纖維黏連蛋白碎片增加整合素α5β1在髓核細胞表達的研究
2010-05-25 01:43:42夏茂盛朱悅高靜杰
中國醫科大學學報 2010年10期
夏茂盛,朱悅,高靜杰
(1.中國醫科大學 附屬第一醫院骨科,沈陽 110001;2.遼河油田第二職工醫院 麻醉科,遼寧 盤錦 124010)
纖維黏連蛋白(fibronectin)是一種間盤細胞外基質糖蛋白,可以被基質金屬蛋白酶(matrix metalloproteinases,MMP) 分解為纖維黏連蛋白碎片(fibronectin fragments,Fn-f)[1]。在間盤退變過程中,Fnf是一種退變的產物,其含量明顯增加。同時研究表明,Fn-f又具有誘導間盤退變的作用[2]。將Fn-f注入髓核內,間盤出現了退變樣改變;將Fn-f加入間盤細胞的培養液,間盤細胞基質蛋白的合成明顯減少,而 MMP 的表達明顯增加[3,4]。然而目前 Fn-f誘導間盤退變的具體機制并不清楚。而纖維黏連蛋白和 Fn-f在細胞膜上主要的受體是整合素 α5β1[5,6]。因此,本實驗將Fn-f加入髓核細胞的培養液,觀察Fn-f對其受體整合素α5和β1亞基表達的影響,探討Fn-f誘導間盤退變的機制。
1 材料與方法
1.1 材料
動物為日本大耳白兔,3月齡,10只,體質量(2 500±200)g;試劑為纖維黏連蛋白碎片(110 kDa,Sigma)、用于免疫沉淀的整合素α5和β1亞基抗體和蛋白G瓊脂糖小球(upstate biotechnology)、用于Western blot和免疫熒光的鼠抗整合素 α5、β1、II型膠原蛋白、MMP9和MMP13的單克隆抗體(SantaCruz)。
1.2 方法
