產α-半乳糖苷酶菌株的篩選、鑒定及其酶學特性的研究
2011-02-09 07:18:28董巖巖張燕婕黃遵錫周峻沛唐湘華李俊俊
飼料工業 2011年14期
董巖巖 張燕婕 黃遵錫 周峻沛 唐湘華 李俊俊
α-半乳糖苷酶能專一性催化α-D-半乳糖苷鍵的水解,可以作用于具有α-半乳糖苷結構的碳水化合物,故稱α-半乳糖苷酶。具有α-D-半乳糖苷鍵結構的碳水化合物稱α-半乳糖苷類物質。由于單胃動物腸道內缺乏α-半乳糖苷酶,所以α-半乳糖苷類物質在小腸內不能被消化、吸收,進入大腸后,其被大腸桿菌等微生物發酵利用,產生揮發性脂肪酸及CO2、H2和少量的CH4氣體,引起脹氣、嘔吐、下痢等現象,因此,α-半乳糖苷類物質又稱抗營養因子。在飼料中添加α-半乳糖苷酶,不僅可以降解不溶性的低聚糖,同時還可以消除腸道的脹氣現象,增加動物的采食量。因此,對α-半乳糖苷酶在飼用添加劑中的研究具有非常重大的意義[1-5]。此外,α-半乳糖苷酶在醫藥、食品工業等領域也有廣泛應用,特別是近年來α-半乳糖苷酶在血型改造、異種器官移植及法布里病等方面的應用,顯示出誘人的前景[6-9]。為了滿足人們生產生活的要求,針對微生物易培養、繁殖快、產物易收集和提純等特點,我們選擇研究微生物源的α-半乳糖苷酶。
由于本試驗中的α-半乳糖苷酶是作為酶制劑應用到飼料中,所以需要對該酶進行酶學性質的研究。在飼料加工過程中不可避免地出現的高溫,如制粒、膨化過程中的溫度可達70~150℃以上[10],從而對酶制劑的應用效果產生影響,因此就需要進行酶的熱穩定性的研究。在消化道中,胃的pH值為2.0~3.0,酶在體內起催化作用的部位是小腸,其pH值為6.0左右[11],因此就需要對該酶進行pH值耐受性研究。作為飼料,還要添加多種無機離子,對酶活性有一定的影響,所以還需要研究金屬離子對酶活的影響。
綜合目前有關微生物源α-半乳糖苷酶方面的文獻發現,不同菌種,或同一菌種在不同培養條件下產酶水平相差較大,酶學性質也有差別,其中霉菌類的產酶水平最高。因此,利用篩選培養基獲得酶活力較高的一株霉菌并對其所產的α-半乳糖苷酶的酶學性質進行了初步探討,旨在為α-半乳糖苷酶在飼料與食品中的應用研究提供參考。
