距今4000年前后黃土高原西北邊緣的生業經濟:穩定同位素的證據
2024-07-09 03:04:00侯亮亮楊柳紅王彬馨賈堯張光輝
文物季刊 2024年2期
侯亮亮 楊柳紅 王彬馨 賈堯 張光輝
譯自:Hou LL. Yang LH. Wang BX. Jia Y. Zhang GH. 2022. The subsistence economy on the northwest edge of the Loess Plateau during c. 4000 a BP: Evidence from stable isotopes. Journal of Archaeological Science: Reports, 45, 103616. 譯稿略有刪減。
該研究系國家重點研發計劃項目“中華文明探源研究·北方長城地帶文明進程研究”(編號:2020YFC1521601)階段性成果。
摘要:為揭示距今4000年前后黃土高原西北邊緣傳統粟黍農業與牛羊飼喂等新生產力要素間的關系,本文以碧村遺址3例人骨和53例動物骨骼為研究對象,運用碳氮穩定同位素分析的方法,還原了他們的食譜及家畜飼養方式。結果顯示,部分先民(-7.3‰,8.8‰,n=1)、家豬(-7.9‰±1.7‰,5.9‰±0.7‰,n=15)和家犬(-7.8‰±0.3‰,7.5‰±0.4‰,n=2)主要以粟黍類食物為生;其余人(-13.4‰±0.2‰,6.7‰±1.0‰,n=2)、牛(-13.9‰±1.3‰,5.6‰±0.6‰,n=11)和羊(-16.0‰±1.1‰,5.3‰±0.7‰,n=15)則以C3/C4類混合食物為生。研究顯示,牛和羊既在野生系統中生存,又食用一定量粟黍類食物。當地眾多深溝大壑并不適合粟黍農業發展,但卻適宜放牧牛羊,這無疑能增加物資產出。同時,外來牛和羊沒有侵占傳統家豬的飼喂規模,反而優化了粟黍資源的利用。
關鍵詞:碧村遺址 距今4000年前后 碳氮穩定同位素 粟黍農業 牛羊飼喂
Abstract: To reveal the relationship between millet-based agriculture and new productivity factors (cattle and sheep husbandry) on the northwest edge of the Loess Plateau during c. 4000 a BP, stable carbon and nitrogen isotope analyses of human (n = 3) and faunal (n = 53) bones from the Bicun site were undertaken. Results show that some humans (–7.3‰, 8.8‰, n = 1), pigs (–7.9 ± 1.7‰, 5.9 ± 0.7‰, n = 15), and dogs (–7.8 ± 0.3‰, 7.5 ± 0.4‰, n = 2) mainly lived on millet-based food, while other humans (–13.4 ± 0.2‰, 6.7 ± 1.0‰, n = 2), cattle (–13.9 ± 1.3‰, 5.6 ± 0.6‰, n = 11), and sheep (–16.0 ± 1.1‰, 5.3 ± 0.7‰, n = 15) lived on C3/C4 mixed food. Cattle and sheep not only grazed in wild ecosystems but also ate millet-based foods. Many deep gullies and ravines on the northwest edge of the Loess