山桐子天然群體表型遺傳多樣性研究
2010-07-09 13:00:02李大偉龔榜初白杰建彭佳龍陳紅星
湖南農業科學 2010年11期
李大偉,龔榜初,白杰建,彭佳龍,陳紅星
(1.中國林業科學研究院亞熱帶林業研究所,浙江 富陽 311400;2.青川縣林業局,四川青川281000;3.臺州市仙居縣林業局,浙江 臺州 317300;4.金華市磐安縣林業局,浙江 金華 322300)
山桐子(Idesia polycarpa)為大風子科山桐子屬植物,在中國主要分布于長江流域、華北和西北南部的17個省區。另外,朝鮮、日本的南部也有分布。山桐子是陽性速生樹種,適應性強,對氣候要求不嚴格。山桐子結實量大,含油率高,果實成串,鮮紅艷麗形似瑪瑙,既可以作為冬季觀果的園林綠化樹種,又是良好的新型能源樹種。其油脂成分中不飽和脂肪酸的含量在80%以上,作為食用油對人體健康十分有益,并且對高血壓、高血脂等心腦血管疾病有很好的預防作用和輔助療效[1]。迄今,陜西、四川等地仍有食用山桐子油的習慣。但是對于山桐子的研究基礎十分薄弱,近年來一些學者對山桐子的栽培技術[2],果實成分[3]、組織培養[4]以及引種育苗方面[5]做過一些研究,但有關山桐子天然居群遺傳多樣性方面的研究尚未見報道。筆者以山桐子葉片、果實、果穗和種子等形態特征為研究內容,對秦嶺淮河以南、四川以及陜西等八個省份山桐子天然居群的群體遺傳結構和遺傳多樣性進行了研究,旨在為我國新一代能源樹種的資源保護和生產應用提供幫助,同時也為基因資源保存和遺傳改良提供參考。
1 材料與方法
1.1 群體選擇和試驗材料的采集
在全面收集山桐子天然分布區資料和實地勘測的基礎上,于2008年9~12月在我國山桐子自然分布區內選擇12個群體進行調查采樣。經調查和試驗獲得各居群生態與地理狀況見表1。在每個群體內,選取15~20株、10~20 a生處于盛果期的健康植株作為樣株,株間距離在200 m以上。對樣株進行定位、標記,在樣株樹冠中上部外緣隨機采集30片葉子和30穗成熟果穗做測試材料。由于山桐子種子自然萌發率極低,天然分布稀少,所以湖北宜昌、湖南通道和浙江磐安三地采集的樣品數量不足,以下主要對其他9個群體所采集樣品的數據進行分析。
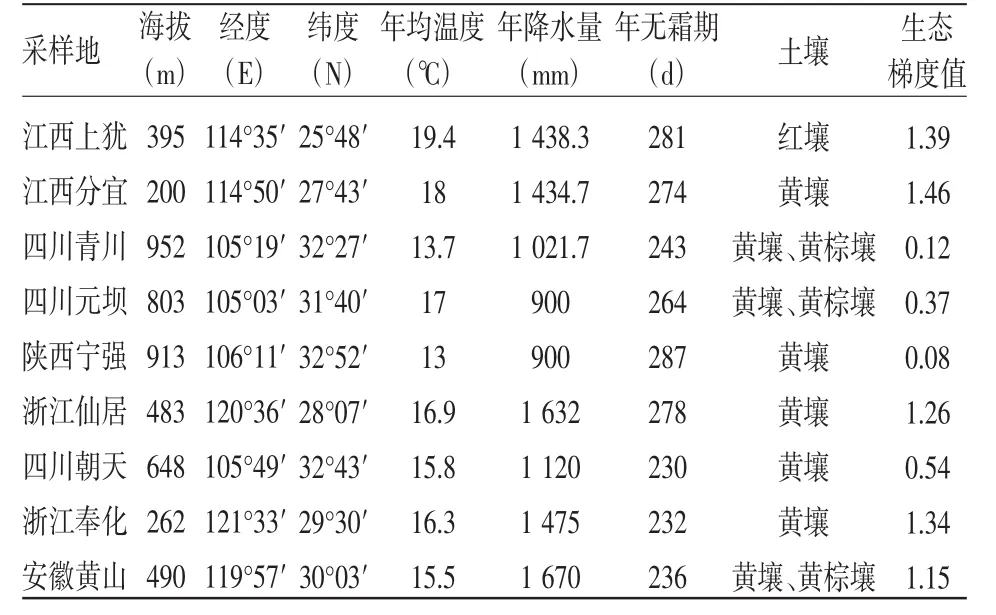
表1 山桐子采樣群體的地理位置及生態因子情況
1.2 性狀測定方法