1 材料與方法
1.1 材料
1.1.1 樣品
不同豆制品作坊附近的土樣。
1.1.2 主要試劑
對硝基酚、對硝基酚-α-D-吡喃半乳糖苷(pNPG)2 mmol/l、檸檬酸—磷酸氫二鈉緩沖液(pH值3.0~8.0)、Tris堿、SDS、EDTA、PCR試劑。
1.1.3 主要儀器
實驗室專用超純水機CRM-420型、電子天平PL203型、隔水式電熱恒溫箱PYX.DHS型、電熱烘干箱101-2型、分光光度計JH722s型、數顯恒溫水浴鍋Hll-2型、小型臺式高速離心機SW-CJ-2FD型、搖床HZ-9310K型、基因擴增儀EDC-810等。
1.1.4 培養基
種子培養基為(g/100 ml):PDA培養基。
初篩培養基為(g/100 ml):Peptone 0.1、NaCl 0.1、豆粕0.5、樣品1、瓊脂1.6,dH2O 100 ml。
復篩培養基為(g/100 ml):棉籽糖0.5、(NH4)2SO40.5、瓊脂1.6,dH2O 100 ml。
發酵培養基為(g/100 ml):豆粕2、(NH)2SO42、KH2PO40.2、MgSO4·7H2O 0.2,dH2O 100 ml。
1.2 方法
1.2.1 產α-半乳糖苷酶菌株的篩選
在無菌條件下,用無菌雙蒸水對樣品進行逐級梯度稀釋(10-1~10-7),吸取10-5、10-6及10-73個梯度各100 μl溶液分別涂布初篩固體培養基上,30℃下培養4~7 d;用接種環將培養出的不同菌種從初篩培養基轉接到復篩培養基上培養,檢測其是否具有α-半乳糖苷酶活性;然后將有α-半乳糖苷酶活性的菌株在PDA培養基上進行劃線分離,并按上述方式重復純化兩次后得到純菌株;將純化后的菌種依次轉接到發酵培養基中,30℃、150 r/min的條件下培養一定的時間后,對其發酵液進行α-半乳糖苷酶酶活測定;將有活性的菌株加入15%(v/v)甘油后,于-70℃下保存。
1.2.2 酶活力測定pNPG法
1.2.2.1 原理
利用對硝基酚-α-D-吡喃半乳糖(pNPG)與α-半乳糖苷酶反應后,生成有色物質對硝基苯酚(pNPH),測定其含量即可計算出酶活。
