濕質量)的半同胞遺傳力h2S從0.257到 0.384,全同胞遺傳力h2D從 0.369到 0.769,半同胞與全同胞的平均遺傳力h2SD從0.358到0.513,t檢驗表明它們都達到了顯著水平(P< 0.05)。
本研究采用巢式設計和父系半同胞相關分析法對蝦夷扇貝不同發育階段(幼蟲期、稚貝期、養成期)殼長和殼高的遺傳力進行了估計,殼長遺傳力估計范圍為0.307~0.521,殼高遺傳力估計范圍為0.280~0.423。與其他貝類相比,在蝦夷扇貝生長發育過程中,殼長和殼高的遺傳力為中等大小。
遺傳力不僅是性狀的特征而且也是群體和個體所處環境的特征。遺傳力的估值取決于各方差成分的數值。其中,遺傳方差受研究群體的基因頻率影響;環境方差受培育條件的限制。當指某性狀的遺傳力時,它應屬于特定的條件下的特定群體。因此,對于不同種或者同種不同地理群體,估算的遺傳力具有一定的差異。此外,根據全同胞家系估計的遺傳力,其中包含了顯性效應和母本效應。許多參考文獻報道了在貝類幼體發育過程中,存在較大的顯性效應和母本效應。一般認為,基于全同胞資料估計的遺傳力,較遺傳力實際值偏大,其中包含了顯性效應和母性效應,這種設計方法難以克服其自身缺陷。本研究采用巢式設計和父系半同胞相關分析法,克服了用全同胞資料估計遺傳力的缺點,采用隨機抽樣,代表性強,估計結果精確合理。
在本研究中,基于家系內的個體進行遺傳力的估算。然而,在所測定的生長階段隨著生長,家系內個體間的差異增加,從而導致環境方差比例增加和加性方差比例下降,因此隨著生長,殼長和殼高的遺傳力逐漸降低。遺傳力的大小直接影響選擇反應的效果。一般而言,基于加性方差估算的遺傳力越大,選擇反應的效果越明顯。而對于遺傳力低的性狀,選擇反應的效果不明顯。
[1]Lannan J E.Estimating heritability and predicting response to selection for the Pacific oysterCrassostrea gigas[J].Proc Natl Shellfish Assoc,1972,62:62-66.
[2]Newkirk G F.Interaction of genotype and salinity in larvae of the oysterCrassostrea virginica[J].Marine Biology,1978,48:227-234.
[3]Mallet A L,Haley L E.General and specific combining abilities of larval and juvenile growth and viability estimated from natural oyster populations[J].Marine Biology,1984,81:53-59.
[4]Toro J E,Newkirk G F.Divergent selection for growth rate in the European oyster Ostrea edulis:response to selection and estimation of genetic parameters[J].Mar Ecol Prog Ser,1990,62,219-227.
[5]Rawson P D,Hilbish T J.Heritability of juvenile growth for the hard clamMercenaria mercenaria[J].Marine Biology,1990,105:429-436.
[6]Hilbish T J,Winn E P,Rawson P D.Genetic variation and covariation during larval and juvenile growth inMercenaria mercenaria[J].Marine Biology,1993,115:97-104.
[7]Jonasson J,Stefansson S E,Gudnaso A,et al.Genetic variation for survival and body size during the first life stages of red abalone,Haliotis rufescens,In Iceland [R].Monterey,California:Third International Abalone Symposium,1997.
[8]鄧岳文.皺紋盤鮑數量性狀遺傳于育種[D].青島:中國科學院海洋研究所,2005.
[9]王慶成.蝦夷扇貝的引進及其在我國北方增養殖的前景[J].水產科學,1984,3(4):24-27.
[10]張福綏,何義朝,馬江虎.蝦夷扇貝的引種、育苗及試養[J].海洋科學,1984,1(5):38-45.
[11]Becker.數量遺傳學手冊[M].區靖祥譯.北京:科學出版社,1985.
[12]Falconer D S.Introduction to Quantitative Genetics[M].New York:Longman Inc,1981.
[13]Davis C V.Estimation of narrow-sense heritability for larval and juvenile growth traits in selected and unselected sub-lines of eastern oyster,Crassostrea virginica[J].Journal of Shellfish Research,2000,19:613.