Plateau are not suitable for the development of millet agriculture but are the ideal pasture for herding cattle and sheep, which undoubtedly promoted material productivity. Meanwhile, instead of encroaching on the development of pig feeding, cattle and sheep husbandry made the utilization of millet-based food more effective.
Keywords: Bicun site c.4000 a BP Carbon and nitrogen stable isotopes Millet agriculture Herding sheep and cattle
一、引 言
史前跨歐亞大陸農作物和家畜的傳播,不僅極大地促進了東西方文化的交流,而且還促使農牧混合經濟模式在中國北方地區的出現和傳播。其中,新的生產力要素,如牛和羊的飼喂畜牧業日益成為改變和推進社會復雜性進程的重要因素。
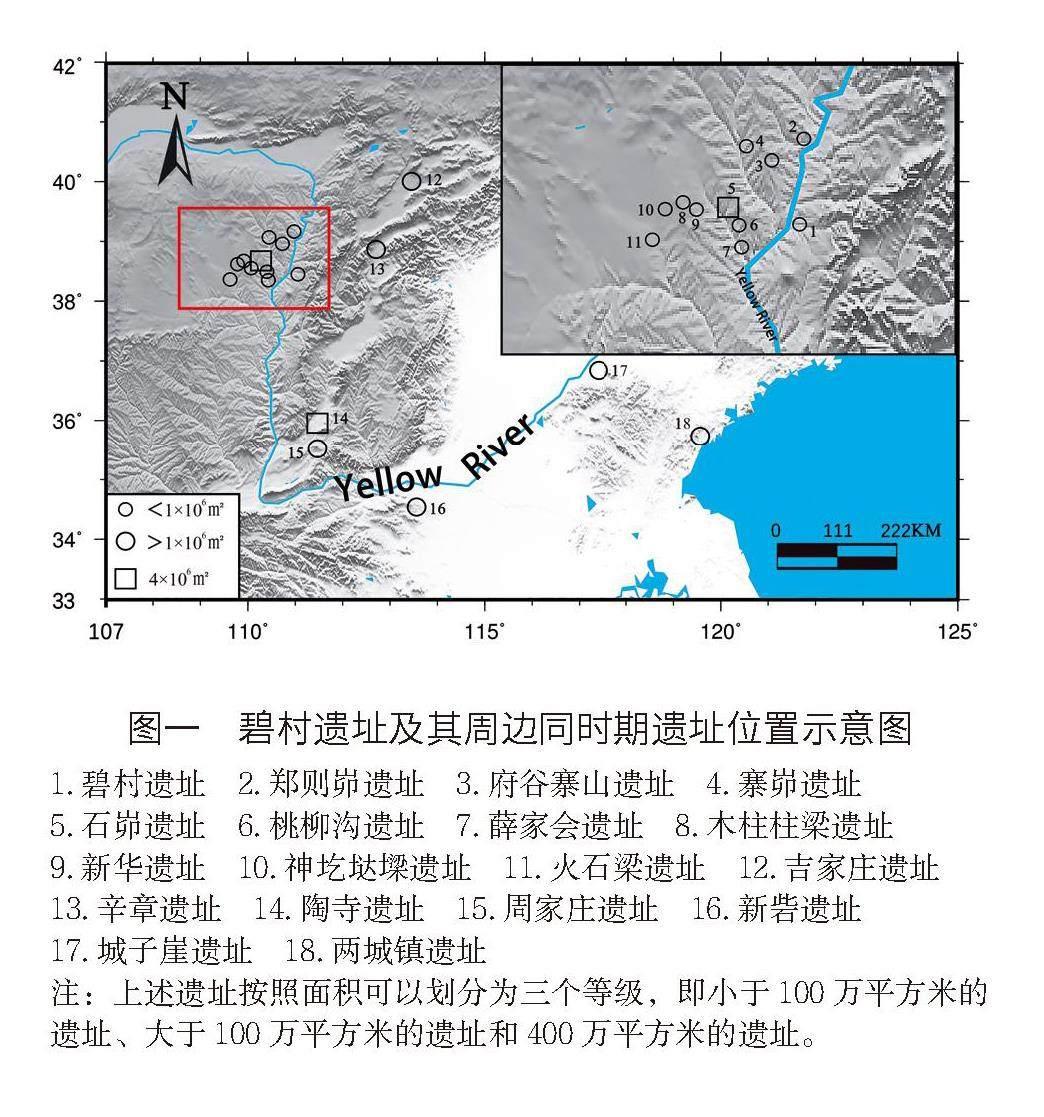
距今4000年前后,中國北方考古學文化的格局急劇變化,不同地區紛紛涌現出各自的文化中心(圖一)。在黃河上游,馬家窯文化之后,齊家文化(4300 BP-3600 BP)迅速發展,很快遍布整個甘青地區的東部。在黃河中游,廟底溝二期文化之后,陜西渭水及其支流兩岸臺地上的客省莊文化(4600 BP-4000 BP),晉南臨汾盆地一帶的陶寺文化(4300 BP-3900 BP),豫中地區的王灣三期文化(4600 BP-3900 BP)、二里頭文化(3750 BP-3500 BP)以及豫北冀南地區的后岡二期文化(4600 BP-4000 BP)紛紛崛起,并各自成為當地的核心文化。在黃河下游,大汶口文化之后,龍山文化(4600 BP-4000 BP)快速發展,并對周邊的考古學文化產生強烈的影響。最近,隨著石峁(約4000 BP)、新砦(4050 BP-3750 BP)和周家莊(4300 BP-3750 BP)等遺址為代表的超大型聚落或城址的發現和研究,為中國早期國家的起源和社會復雜化的研究提供了更為豐富的元素。
多重證據顯示,歐亞大陸農作物(如大麥、小麥)、家畜(如牛、羊)及金屬(如青銅器)的傳播與交換的強化對相關文化的影響和推動尤為明顯。以粟黍農業為基礎兼具家畜(豬)飼喂、同時對牛和羊也采取一定程度的管理措施的生計方式毫無疑問推動了以上地區考古學文化的發展。黃土高原一些地區由于分布著眾多的深溝大壑,可能不太適宜種植粟黍農業,但是牛和羊(特別是羊)的出現使得無法耕種的深溝大壑變成了牧場,變成了創造更多社會物質基礎的生產力。動物考古研究發現,牛和羊在家養動物中所占的比例大體呈現由少到多的趨勢,羊的二次產品(奶、毛等)開始被開發和利用,且傳統粟黍農業也對牛的飼喂產生明顯影響。然而,相對于牛和羊的引入和利用而言,麥類作物和青銅等的開發和利用則相對有限。此外,特別值得一提的是,上述地區的生業經濟模式幾乎一致,但也存有差異,特別是隨著時間的發展使多元化文化元素不斷增加,導致當地生業經濟成分不斷豐富,即多種農作物和家畜互為補充的生業經濟變得尤其重要。
近些年,作為支撐和鏈接石峁等超級大城址發展的衛星城和樞紐點,黃河南流兩岸地區的碧村遺址、寨山遺址、桃柳溝遺址、薛家會遺址等面積在60~100萬平方米的次級聚落開始不斷被發現和研究。同時,同時期面積在20萬平方米及以下的聚落也大量涌現,被不斷地發現和研究。這體現了石峁王國內部的聚落等級分化,面積在60~100萬平方米的聚落當屬石峁都邑下的次級聚落,同時也是不同流域的中心聚落。這些遺址的“零星”發現,反映了距今4000年前后黃土高原西北邊緣不同層級聚落間的階級分化(圖一)。