1.2.2.2 方法
先加400 μl 2 mmol/l底物pNPG和500 μl緩沖液,在55℃、pH值5.0條件下預熱5 min,再加入100 μl酶液反應10 min后,加0.5 mol Na2CO35 ml終止反應,在405 nm下測定OD值。根據標準曲線的公式計算酶活力。
1.2.2.3 酶活單位定義
在一定條件下,每分鐘分解底物生成1 μmol對硝基酚所需的酶量為一個α-半乳糖苷酶酶活單位(IU/ml)。
1.2.2.4 標準曲線的制作
以吸光值為橫坐標,對硝基苯酚含量為縱坐標繪制標準曲線,每組設三個平行,充分搖勻后于405 nm處比色,0號管為對照管。
對硝基苯酚標準曲線的繪制見表1。

表1 對硝基苯酚標準曲線
1.2.3 D-1基因組總DNA的提取
①將菌種接種于30 ml的PDA液體培養基中,30℃培養2~3 d;
②滅菌紗布過濾得菌絲球,用無菌水沖洗菌絲球2次;
③加入液氮研磨3次后,立即加入提取緩沖液,65℃裂解5 min,12 000 r/min離心10 min,取上清;
④上清液等體積與氯仿混勻,上下搖勻2次,12 000 r/min離心10 min,取上清;
⑤重復步驟④;
⑥上清液加1/10體積的醋酸鈉(3 mol/l,pH值5.2)和2倍體積的無水乙醇,-70℃放置2 h,4℃、12 000 r/min,離心20 min,沉淀;
⑦加1 ml 70%的乙醇洗鹽,4℃、12 000 r/min,離心10 min,重復一次;
⑧37℃開蓋放置10~20 min控干;
⑨加入30 μl TE-RNaseA,37℃放置30 min以上,取5 μl提取物于0.8%瓊脂糖凝膠電泳檢測提取效果,剩余的提取物-20℃保存備用。
1.2.4 D-1 18SrDNA PCR擴增體系及條件
以提取的D-1基因組為模板,用真菌的ITS1和ITS4引物來擴增D-1的18S rDNA,引物序列為:
ITS1:5’-GGAAGTAAAAGTCGTAACAAGG-3’
ITS4:5’-TCCTCCGCTTATTGATATGC-3’
取5 μl PCR產物進行0.8%瓊脂糖凝膠電泳檢驗。