選取相對穩定、便于測量的表型性狀,每個地點選取15株樹,每株樹選取葉片30個,果穗30串,隨即選擇果實30顆。用直尺測定葉長、葉寬、葉柄長、果穗長、果穗寬,精確到0.1 cm。用數顯游標卡尺測量葉柄徑、果長、果寬,精確到0.01 cm。定義葉長/葉寬為葉形指數,果長/果寬為果形指數。從每個單株中隨機選取50粒種子稱重,換算成千粒重。
1.3 統計分析
對各性狀采用巢氏設計方差分析[6],生態梯度綜合值(EGA)是生態因子經過CA、PCA降維的綜合值,能夠綜合反映生態諸因素的綜合效應[7]。
采用雙變量相關分析對山桐子各表型性狀和采集點的地理生態因子進行相關分析。表型數據標準化以后,利用歐式離差平方和法對表型性狀進行聚類分析。
其他統計運算按照常規方法并利用EXCEL2003,DPS11.5,SPSS16.0 軟件包提供的有關程序進行。
2 結果與分析
2.1 山桐子群體間表型性狀的形態變異特征
表2為山桐子群體間和群體內表型性狀的變異均方及F值,葉片、葉柄、果實、果穗、種子等15個表型性狀在群體間和群體內均存在極顯著差異。從各表型性狀的均值及均值的多重比較結果可知,山桐子各性狀在群體間大多存在顯著差異,葉長和葉寬變異相近,均值較大的群體均為朝天、寧強。各樣本間葉長差異顯著,而葉寬不顯著。葉形指數測量值較大的群體為奉化和黃山相互差異不顯著,較小群體為上猶和分宜相互之間差異顯著。葉柄長均值較大群體為奉化和朝天相互之間差異顯著,較小群體為元壩和黃山相互差異顯著。果穗的性狀研究結果表明,每穗出果數變異最大,各群體間均存在顯著差異,其次為果穗長。果穗寬比較穩定,各樣本間均值的變幅為5.772~9.324 cm,果實大小較為穩定,各群體內果長和果寬的均值都在8.0~9.5 mm之間,只有奉化群體的均值達到10.572 mm。其余性狀例如果形指數、果重、單果種子重、千粒重各群體間均值差異較小,只有單果出籽數這一指標各地樣本的均值變幅18.593~41.937,并且各群體間存在顯著差異。由此可見,山桐子群體間各性狀差異較大,從葉片和果實表型指標測定結果而言,采集于西北地區朝天和寧強的山桐子種源優于東部采集地。
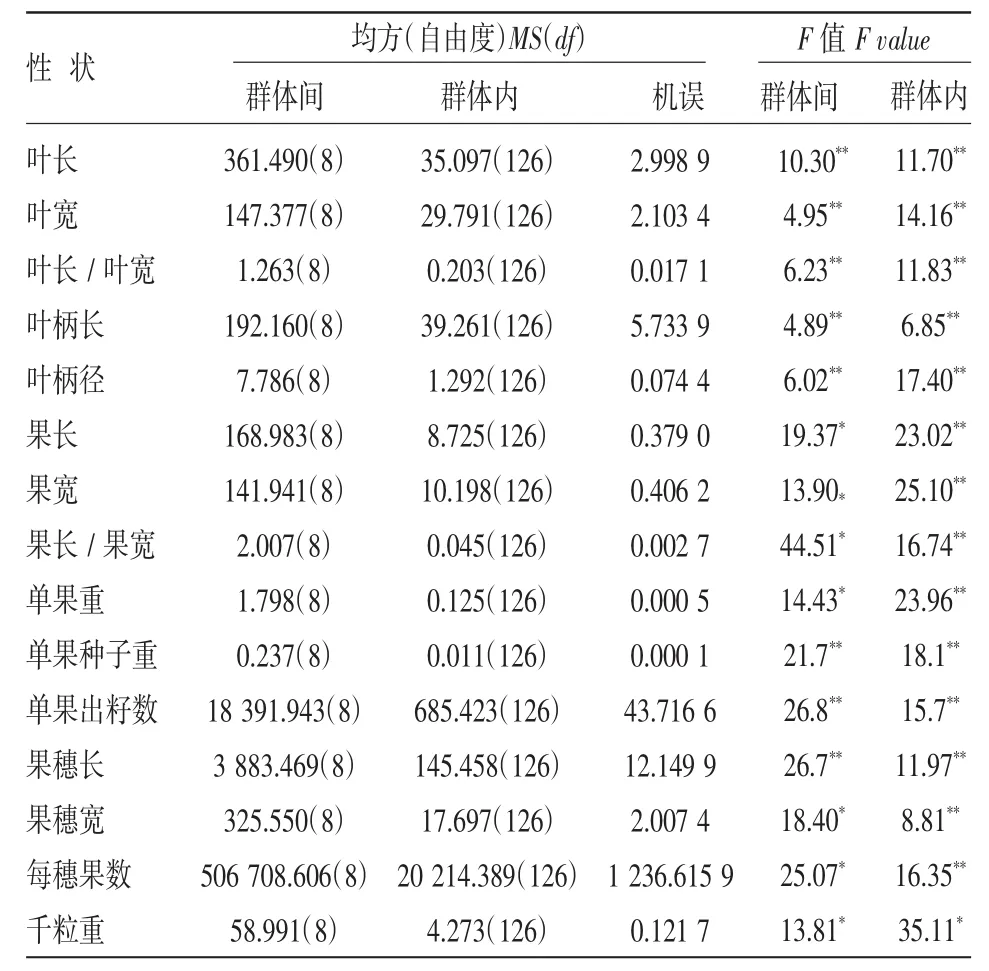
表2 山桐子群體間及群體內表型性狀方差分析結果
2.2 山桐子表型變異和地理、氣候生態因子的相關性