與石峁遺址隔黃河相望,碧村遺址的發現和研究,經確認屬于石峁遺址之下的次級聚落。碧村遺址地處黃河與蔚汾河的關鍵出入口,遺址規模大,等級高,與神木石峁直線距離也不足百公里,戰略地位和社會輻射效用顯著,這為探索黃土高原西北邊緣地區文化與社會交流,提供了一個重要突破口。同時,碧村遺址石砌房址和城墻的發現,被確認為是規模最大的石砌房址,并在小玉梁上確認了大型石砌排房的存在,進一步確定了這是一個出現過社會復雜化的聚落。
然而,在歐亞大陸農作物、家畜及金屬傳播與交換強化的全球化大浪潮下,黃河南流兩岸地區的考古學文化大發展是否也與此轉變有關,傳統的粟黍農業及外來的牛羊家畜飼養業在本地考古學文化大發展中起到何種作用,本地與周邊相關區域的互動關系等學術問題,依然缺乏相應的探討。
鑒于此,本研究擬以黃河南流東岸山西興縣蔚汾河流域的碧村遺址人和動物骨骼為研究對象,開展C、N穩定同位素分析,重建先民和相關動物的食物結構,探討距今4000年前后黃土高原西北邊緣先民的生業經濟及其與相關考古學文化發展的關系,特別是史前歐亞大陸農業傳播和東西方文化交流對該區域的影響程度等。
二、材料與方法
(一)樣品選取
碧村遺址(38°51′N,110°89′E)位于山西省呂梁市興縣高家村鎮碧村村北,地處黃河與蔚汾河的交匯處。據出土器物及其組合群初步研究表明,碧村遺址的相對年代主要集中在龍山文化晚期至夏紀年前后。同時期臨近的石峁遺址、木柱柱梁遺址的人骨AMS-14C測年顯示它們的年代均在距今4000年前后。
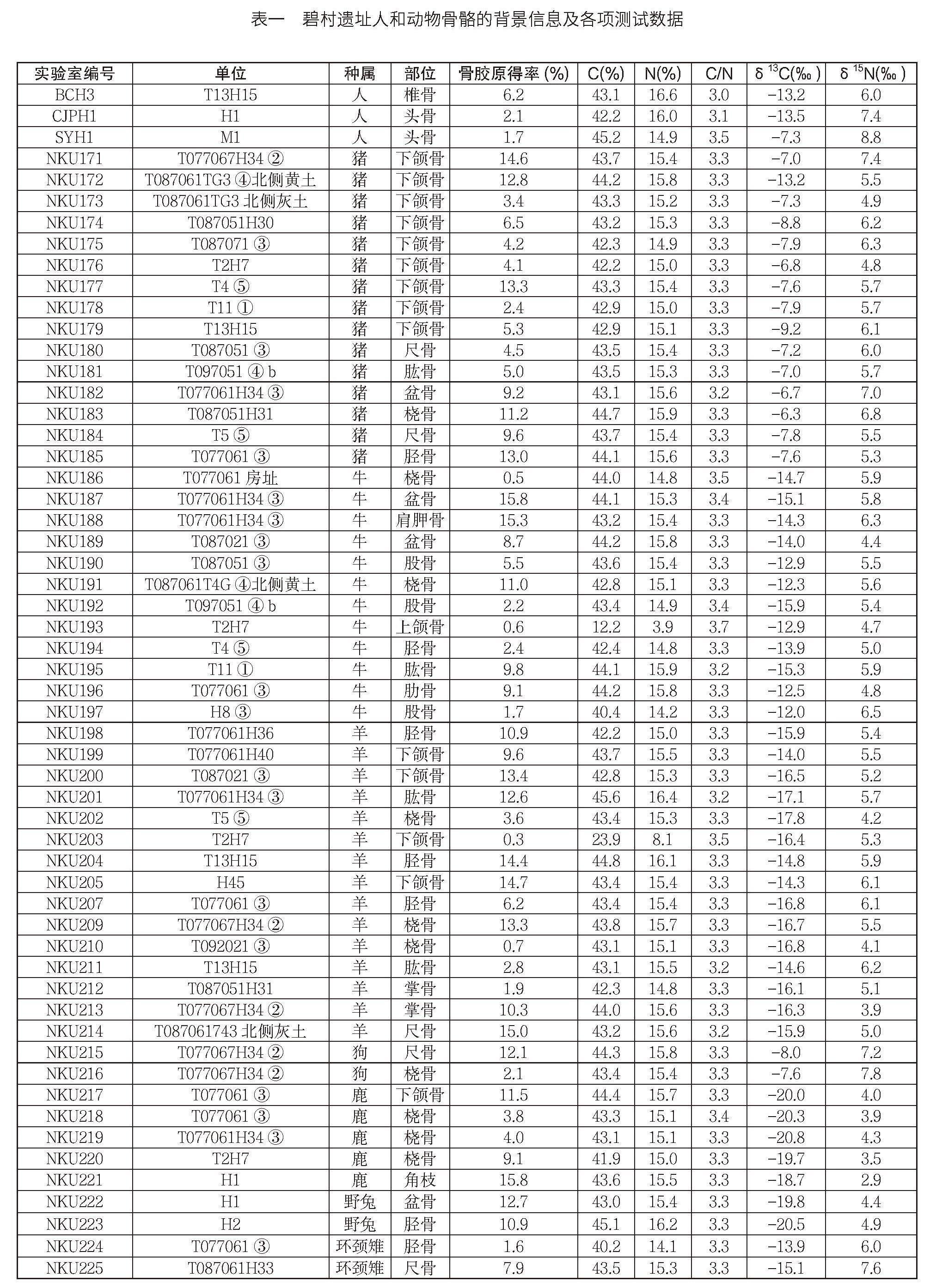
根據人和動物骨骼出土情況,本研究選取碧村遺址家豬15例、狗2例、牛12例、羊15例、鹿5例、野兔2例、環頸雉2例及分屬在1座墓葬和2個灰坑的先民3例,進行C、N穩定同位素分析(表一)。依據遺址不同區域內灰坑出土的骨骼樣本分布特征可知,本次提取的樣本可能超過10只豬、4只羊、2頭牛和1只狗。樣本的詳細信息如表一所示。
(二)穩定同位素分析
依據Richards和Hedges文中的方法,并參考Jay和Richards對該方法的修改。首先,機械去除骨樣表面污染,取1g左右的骨樣放入試管,倒入10ml濃度為0.5mol/L的HCl 溶液4℃浸泡,每隔2~3天更換酸液,直至骨樣松軟,無明顯氣泡,用去離子水清洗至中性。置于濃度為0.001mol/L的HCl溶液,在烘箱70℃條件下明膠化48小時。取出后熱濾,再經Millipore Amicon Ultra-4超濾后收集分子量>30 kD的溶液,放入冰箱冷凍。最后,真空冷凍干燥,收集骨膠原,稱重并計算骨膠提取率(骨膠原重量/骨樣重量,表一)。
骨膠原的C、N元素含量及同位素比值測試由中國科學院大學考古同位素實驗室完成,測試所用儀器是穩定同位素質譜分析儀(Isoprime 100 coupled with Elementar Vario)。測試C、N元素含量所用的標準物質為磺胺(Sulfanilamide)。C、N穩定同位素比值,分別以IAEA-600、IAEA-CH-6標定碳鋼瓶氣(以VPDB為基準)和IAEA-600、IAEA-N-2標定氮鋼瓶氣(以AIR為基準)。此外,每測試10個樣品中插入一個實驗室自制膠原蛋白標樣(δ13C值為-14.7‰±0.1‰,δ15N值為7.0‰±0.1‰)。樣品的同位素比值,以δ13C和δ15N值表示,分析精度均低于±0.2‰。測試數據見表一。
三、結果與討論
(一)骨骼污染鑒別