擴增體系為無菌ddH2O 35.5 μl;10×PCR緩沖液5.0 μl;dNTP混合液(2.5 mmol/l)4.0 μl;引物ITS1(10 pmol/l)2 μl;引物ITS4(10 pmol/l)2 μl;模板DNA 1 μl;TaqDNA聚合酶0.5 μl;總體積50 μl。
擴增條件為94℃預變性4 min、94℃變性30 s、52℃退火30 s、72℃延伸1 min,反應35個循環,最后72℃延伸10 min。
1.2.5 酶學特性的研究
1.2.5.1 溫度對酶活性的影響
以pNPG為底物,在pH值5.5,不同溫度(20、30、35、40、45、50、55、60、70、80℃)下反應10 min,測定相同的酶液在不同的溫度下所具有的酶活力單位,酶活最高的為最適反應溫度。
1.2.5.2 pH值對酶活性的影響
以pNPG為底物,溫度為55℃,不同pH值條件(3.0、4.0、5.0、5.5、6.0、6.4、7.0、8.0)下反應10 min,測定相同的酶液所具有的酶活力單位,酶活最高的為最適反應pH值。
1.2.5.3 α-半乳糖苷酶的pH值穩定性
將原酶液在不同pH值(3.0、4.0、5.0、5.5、6.0、6.4、7.0、8.0)的緩沖液中于37℃下保溫1 h,在最適條件下測定酶殘余活性以研究酶的pH值穩定性。以未處理的酶液的酶活為100%計。
1.2.5.4 α-半乳糖苷酶的熱穩定性
將原酶液在不同溫度(40、50、55、60℃)保溫30、60 min,在最適條件下測定酶殘余活性以研究酶的熱穩定性。以未處理的酶液的酶活為100%計。
1.2.5.5 金屬離子對酶活力的影響
用濃度為0.01 mol/l的金屬離子Mg2+、Na+、Ca2+、K+、Fe3+、Al3+、Co2+、Cu2+、Zn2+、Pb2+、Mn2+、EDTA分別與原酶液以1∶1的比例進行混合,37℃保存1 h后,分別在最適條件下進行酶活的測定。以未處理的酶液的酶活為100%計。
2 結果與分析
2.1 酶活力計算公式
根據pNPH標準曲線(見圖1)得到回歸方程,從而得到α-半乳糖苷酶酶活力。
酶活力(IU/ml)=(0.350 4X+0.000 4)×n/(T×V)。式中:X——吸光度;