進一步對山桐子15個表型性狀與海拔、經緯度、年均溫、年降雨量、無霜期和生態梯度值等生態因子進行相關性分析。結果表明:果形指數與經度和年降雨量,葉片長、寬與緯度分別呈正相關關系,相關系數分別為 0.856**、0.668*、0.732*、0.794*。葉片長、寬與年均溫和EGA,單果出籽數和經度,千粒重與無霜期分別呈負相關,相關系數分別為-0.786*、-0.752*、-0.696*、-0.738*、-0.708*。15個性狀與海拔的相關性均為不顯著,以上可知,生態環境因子對山桐子表型性狀的影響主要集中在葉片長、寬,果實大小,單果出籽數和千粒重。
2.3 山桐子群體表型聚類分析
為進一步研究山桐子群體間的遺傳關系,利用歐式離差平方和法對9個天然群體的葉形指數、葉柄長、果穗長、單果重等15個表型數據進行聚類分析(圖 1)。
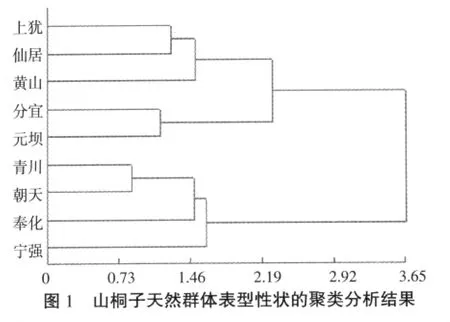
根據聚類分析結果,可將9個群體歸為3個類群。其中上猶與仙居表型性狀相近,黃山與其接近,這3個種源屬于第一類群。分宜與元壩歸為第二類群。青川與朝天距離很相近,奉化與這兩個種源接近,寧強距離稍遠但也同屬于第三類群。由圖可知,山桐子的表型性狀聚類與地理距離聚類不相一致。分宜與元壩分別屬于江西省與四川省,但山桐子的表型性狀卻很相近,具體原因還有待進一步研究。
3 結論與討論
3.1 山桐子遺傳變異豐富
本研究結果表明山桐子表型性狀存在廣泛變異,通過群體間和群體內表型性狀的變異均方及F值檢驗,所研究的葉長、寬,果實大小,果穗,種子等15個表型性狀在群體間和群體內均存在極顯著差異。各性狀指標變異系數的變幅為12.65%~74.22%。充分說明山桐子群體表型性狀變異豐富。山桐子這種變異是自身遺傳因素和環境因素共同作用的結果。這種多層次的變異為優質種質資源和生物多樣性保護提供了物質基礎,同時也表明了生物多樣性保護任務的艱巨性。
3.2 環境對山桐子遺傳變異的影響較大
山桐子在我國的17個省區都有分布,植物在與環境長期互作或生存競爭的過程中,形成了許多適應或防御機制,包括形態結構、生理生化代謝等方面,這些特性逐漸轉變為可遺傳的性質保留下來,這是植物適應環境發展的結果[8]。形態或表型特征變異反映基因型、群體或生態型的變異豐富度,表型變異是基因組的遺傳變異與環境互作的結果[9]。山桐子分布區內地形、氣候等環境生態因子復雜多變,以及通過長期的地理隔離和自然選擇,使其產生了極其豐富的種間和種內變異。本研究表明,山桐子對緯度變化表現出一定的梯度規律性,隨著緯度的變化,由南向北,葉片長和葉寬等性狀都有增加的趨勢,作為一個優良的速生陽生樹種,這也是山桐子適應環境的一種表現。