從表一可知,56例樣品均提取到足量的骨膠原。56例樣品的骨膠原的C、N含量分布范圍分別在12.2%~45.6%和3.9%~16.6%之間,均值分別為42.5%±5.0%和15.0%±1.9%,與現代樣品的C、N含量(分別為41.0%、15.0%)接近。同時,骨膠原提取率的差異也較大,分布范圍在0.3%~15.8%之間,均值為7.7%±4.9%,與現代樣品中骨膠原的含量(20.0%)有很大的差別,說明骨樣在埋藏過程中受到一定的污染。此外,樣品的C/N摩爾比值在3.0~3.7之間,部分樣品超出了未受污染樣品的C/N摩爾比值(2.9~3.6)的范圍內,說明部分樣品受到了污染。
經過分析,可知NKU193的C/N摩爾比值為3.7,說明不可用于后續的穩定同位素分析。因此,除NKU193外,以上55個樣品提取出的骨膠原,均可用作穩定同位素分析。
(二)動物的穩定同位素分析
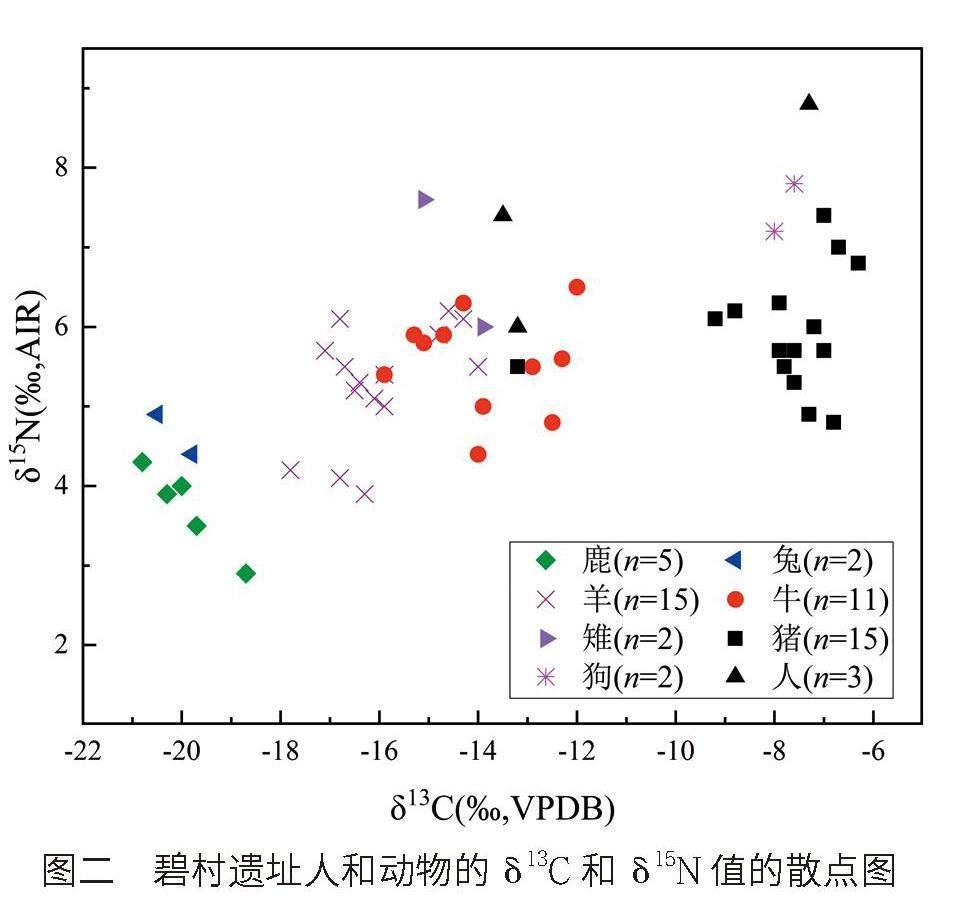
圖二為碧村遺址人和動物骨膠原的δ13C和δ15N值的散點圖。由圖二可見,不同種屬動物的δ13C和δ15N值較為分散,說明它們的食物結構存在較大的差異,這應該與它們的生活方式和飲食習慣相關。
野生草食動物,如草兔、鹿等的穩定同位素比值,一般可以作為當地野生植被穩定同位素基準值的重要參考。如圖二所示,本研究中鹿的δ13C值和δ15N值最低(-19.9‰±0.8‰,3.7‰±0.5‰,n=5),說明此鹿主要以C3類草類和樹葉等為食,也反映了當地的野生植被是以C3類植物為主。草兔的δ13C值(-20.2‰±0.5‰,n=2)與鹿的相應值接近,δ15N值(4.7‰±0.4‰,n=2)略高于鹿的相應值,但在食草動物的δ15N值范圍之內,進一步說明當地的野生植被是以C3類植物為主。因此,上述野生食草動物的穩定同位素特征(-20.0‰±0.7‰,4.0‰±0.6‰,n=7),確定了當地的野生植被是C3類植物為主。此外,蔚汾河流域的考古調查過程中浮選并提取到的大植物遺存結果顯示,和粟黍農作物伴生的雜草既有C3類植物也有C4類植物,即以黍亞科、藜科和豆科植物為主,這也為當地的植被類型提供了一定的參考。
如圖二所示,羊的δ13C值和δ15N值(-16.0‰±1.1‰,5.3‰±0.7‰,n=15)明顯高于鹿和兔的相應值,表明它們的食物也主要為C3類食物,輔以少量C4類食物。然而,這些羊的δ15N值差異較大,說明部分羊的食物中包含了δ15N值較高的植物,這可能和季節性投喂飼料有關,如冬春季節投喂粟黍類的秸稈等。牛的δ13C值(-13.9‰±1.3‰,n=11)總體上略高于羊,但其δ15N值(5.6‰±0.6‰,n=11)卻相對集中,說明它們食物中包含了更多的C4類植物,可能和粟黍類植物的秸稈有關。
如圖二所示,環頸雉的數量較少,其δ13C值和大部分的牛接近,但δ15N值略高于牛的相應值,這說明它們除了食用野生植物的種子外,還食用粟黍類食物,特別是米蟲或其他昆蟲等高蛋白類食物。
如圖二所示,豬和狗的δ13C值高于牛和羊,這應該與其食物結構差異相關。具體表現為,豬(-7.9‰±1.7‰,5.9‰±0.7‰,n=15)和狗(-7.8‰±0.3‰,7.5‰±0.4‰,n=2)的δ13C值和δ15N值比較接近,明顯表現出以C4類食物為主的食物結構特征。同時,由圖二可見,狗的δ15N均值(7.5‰±0.4‰,n=2)高于家豬的δ15N均值(5.9‰±0.7‰,n=15),說明其食物中的動物蛋白比例更多。
(三)先民的穩定同位素分析
鑒于在碧村遺址及其周邊地區并沒有發現大型的公共墓地,因此沒有有效地獲取足量的先民骨骼,進而無法獲取當時先民的充足的穩定同位素數據,本研究僅獲取3例先民的穩定同位素數據。
如圖二所示,據碧村遺址3例先民的δ13C值和δ15N值差異,可以初步分為兩組群體:BCH3和CJPH1(A組)的δ13C值和δ15N值相對較低(-13.4‰±0.2‰,6.7‰±1.0‰,n=2),說明他們主要以C3/C4類混合類食物為生;SYH1(B組)則擁有較高的δ13C值和δ15N值(-7.3‰,8.8‰,n=1),主要以C4類食物為生。