n——酶液稀釋倍數;
V——稀釋后反應中的加酶量(ml);
T——酶-底物反應時間(min)。
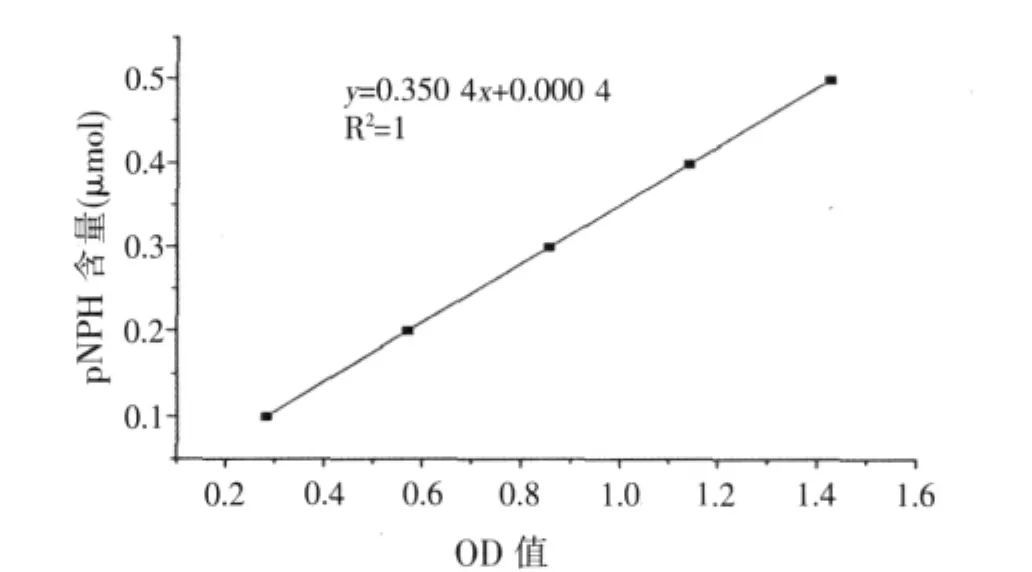
圖1 標準曲線
2.2 產α-半乳糖苷酶菌種的篩選(見圖2)
利用篩選培養基從20個不同豆制品作坊附近的土樣中篩選得到7株產α-半乳糖苷酶的菌株(如圖2),從中選取一株產α-半乳糖苷酶酶活最高的菌株D-1并進行酶學特性研究。
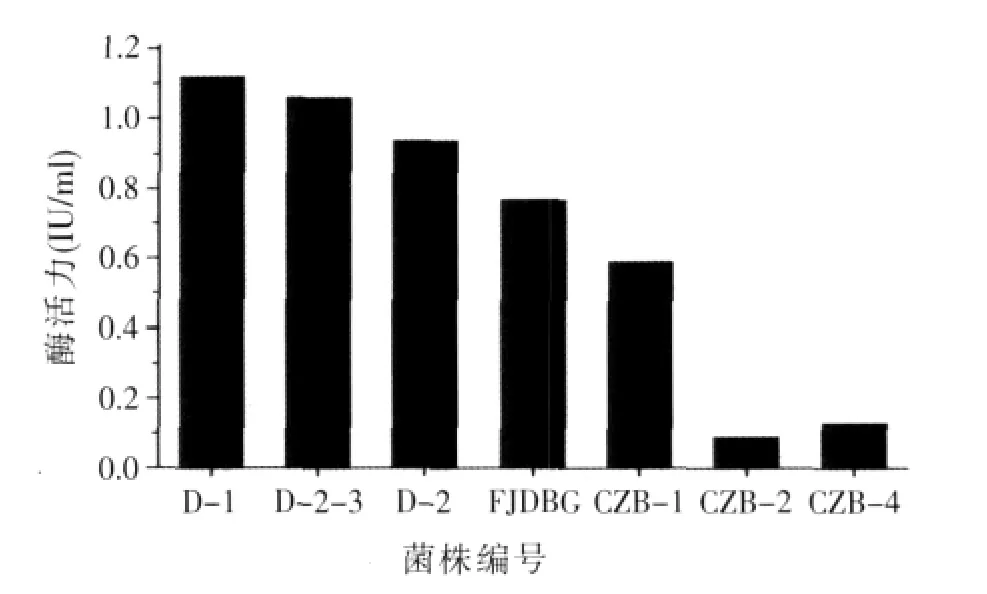
圖2 不同菌株酶活力的比較
2.3 α-半乳糖苷酶的酶學性質研究
2.3.1 溫度對酶活力的影響(見圖3)
在不同溫度梯度、pH值5.5的條件下,以pNPG為底物,反應10 min后,測定酶活。結果如圖3,酶活性隨溫度的升高而提高,在最適反應溫度處酶活力最高,超過這一溫度,酶活力便急劇下降。因此,α-半乳糖苷酶的最適作用溫度為55℃,當溫度超過55℃,酶活顯著降低。
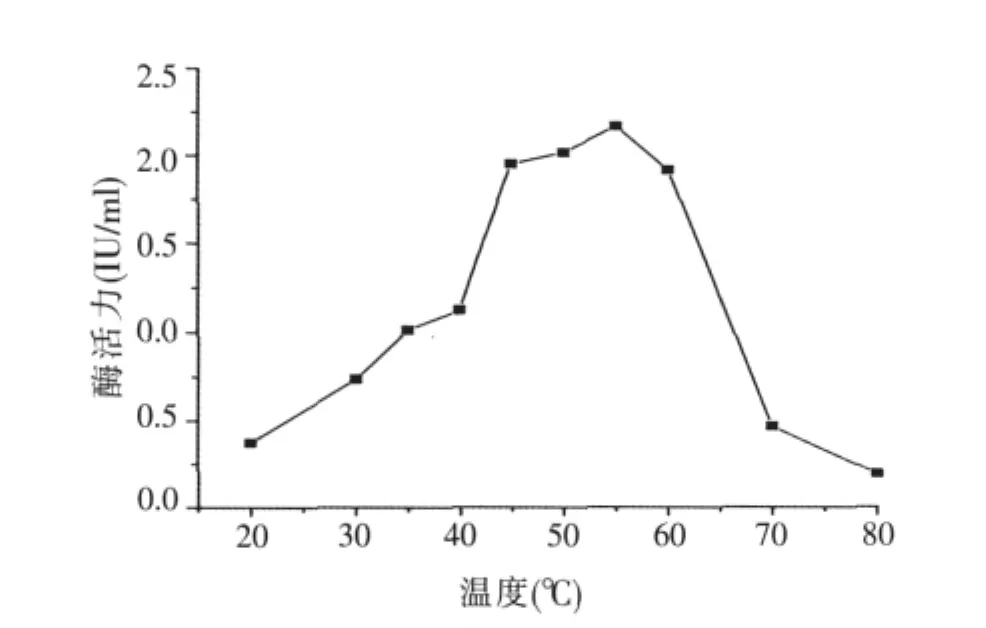
圖3 溫度對酶活力的影響
2.3.2 pH值對酶活力的影響(見圖4)
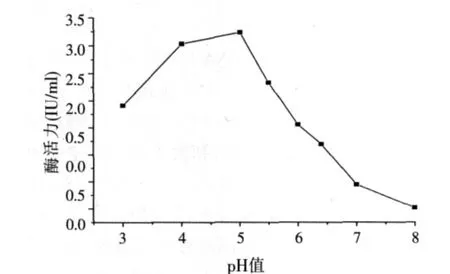
圖4 pH值對酶活力的影響
在不同pH值、55℃條件下,以pNPG為底物,反應10 min后,測定酶活。結果如圖4,α-半乳糖苷酶在pH值3.0~5.5之間表現較高的酶活力,說明該酶較耐酸,并且其反應最適pH值為5.0左右。堿性條件則不同程度地抑制酶活。
2.3.3 α-半乳糖苷酶的pH值穩定性(見圖5)
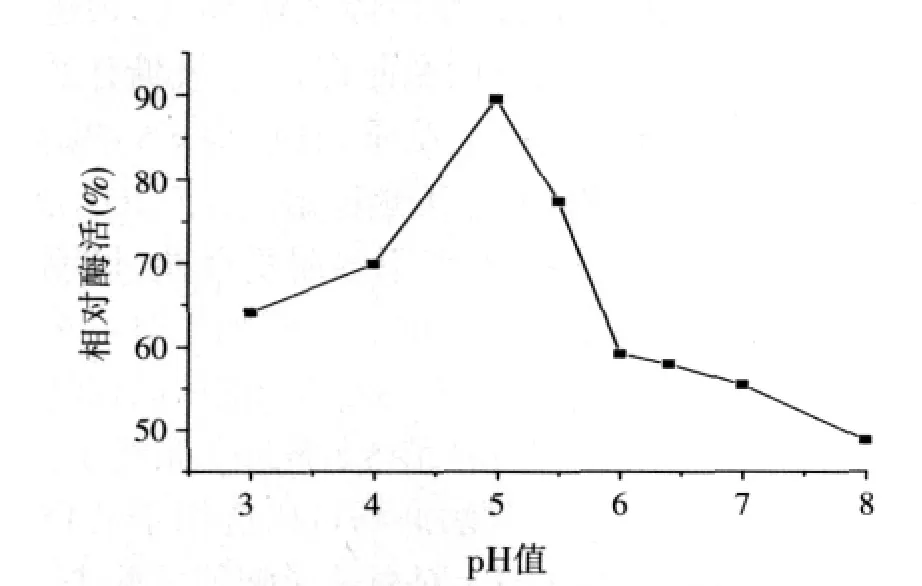
圖5 α-半乳糖苷酶的pH值穩定性
由于動物消化道食糜的pH值變化范圍比較大,所以單純考慮在某一pH值條件下的酶活值并不全面,最好能測定該酶在比較廣泛的pH值范圍內的酶活表現。因此,將酶液在不同pH值的緩沖液中于37℃下保溫1 h,在55℃、pH值5.0條件下測定酶殘余活性。結果如圖5,表明在pH值3.0~5.5范圍內保持64.1%以上的酶活,其中在pH值5.0條件下耐受1 h酶活只下降了11%左右。在消化道中,胃的pH值為2.0~3.0左右,酶在體內起催化作用的部位是小腸,其pH值為5.0~6.0左右。所以該酶在動物腸道中可發揮其作用,能很好的應用到動物飼料中。