1.2.1 細胞的分離和培養:無菌取出兔間盤髓核組織,0.2%的膠原酶消化4 h,用D-Hanks進行酶液清洗3次。然后用培養液DMEM/F12+15%胎牛血清重懸細胞,以2×104/cm2接種于培養皿內,置于37℃5%CO2條件下培養。約6~7 d后,待細胞貼壁完全第1次換液,以后隔2~3 d換液,細胞生長到80%融合后,用0.25%胰酶消化傳代。第3代細胞長到70%~80%融合后,將培養液去除,實驗組加入含有Fn-f的無血清DMEM/F12培養液,Fn-f的濃度為100nmol/L,對照組則只加入無血清DMEM/F12培養液,37℃5%CO2條件下培養2 d。
1.2.2 Western免疫印跡:培養2 d后,去除培養液,加入裂解液,收集細胞裂解液,用Bradford法測各個樣品蛋白含量。比較兩組間整合素α5和β1含量,取實驗組和對照組樣品各1 000μg加入8μg抗-整合素亞型(α5或 β1)的抗體,4 °C 孵育 12 h。然后向樣品中加入200μl蛋白G瓊脂糖小球,4°C孵育2 h。離心(14 000 g,5 s)收集瓊脂糖小球,并在沸水中煮5 min。取上清加入準備好的10%SDS-聚丙烯酰胺凝膠中,電泳分離樣品。而比較II型膠原蛋白、MMP9、MMP13在兩組間含量時,樣品蛋白含量定量后,每組樣品取的蛋白量一致,均為50μg,直接將樣品加入準備好的10%SDS-聚丙烯酰胺凝膠中,電泳分離樣品。以后實驗步驟相同,電泳分離樣品后,將蛋白轉移到硝酸纖維素膜上。封閉后,在膜上分別加入鼠整合素亞型 (α5、β1)、II型膠原蛋白、MMP9 和MMP13的特異性一抗,1∶1 000稀釋,孵育2 h。洗膜1 h后,加入羊抗鼠的二抗,辣根過氧化物酶標記,1∶2 000稀釋,孵育2 h。最后纖維素膜用ECL底物顯影到膠片上,以β-actin為內參,結果用圖像分析軟件進行分析。