據C和N穩定同位素在食物鏈中的分餾效應(在不同營養級間C富集1‰~1.5‰,N富集3‰~5‰),可以看出豬(-7.9‰±1.7‰,5.9‰±0.7‰,n=15)和狗(-7.8‰±0.3‰,7.5‰±0.4‰,n=2)的δ13C和δ15N均值與先民SYH1的相應值(-7.3‰,8.8‰,n=1)都在一個營養級之內,說明家豬、家犬與先民的食物結構較為一致。同時,在龍山文化時期北方粟黍農業體系下,先民和豬、狗的共生互惠關系已經非常明確,在特殊情況下家豬和家犬可以作為探索先民生業經濟的替代性指標。鑒于本研究中人骨樣品較少,可以嘗試以碧村遺址家豬和家犬的數據來窺探大部分群體的食物結構和生業經濟。上文分析顯示,家豬和家犬主要以C4類食物為生,因此碧村大部分先民可能和SYH1一樣都以C4類食物為生。此外,如表二和圖三所示,與碧村遺址鄰近的同時期的木柱柱梁遺址先民(-10.3‰~-6.7‰,-8.2‰±1.5‰;7.8‰~9.5‰,8.8‰±0.6‰,n=8)、神圪垯墚遺址大部分先民(-10.5‰~-6.7‰,-8.1‰±1.0‰;6.2‰~11.8‰,8.9‰±1.4‰,n=26)與SYH1的穩定同位素數據接近,這和北方同時期的石峁、新華、辛章、陶寺、新砦及兩城鎮遺址先民的食物結構相似(表二;圖三),說明以種植粟黍為基礎、兼具家畜(豬)飼喂是距今4000年前后中國北方大部分先民的主要生計方式。
先民BCH3和CJPH1(-13.4‰±0.2‰,6.7‰±1.0‰,n=2)與牛(-13.9‰±1.3‰,5.6‰±0.6‰,n=11)和羊(-16.0‰±1.1‰,5.3‰±0.7‰,n=15)的δ13C值和δ15N值接近,說明他們基本都以C3/C4混合類食物為生。然而,需要特別指出的是,先民BCH3和CJPH1的δ15N值相對較低,說明這些先民食物結構中的肉食含量極其有限,因此,這兩例先民更可能以植物性食物為生。結合表二和圖三可知,神圪垯墚遺址有兩例先民(SGDL43,δ13C:-13.3‰,δ15N:8.4‰;SGDL50,δ13C:-14.6‰,δ15N:7.0‰)與碧村遺址BCH3和CJPH1先民的數據接近,這可能與先民的遷徙活動、社會地位以及對外來文化因素(牛、羊、小麥、大麥等)的適應程度等有關。
首先,BCH3和CJPH1和神圪垯墚遺址的兩例居民一樣,都可能并非本地居民。至于他們具體的來源地,今后還需要做進一步研究。
當然,這兩例個體存在特殊值也可能與他們的社會等級有關,他們可能有更多機會獲得不同種類的食物資源。墓葬中的葬式及其隨葬品組合可以在一定程度上反映先民的社會地位等信息。如碧村遺址BCH3和CJPH1的人骨發現于灰坑之內,但SYH1則發現于完整的墓葬之內,這似乎表明前者的社會地位較低。近期,在鄰近的同時期的府谷寨山遺址和石峁遺址都發現了等級分明的群體,也間接說明距今4000年前后黃土高原社會存在階層分化的現象。
另外,值得一提的是,水稻和麥類等C3類谷物在距今4000年前后的陜北地區考古遺址中的出土概率和絕對數量極為有限,但豆類等C3類植物卻時有發現,相關比例僅次于粟黍等。碧村及神圪垯墚部分人群消費了一定比例的C3類植物性食物,很可能就是來自于豆類。《戰國策》記載“韓地險惡,山居,五谷所生,非麥而豆;民之所食,大抵豆飯藿羹”,也說明黃土高原先民以豆及葉果腹。因此,碧村遺址所謂的BCH3和CJPH1低等級人群的動物蛋白攝入量極其有限,但可能大量食用“豆飯藿羹”。
(四)動物的飼喂策略
現有考古發現和研究成果表明,距今4000多年前后的碧村遺址與陜北地區的木柱柱梁、神圪垯墚及石峁遺址一樣,都出土了大量炭化的粟黍、豬、犬及牛和羊的骨骼(圖一)。
根據表一及表三可知,碧村遺址2例家犬和15例家豬的C、N穩定同位素數據與神圪垯墚、新砦、陶寺、辛章等遺址的相關值基本接近(圖一),證實距今4000年前后中國北方地區存在堅實的粟黍農業體系。在這樣的農業體系之下,碧村遺址的豬和狗都受到了先民的控制和管理,依賴粟黍及其副產品為食,同時也經常性地食用先民的殘羹冷炙、剩飯,甚至糞便等。