2.3.4 α-半乳糖苷酶的熱穩定性(見圖6、圖7)
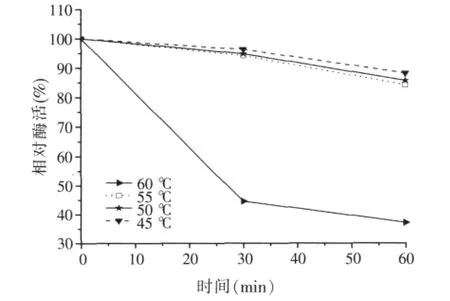
圖6 α-半乳糖苷酶的熱穩定性
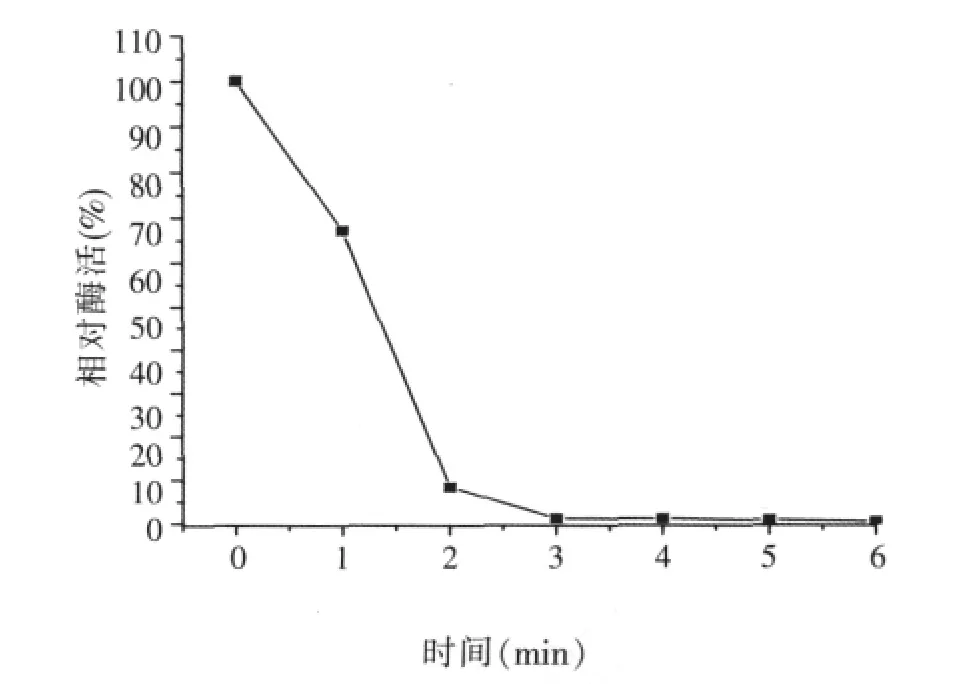
圖7 α-半乳糖苷酶的熱穩定性(70℃)
將酶液分別在45、50、55、60℃下,保溫30、60 min,隨著溫度的升高和放置時間的延長,酶活保留率降低(如圖6),在60℃以下存放穩定性較好。在45、50、55℃中耐受30 min至60 min,酶活下降趨勢平緩。60℃保溫0.5 h,酶活剩余44.6%,60℃保溫1 h,酶活剩余37.2%。此外,還做了在70、80℃下的耐受性試驗,70℃下1 min時酶活剩余67.2%,2 min后酶活完全消失(如圖7),80℃下酶活完全消失,說明該菌產的酶不夠耐熱。飼料加工制粒、膨化過程中的溫度可達70~150℃以上,所以飼料加工時可在加熱后進行添加,如制成液體酶制劑于飼料調質制粒后在顆粒表面進行噴涂。
2.3.5 金屬離子對酶活力的影響(見圖8)
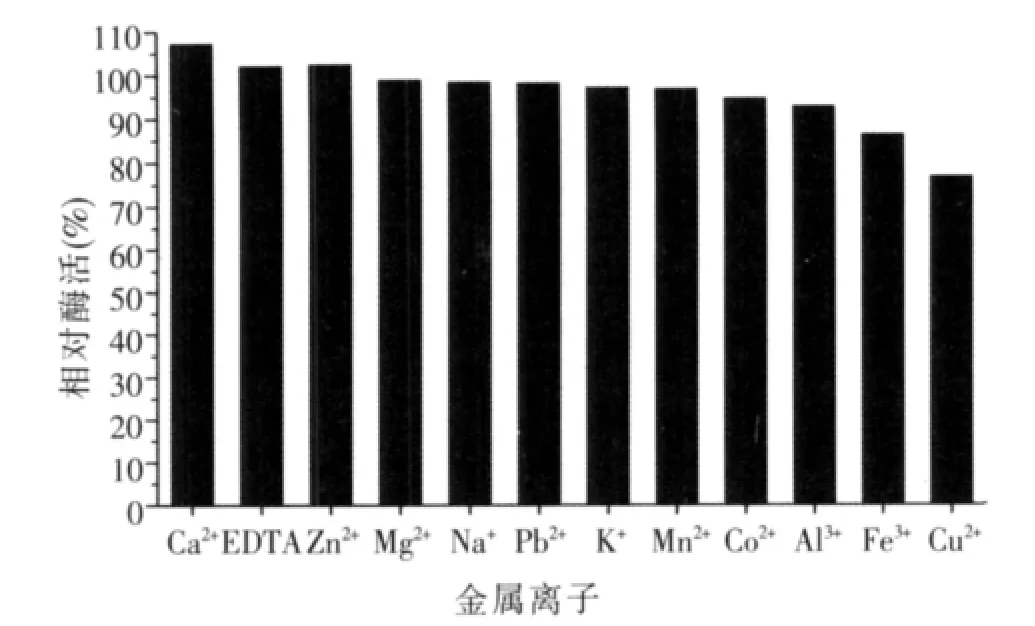
圖8 金屬離子對酶活力的影響
原酶液在不同種類的金屬離子中耐受1 h后,大多數金屬離子對α-半乳糖苷酶均具有不同程度的抑制作用,其中Cu2+和Fe3+的抑制作用較為明顯,而Ca2+、EDTA、Zn2+對α-半乳糖苷酶有一定的促進作用,所以在添加酶制劑時盡量避免與鐵器、銅器長時間接觸。
2.4 D-1基因組DNA的提取(見圖9)
提取的D-1基因組DNA經0.8%瓊脂糖凝膠電泳鑒定結果,說明提取的總DNA純度良好。
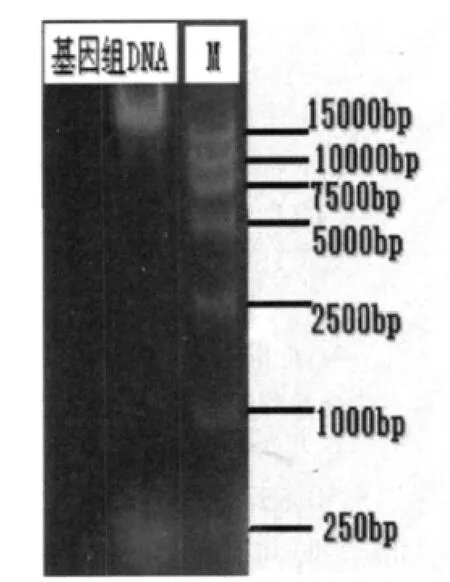
圖9 D-1基因組DNA電泳圖