1.2.3 免疫熒光:細胞培養2 d以后,將培養液倒出,用PBS稍洗細胞,然后在-20°C條件下用純甲醇固定6 min。固定后的細胞經PBS稍洗細胞后,封閉30 min,然后加入一抗,鼠抗兔的針對整合素α5亞基和Ⅱ型膠原蛋白的一抗同時加入,針對整合素β1亞基和Ⅱ型膠原蛋白的一抗同時加入,4°C過夜。洗去一抗后,加入TRITC和FITC結合的二抗,室溫2 h。然后PBS洗細胞30 min,甘油封片,熒光顯微鏡觀察結果。
1.2.4 逆轉錄聚合酶鏈反應(RT-PCR):將細胞裂解后,應用Trizol等提取總核糖核酸(RNA)。用TAKARA試劑盒,將提取的RNA逆轉錄為cDNA,再進行PCR擴增,引物見表1。反應條件是94℃2 min;94 ℃ 30 s,58 ℃ 45 s,72 ℃ 1 min,共 35 個循環。用1.5%瓊脂糖凝膠電泳,分離PCR擴增產物。經紫外分光儀比較PCR產物的含量。
1.3 統計學分析
實驗結果利用SPSS12.0統計軟件進行分析,多樣本采用方差分析,以P<0.05為差異有統計學意義。
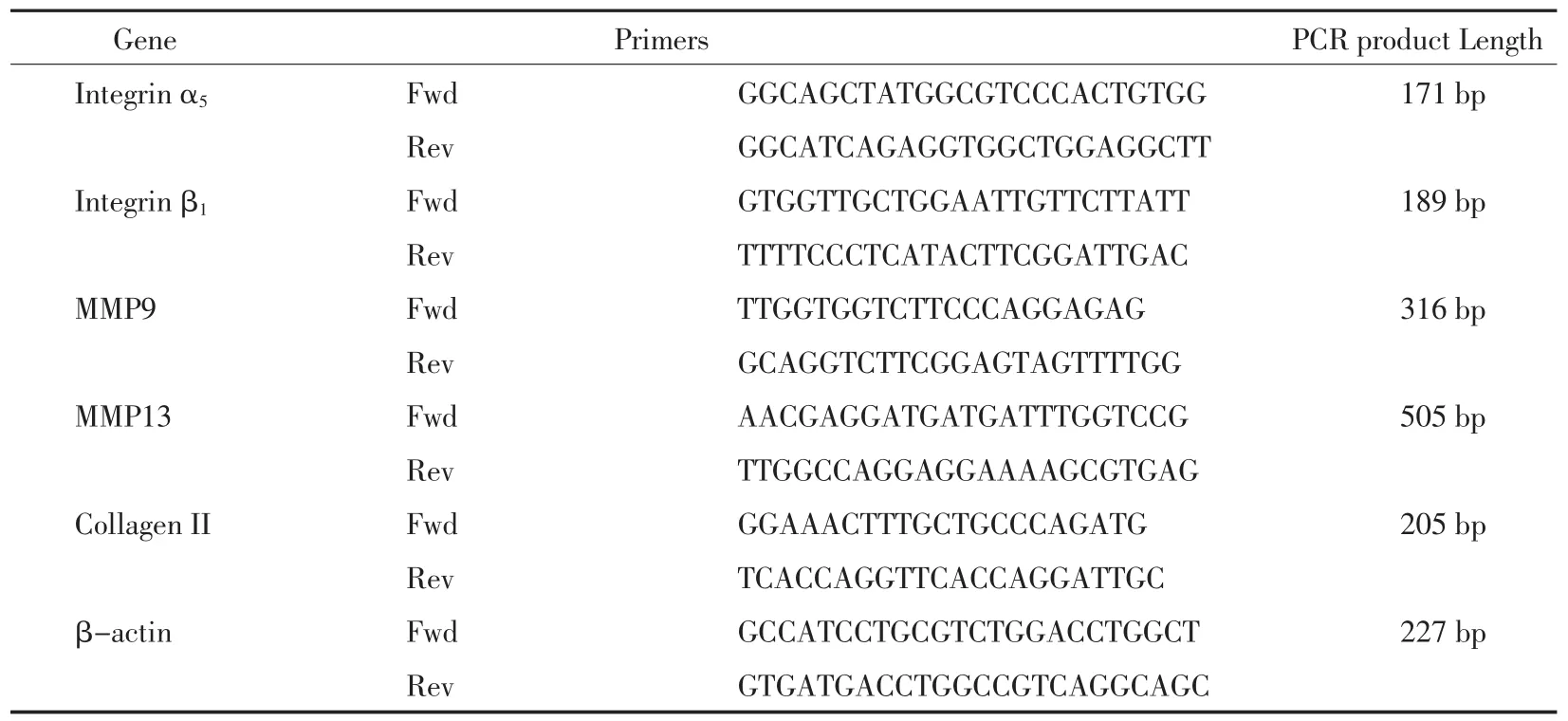
表1 用于RT-PCR的引物序列Tab.1 The sequences of the primers used for RT-PCR
2 結果
2.1 細胞形態的變化
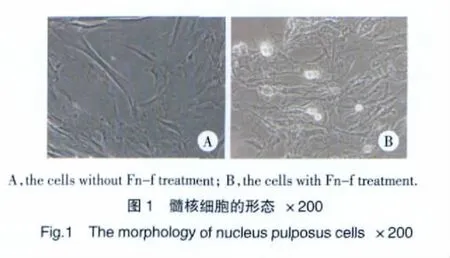
加入Fn-f后髓核細胞形態變化明顯,細胞體積縮小,有多角形轉變為細長形,并可見少量細胞碎裂,細胞內和細胞間顆粒樣物質明顯增多(圖1)。
2.2 蛋白表達的變化
經Fn-f作用后,整合素 α5、β1亞基和MMP9、MMP13的含量都明顯增加,與對照組相比均有統計學意義(P<0.05)。而Ⅱ型膠原蛋白的含量明顯下降,與對照組相比差異有統計學意義(P<0.05)(表2)。免疫熒光技術觀察到整合素α5、β1均表達在細胞膜上,經Fn-f作用后,這兩種蛋白在胞膜上的熒光強度明顯增強,而Ⅱ型膠原蛋白的熒光強度明顯降低(圖 2)。
2.3 基因表達的變化