然而,仔細觀察可以發現,神圪垯墚、新砦、陶寺、辛章和碧村遺址的個別家豬/犬還攝入了一定量的C3類食物。根據地理位置分布情況,大致可以分為兩組來進行討論。首先,新砦和陶寺遺址位于中原腹地,距今4000年前后,該地區“五谷豐登”的農業格局已經基本形成。因此,新砦和陶寺遺址的家豬(犬)很可能攝入一定量的C3類麥類、稻類、豆類及其副產品等。當然,這也可能與多樣化的飼喂方式有關,如在野外采食野生生態系統之下的食物等也可能導致家畜的δ13C值偏低。其次,辛章、碧村和神圪垯墚遺址的地貌環境各具特色,如辛章遺址位于相對封閉的忻定盆地內,碧村和神圪垯墚遺址則位于黃土高原的墚峁之間。植物考古的研究也顯示,辛章、碧村和神圪垯墚遺址所在區域依然以單純的粟黍農業為主,但也發現少部分豆類作物,因此,這些地區部分豬和狗可能也會消耗一些C3類食物,如豆類或野生食物資源。
和本土傳統家豬(犬)的飼喂方式不同,外來牛和羊的飼喂方式更應值得關注。上文分析顯示,碧村遺址牛和羊的食物結構明顯不同于豬和狗,這可能與它們的飼喂方式相關。首先,碧村遺址羊的C穩定同位素值略高于鹿的相應值,說明羊在野外放養的同時,也有少量人為干預的情況,即它們的食物中被添加了少量C4類粟黍的副產品等食物,先民在收獲和加工粟黍以后,秸稈和谷糠等很可能成為羊的食物來源。在冬春季節,野生植被資源稀缺的條件下,羊群在野外無法獲得足量的食物,這時粟黍的副產品極有可能成為其食物的重要補充。同時,碧村遺址還有個別羊所消耗的粟黍類副產品比例較高,說明先民可能會對個別羊進行特殊關照,向其投喂更多的粟黍類食物。
特別值得一提的是,碧村遺址牛食物中的粟黍類副產品比例明顯高于羊,這與同時期多個遺址的情況類似,說明先民對外來的牛采取了與羊不同的飼喂方式。牛是大型反芻動物,單純在野外放養并不能滿足其食物需求,因此,牛在野外放養的同時,必須添加相應的飼料,粟黍的秸稈自然成為牛群飼料的不二之選。因此,以粟黍的副產品(如秸稈等)舍飼牛可能在當時已經成了一種流行模式。顯然,碧村遺址牛和羊等外來家畜的飼喂方式也受到粟黍農業的影響,這些全新的生產力要素在很多不適宜農耕的區域(如黃土高原的深溝大壑)被飼喂和放養。可見,在原有家豬飼喂的基礎上,碧村遺址牛和羊的引入使得家畜體系更加多元,這為該地區原有的生業經濟增添了新的活力和生產力要素。
土壤有機碳分析顯示,北方地區絕大多數野生植被都是以C3類植物為主。多重證據(植物考古、動物考古和穩定同位素考古)表明,中國北方地區早在新石器時代就確立了粟黍農業及其基礎上的家畜(豬)飼喂業。龍山文化晚期,牛、羊等全新的生產力要素的傳入對中國北方傳統的以粟黍為主、兼具家畜(豬)飼喂的生業經濟產生了一定的影響。已有的研究成果發現,豬和狗通常以粟黍及粟黍的副產品、先民的殘羹冷炙、剩飯,甚至糞便等為食。自新石器時代以來,豬往往作為中國北方先民重要的肉食來源,家犬則是人類忠實的伙伴和看家護院的工具,二者屬于共生互惠關系,但家犬有時也是先民肉食資源的重要補充。然而,北方先民對于外來牛和羊的利用則是對其次級產品的開發(如羊毛、牛奶等)。
可見,碧村遺址的豬和狗與先民的共生互惠關系已非常明確,它們受到當地先民嚴格的控制和管理,主要依賴粟黍及其副產品為食,經常性地食用先民的殘羹冷炙、剩飯,甚至糞便等;外來的牛和羊則被放牧于碧村遺址周邊不適合進行農耕的深溝大壑里。在冬春季節,先民可能對其投喂粟黍副產品,如秸稈等,作為其食物的補充,尤其是對于食量較大的牛而言,其對于粟黍類副產品的依賴程度較高。
(五)外來牛和羊的飼喂策略