2.5 D-1 18S rDNA基因獲得
用D-1的基因組DNA為模板進行18S rDNA PCR,PCR擴增產物經瓊脂糖凝膠電泳結果見圖10,PCR擴增所產生的DNA片段為單一條帶,片段大小在600 bp左右。
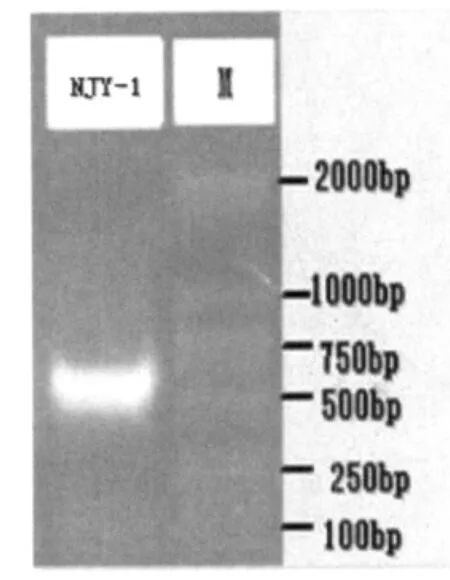
圖10 D-118S rDNA的擴增片段
2.6 D-1 18S rDNA基因的序列分析及進化樹的構建(見圖11)
測序得到完整的18S rDNA序列,將此序列與NCBI上已有序列進行Blast比對,發現其18S rDNA與多數Aspergillus niger同源性達到99%,用MEGA4和BioXM對目的序列進行比對和聚類分析構建系統進化樹,從圖11可以看出,D-1與Aspergillus niger聚為一類,且與Aspergillus niger AB369898.1有最大的同源性,遺傳距離幾乎為0,因此可初步確定D-1屬于Aspergillus niger。
3 討論
在關于產α-半乳糖苷酶的微生物的文獻中發現,霉菌等真菌類微生物的α-半乳糖苷酶的產量較高,因此,本研究直接從樣品中篩選真菌,篩選出酶活最高的一株霉菌D-1,通過對D-1進行液體發酵獲得原酶液的最大酶活為3.48 IU/ml,將粗酶液進行離心后對其進行酶學性質的研究。
經研究發現,該酶的最適反應溫度是55℃,最適反應pH值為5.0,與已報道的黑曲霉α-半乳糖苷酶的最適反應溫度是60℃,最適反應pH值為4.5的結果有差異[12]。該酶在液體狀態下熱穩定性較好,60℃以下條件都很穩定,在70、80℃下的耐受性差,說明該菌產的α-半乳糖苷酶不夠耐熱,飼料加工時可在制粒后進行液體噴涂[13];在pH值3.0~5.5范圍內保持64.1%以上的酶活,其中在pH值5.5條件下耐受1 h酶活只下降了11%左右,因此,此酶可以應用于動物飼料中。在金屬離子及EDTA對酶活影響的試驗中,發現Ca2+、EDTA、Zn2+對α-半乳糖苷酶有一定的促進作用,而Cu2+和Fe3+的抑制作用較為明顯,所以在添加酶制劑過程中盡量避免與鐵器、銅器長時間接觸。