經 Fn-f作用后,整合素 α5、β1亞基、MMP9、MMP13的表達均明顯升高,而Ⅱ型膠原蛋白的表達明顯降低(圖3)。
3 討論
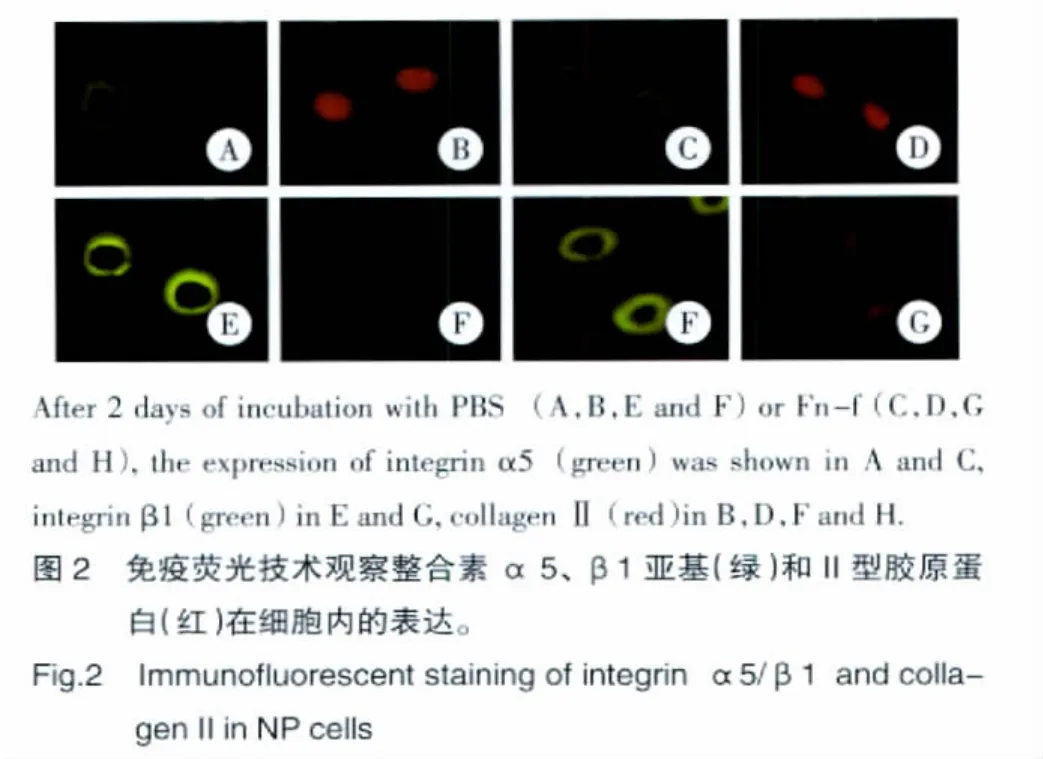
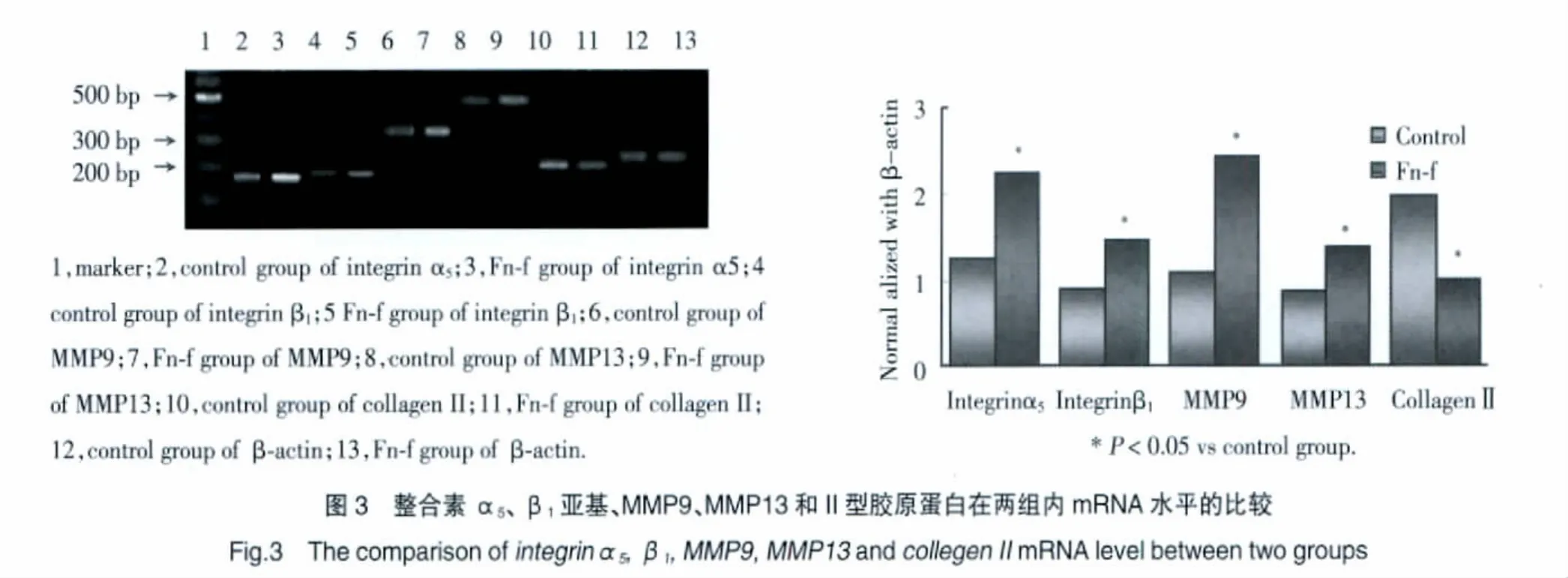
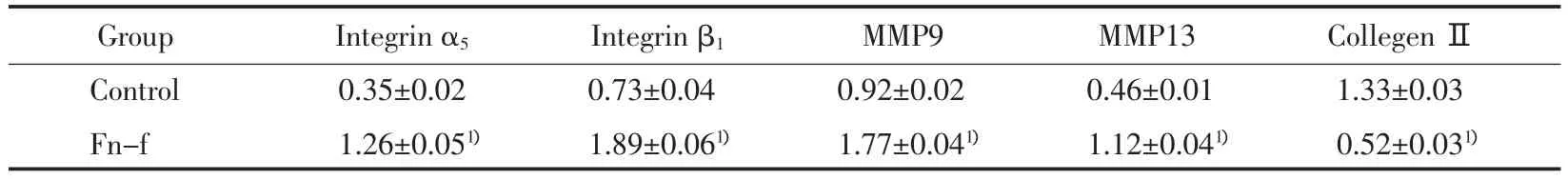
表2 兩組整合素α5、β1亞基、MMP9、MMP13和II型膠原蛋白含量的比較Tab.2 The comparison of integrinα5,β1 subunits,MMP9,MMP13 and collegen IIprotein content
本實驗發現,Fn-f可以誘導間盤髓核細胞退變。在加入Fn-f后,髓核細胞的Ⅱ型膠原蛋白表達降低,而MMP9和MMP13的表達明顯升高。在Fn-f的干預下,整合素α5β1的表達增加,這說明Fn-f有促進其受體表達增加的作用。
Fn-f是間盤退變過程中纖維黏連蛋白的分解產物,在退變間盤內的含量明顯增加[8]。究其原因有兩方面,一方面是纖維黏連蛋白在退變間盤內的表達增多[9,10],另一方面在退變過程中MMP的表達明顯增加,而正是MMP將纖維黏連蛋白分解為Fn-f[11]。體內和體外實驗都證明了Fn-f有減少糖蛋白表達,促進基質蛋白分解的作用[2~4]。本實驗同樣證明了這個結論,Fn-f減少了II型膠原蛋白的表達,增加了MMP9和MMP13的表達。但Fn-f誘導間盤退變的具體機制并不清楚。而Fn-f同樣有誘導軟骨退變的作用,發現Fn-f在軟骨細胞膜上的受體是整合素 α5β1[12,13],并且 Fn-f有增加整合素 α5和 β1亞基表達的作用[14]。整合素α5和β1亞基在間盤內同樣有表達[7],而且整合素α5β1是纖維黏連蛋白和Fn-f在間盤細胞膜上主要的受體[5]。因此,本實驗研究了在Fn-f作用下的髓核細胞整合素α5和β1亞基的表達情況,發現α5和β1亞基表達明顯增加。