為了從宏觀上明確碧村遺址先民對牛和羊的飼喂策略,特別是與周圍地區及中原地區先民對牛和羊飼喂策略的異同,本文收集了已經發表的神圪垯墚、辛章、陶寺和新砦等遺址(圖一)牛和羊的穩定同位素數據與本研究的相關數據(表四)進行比較分析,以進一步探討距今4000年前后黃土高原西北緣先民生業模式的獨特性。
由表四和圖四可知,上述相關遺址牛的δ13C值存在較大的差異。具體而言,碧村遺址的相應值和其臨近的神圪垯墚的數據相近。整體上碧村遺址牛的δ13C均值都略低于辛章、陶寺和新砦遺址的牛。從單因素方差分析(One-way ANOVA)來看,碧村遺址牛的δ13C值與新砦和陶寺牛的δ13C值存在極顯著差異(P=0<0.01, P=0.003<0.01),與其它兩個遺址的數值則沒有明顯區分。另外,盡管碧村遺址的牛的δ15N值最低,但和其他遺址一樣都在食草動物的δ15N值的范圍之內。然而,從單因素方差分析(One-way ANOVA)來看,碧村遺址牛的δ15N值與神圪垯墚、陶寺和辛章牛的δ15N值存在顯著性差異(0.01 由表四和圖五可知,上述遺址羊的相關數據都比較接近,說明它們都主要是在野外放養為生。然而,需要特別指出的是,它們的δ15N值存在較大的差異,這可能與中原地區的羊的食物中被添加部分飼料或生活于缺水的環境中有關。從單因素方差分析(One-way ANOVA)來看,碧村遺址羊的δ13C值與新砦和辛章遺址羊的δ13C值分別存在極顯著差異(P=0.006<0.01)和顯著性差異(0.01 相關研究顯示,約從4500 a BP開始,黃土高原西北邊緣地區的先民已對外來的牛和羊進行了放牧,使得畜牧經濟在當時已成為占據當地先民生產生活主導地位的生業經濟。上文分析顯示,碧村遺址先民不僅對外來的牛和羊進行放牧,也向其投喂粟黍的副產品等,也暗示畜牧經濟的出現和產生。此外,相關研究顯示,碧村遺址當時的生態環境足以支撐粟黍農業的持續發展,黃土高原西北邊緣眾多的深溝大壑不適合粟黍農業的高度發展,但卻是放牧牛羊的天然牧場,這無疑促進了物質生產效率的提高。同時,外來牛和羊并沒有侵占碧村傳統家豬(犬)的食物資源,反而能夠更充分有效地利用粟黍及其副產品。顯然,碧村外來牛和羊的飼喂不僅能夠使當地的粟黍農業得以更長足穩定地發展,而且還大大增加了先民的物質產出。 因此,以種植粟黍為主,兼具家畜(豬)飼喂,同時飼喂牛和羊的全新的生業模式為距今4000年前后黃土高原西北邊緣地區考古學文化的發展奠定了廣泛的物質基礎,這直接促進了不同層級聚落(如石峁遺址)的大規模發展,甚至促使著文明的最終出現。 (六)黃土高原西北邊緣地區的社會復雜化 大量考古證據表明,距今4000年前后黃土高原西北邊緣分布著眾多聚落,特別是超大型聚落(如石峁)(圖一),這直接導致社會復雜化的出現。其中,先民賴以生存的物質基礎無疑是推動社會復雜化進程的重要因素之一。上文分析顯示,碧村遺址及其周邊地區以種植粟黍為主,兼具家畜(豬)飼喂,同時飼喂牛和羊的全新的生業模式不僅增添了先民的物質產出,還進一步促進了人口增長、文化交流及聚落的擴張和繁榮,推動了當地的社會復雜化進程。 隨著財富的積累,社會復雜化的進程得以加快。鄰近碧村的寨山遺址出現了四種不同類型的墓葬(圖一),表明該地可能存在社會等級秩序:第一種類型的墓葬有木棺、壁龕(放置陶器和/或殉牲)和殉人;第二種有木棺和壁龕,無殉人;第三種有木棺、無壁龕和殉牲或殉人;第四種無隨葬品。其中,前兩種墓葬數量較少,后兩種相對較多,這可能間接反映了財富被掌握在少數人手里,這部分人群的地位等級較高。另外,值得一提的是,碧村居民存在著兩種不同的食物類型,且分別被埋葬于不同的區域,一組來自于墓葬中,另一批來自于灰坑,這也證明當時出現了社會階級分化的現象。 然而,距今4000年前后黃土高原西北邊緣先民與中原腹地先民的生業經濟仍然存在很大不同。相較于黃土高原西北邊緣單一且發達的粟黍類農耕經濟,中原腹地此時已經出現了“五谷豐登”(粟黍、麥類、水稻、豆類及麻類)的局面。