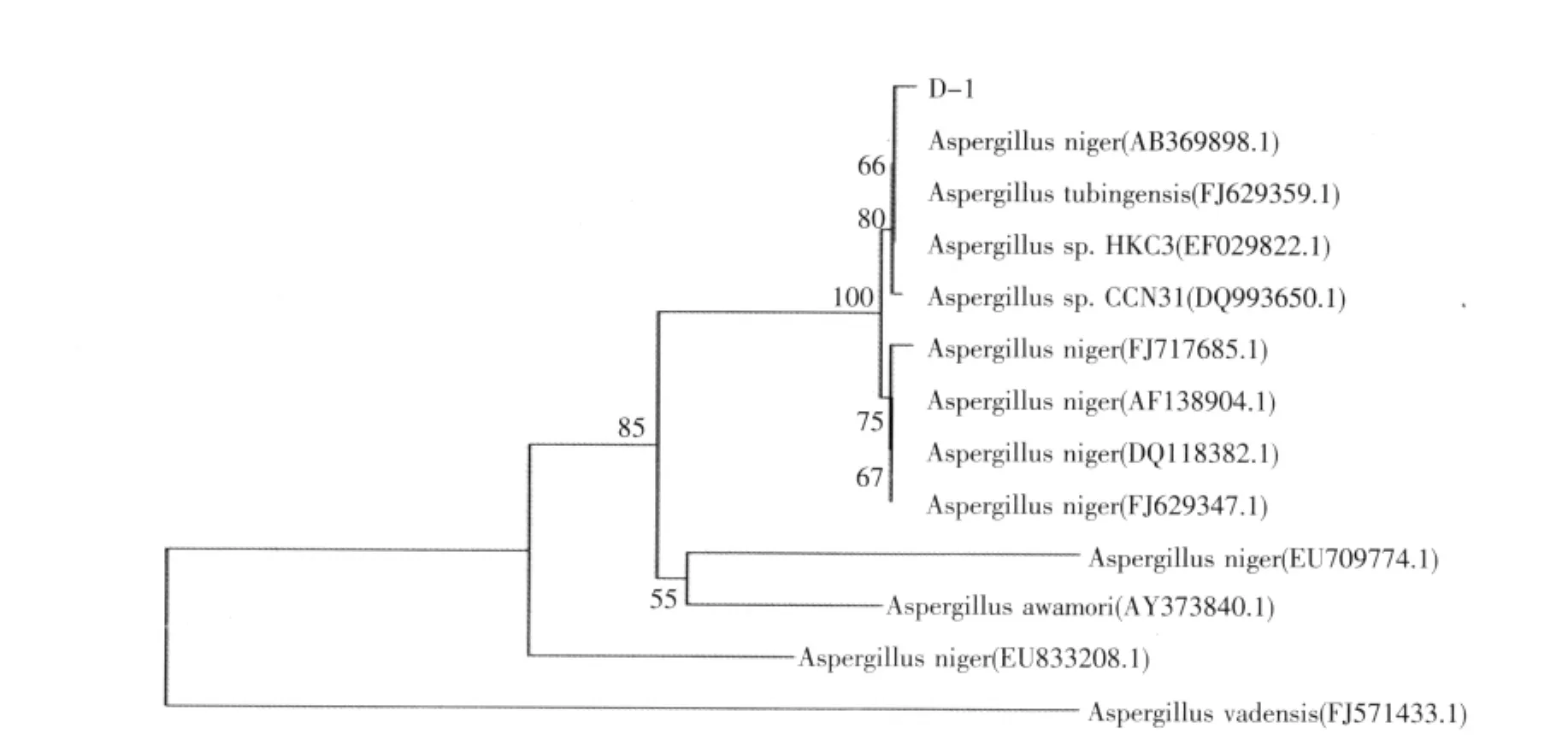
圖11 菌株NJY-1 18S rDNA的進化樹
[1] 張晉輝.畜禽日糧中的α-半乳糖苷以及相應酶制劑的應用[J].中國農業技術早報,2003,3(1):49-54.
[2] 于桂陽.α-半乳糖苷酶制劑在豬日糧中的應用[J].飼料工業,2006,27(18):46-48.
[3] 許堯興,許少春,李艷麗,等.在玉米-豆粕型日糧中添加α-半乳糖苷酶對生長豬生產性能的影響[J].飼料工業,2008,29(4):9-10.
[4] Leske K L,C.J.Jevne,C.N.Coon.Extraction methods for removing soybean alpha—galactosides and improving true metabolizable energy for poultry[J].Anim.Feed Sci.Technol.,1993(b),41:74-78.
[5] 蔣小豐,方熱軍.α-半乳糖苷酶在畜禽日糧中的應用研究[J].飼料工業,2009,30(10):18-21.
[6] 吳勁松,馮萬祥.α-半乳糖苷酶[J].生命的化學,2000,20(2):84-86.
[7] 李孝輝,陳聲明.微生物源α-半乳糖苷酶的研究進展[J].微生物學通報,2002,29(2):71-75.
[8] 高新.α-半乳糖苷酶及其在輸血等方面的應用[J].中國實驗血液學雜志,2000,8(3):236-239.
[9] 崔文.α-半乳糖苷酶的醫學研究進展[J].中外醫療,2007(19):56-57.
[10] 邱萬里.關于飼料制粒工藝中要素的控制[J].糧油加工與食品機械,2002(8):51-52.
[11] 我國飼用酶制劑應用現狀與問題[J].河北畜牧獸醫,2000,16(7):8.
[12] 許堯興,李艷麗,柳永,等.黑曲霉變種RM48α-半乳糖苷酶的分離純化及其酶學性質研究[J].浙江大學學報,2009,35(2):147-152.
[13] 史寶軍.飼用添加劑劑型制備及加工工藝技術創新[J].飼料與畜牧,2008(4):39-42.