另外,研究還表明山桐子的單果重、單果出籽率、單果種子重等性狀對經度變化改變比較明顯,從東到西均有增加的趨勢,形態特征的變異往往具有適應意義,自然群體中保持較大的變異貯存對群體是有利的,因此山桐子豐富的表型變異,也進一步說明了山桐子抗逆性強、適應性廣的原因。
3.3 山桐子種質資源遺傳改良前景廣闊
本研究主要從山桐子表型變異著手,表型變異是遺傳型和環境因子共同作用的結果[10-11],而且表型變異必然蘊涵著遺傳變異,表型變異越大,可能存在的遺傳變異越大。研究自然存在的表型變異,是研究遺傳變異的先導。根據山桐子豐富的自然變異情況,可以推斷山桐子的改良前景是廣闊的。山桐子具有較高的變異水平,遺傳潛力很大。因此,在遺傳改良工作中,要兼顧優良種源、優良群體和優良個體進行選擇。
[1]楊志玲,王開良,譚梓峰.值得開發的幾種野生木本油料樹種[J].林業科技開發,2003,17(2):41-43.
[2]祝志勇.山桐子栽培試驗[J].浙江林業科技,2004,24(4):36-38.
[3]周伯川,楊 帆,薛雅琳,等.水冬瓜油工業化生產的研究[J].中國油脂,1996,21(1):12-14.
[4]蔣澤平,梁珍海,劉根林,等.山桐子莖段離體培養技術研究[J].中國農業通報,2006,22(12):393-396.
[5]梁珍海,蔣澤平,李淑琴,等.日本山桐子引種育苗及苗期生長規律研究初報[J].江蘇林業科技,2006,33(4):9-11.
[6]李 斌,顧萬春,盧寶銘.白皮松天然群體種實性狀表型多樣性研究[J].生物多樣性,2002,10(2):181-187.
[7]顧萬春,李 斌.生態梯度軸(EGA)區劃林木育種區的研究[J].生態學報,1997,17(2):5-8.
[8]龐廣昌,姜冬梅.群體遺傳多樣性和數據分析 [J].林業科學,1995,31(6):543-550.
[9]張恒慶,安利佳,祖元剛.天然紅松種群形態特征地理變異的研究[J].生態學報,1999,19(6):932-938.
[10]羅建勛,顧萬春.云杉天然群體表型多樣性研究[J].林業科學,2005,41(2):66-73.
[11]曾 杰,鄭海水,甘四明,等.廣西西南樺天然居群的表型變異[J].林業科學,2005,41(2):59-65.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
保健醫苑(2021年7期)2021-08-13 08:48:02
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