研究Fn-f的退變機制時,Homandberg提出Fn-f的作用由Fn-f、MMP、纖維黏連蛋白、II型膠原蛋白組成的正反饋惡性循環,即Fn-f增加了MMP的表達,增加的MMP分解纖維黏連蛋白形成更多的Fnf,而II型膠原蛋白的表達減少是這個惡性循環的最終結局[15],本實驗結果與這個理論一致。本研究同樣發現Fn-f有促進整合素α5、β1亞基表達作用。由于整合素α5β1在Fn-f誘導間盤退變過程中起到了膜受體的作用,而Fn-f又有促進其受體表達增加的作用,那么可以將整合素α5β1納入這個正反饋惡性循環,即Fn-f與髓核細胞膜上的整合素α5β1結合后,影響了髓核細胞的代謝,MMP和受體整合素α5β1的表達都增加,MMP分解纖維黏連蛋白使Fn-f的量增加,而整合素α5β1表達的增加使Fn-f的結合位點增多,促進了Fn-f誘導髓核細胞退變的作用。因此,整合素α5β1既是Fn-f在間盤細胞上的受體,同時又是Fn-f誘導間盤退變的正反饋惡性循環中的關鍵環節。
綜上,本實驗測定了Fn-f對于間盤髓核細胞的影響,初步探討了Fn-f對于間盤整合素α5β1表達的影響。但由于本實驗只是體外培養環境下進行的研究,體內環境下Fn-f誘導間盤退變的機制有待進一步的研究。
[1]Omlor GW,Lorenz H,Engelleiter K,et al.Changes in gene expression and protein distribution at different stages of mechanically induced disc degeneration——an in vivo study on the New Zealand whiterabbit[J].JOrthop Res,2006,24(3):385-392.
[2]Aota Y,An HS,Homandberg G,etal.Differential effectsof fibronectin fragment on proteoglycan metabolism by intervertebral disc cells:a comparison with articular chondrocytes[J].Spine,2005,30(7):722-728.
[3]Greg Anderson D,Li X,Tannoury T,et al.A fibronectin fragment stimulates intervertebral disc degeneration in vivo[J].Spine,2003,28(20):2338-2345.
[4]Anderson DG,Li X,Balian G.A fibronectin fragment alters the metabolismbyrabbitintervertebraldisccellsinvitro[J].Spine,2005,30(11):1242-1246.
[5]Gilchrist CL,Chen J,Richardson WJ,et al.Functional integrin subunitsregulatingcell-matrix interactionsin theintervertebral disc[J].JOrthop Res,2007,25(6):829-840.
[6]Homandberg GA,Costa V,Ummadi V,etal.Antisenseoligonucleotides totheintegrin receptor subunit alpha(5)decreasefibronectin fragmentmediatedcartilagechondrolysis[J].OsteoarthritisCartilage,2002,10(5):381-393.
[7]Nettles DL,Richardson WJ,Setton LA.Integrin expression in cells of theintervertebral disc[J].JAnat,2004,204(6):515-520.
[8]Oegema TRJr,Johnson SL,Aguiar DJ,et al.Fibronectin and its fragments increase with degeneration in the human intervertebral disc[J].Spine,2000,25(21):2742-2747.
[9]Omlor GW,Lorenz H,Engelleiter K,et al.Changes in gene expression and protein distribution at different stages of mechanically induced disc degeneration--an in vivo study on the New Zealand white rabbit[J].JOrthop Res,2006,24(3):385-392.
[10]Nerlich AG,Bachmeier BE,Boos N.Expression of fibronectin and TGF-beta1 mRNA and protein suggest altered regulation of extracellularmatrixindegenerateddisctissue[J].Eur SpineJ,2005,14(1):17-26.
[11]Hsu CC,Lai SC.Matrix metalloproteinase-2,-9 and-13 are involved in fibronectin degradation of rat lung granulomatous fibrosis caused by angiostrongyluscantonensis[J].Int JExp Pathol,2007,88(6):437-443.
[12]Homandberg GA,Costa V,Wen C.Fibronectin fragments active in chondrocytic chondrolysis can be chemically cross-linked to the alpha5 integrin receptor subunit[J].Osteoarthritis Cartilage,2002,10(12):938-949.
[13]Forsyth CB,Pulai J,Loeser RF.Fibronectin fragments and blocking antibodies to alpha2 beta1 and alpha5beta1 integrins stimulate mitogen-activated protein kinase signaling and increase collagenase 3(matrix metalloproteinase 13)production by human articular chondrocytes[J].Arthritis Rheum,2002,46(9):2368-2376.
[14]Takahashi I,Onodera K,Sasano Y,et al.Effect of stretchingon gene expression of beta1 integrin and focal adhesion kinase and on chondrogenesisthroughcell-extracellularmatrixinteractions[J].Eur JCell Biol,2003,82(4):182-192.
[15]Homandberg GA.Potential regulation of cartilagemetabolismin osteoarthritisbyfibronectinfragments[J].Front Biosci,1999,4:D713-730.