距今4000年前后,氣候發生突變,單一的粟黍類農耕經濟無法持續穩定地支撐當地的發展,在距今3600年后或稍晚,氣候和環境進一步惡化,黃沙漫布以及毛烏素沙漠的擴張使得該地區不再適合發展傳統的農耕經濟,因此,黃土高原地區最終并沒有產生像中原地區那樣高度發達的文明。然而,不可否認的是,黃土高原西北邊緣先民對外來牛和羊所采取的管理措施毫無疑問為當地的生業經濟增添了新的活力,很大程度上加快了社會復雜化進程。 四、結 論 通過對碧村遺址人和動物骨骼的穩定同位素分析,可以得出以下結論: (一)碧村遺址部分先民(-7.3‰,8.8‰,n=1)、家豬(-7.9‰±1.7‰,5.9‰±0.7‰,n=15)和家犬(-7.8‰±0.3‰,7.5‰±0.4‰,n=2)主要以粟黍類食物為生;其余人(-13.4‰±0.2‰,6.7‰±1.0‰,n=2)、牛(-13.9‰±1.3‰,5.6‰±0.6‰,n=11)和羊(-16.0‰±1.1‰,5.3‰±0.7‰,n=15)則以C3/C4混合類食物為生,說明牛和羊也在一定程度上受到了粟黍農業的影響。 (二)碧村遺址的牛和羊不僅在野外放牧,而且還食用了一定量的粟黍的副產品(秸稈),當地眾多深溝大壑并不適合粟黍農業發展,但卻適宜放牧牛羊,這種飼喂模式使得當時社會的物質產出大大增加,為當地考古學文化的發展乃至文明的最終出現奠定了物質基礎。 (三)碧村遺址先民從事以種植粟黍和家豬飼喂為基礎的生業經濟,同時,飼喂外來的牛和羊,牛羊的引進沒有侵占傳統家豬的飼喂規模,反而優化了粟黍資源的利用,這為距今4000年前后當地考古學文化的發展乃至文明的最終出現奠定了廣泛的物質基礎。 a. Kuzmina E. E., The Prehistory of the Silk Road. Philadelphia: University of Pennsylvania Press, 2008; b. Jones M., Hunt H., Lightfoot E., et al., “Food globalization in prehistory,” World Archaeology, 2011, 43(4): 665~675. 中國社會科學院考古研究所:《中國考古學——新石器時代卷》,中國社會科學出版社,2010年。 a. 同[2];b. 中國社會科學院考古研究所:《二里頭(1999-2006)》,文物出版社,2014年。 同[2]。 a. 陜西省考古研究院、榆林市文物考古勘探工作隊、神木縣文體局:《陜西神木縣石峁遺址》,《考古》2013年第7期;b. 北京大學考古文博院、鄭州市文物考古研究所:《河南新密市新砦遺址1999年試掘簡報》,《華夏考古》2000年第4期;c. 中國社會科學院考古研究所河南新砦隊、鄭州市文物考古研究院:《河南新密市新砦遺址2002年發掘簡報》,《考古》2009年第2期;d. 中國國家博物館田野考古研究中心、山西省考古研究所、運城市文物保護研究所:《山西絳縣周家莊遺址居址與墓地2007~2012年的發掘》,《考古》2015年第5期;e. 中國國家博物館田野考古研究中心、山西省考古研究所、運城市文物保護研究所:《山西絳縣周家莊遺址2013年發掘簡報》,《考古》2018年第1期。 a. 趙志軍、何駑:《陶寺城址2002年度浮選結果及分析》,《考古》2006年第5期;b. Flad R., S. Li S. C., Wu X. H., et al., “Early wheat in China: Results from new studies at Donghuishan in the Hexi Corridor,” The Holocene, 2010, 20(6): 955~965. a. 傅羅文、袁靖、李水城:《論中國甘青地區新石器時代家養動物的來源及特征》,《考古》2009年第5期;b. 袁靖:《中國古代家養動物的動物考古學研究》,《第四紀研究》2010年第2期。 李水城:《西北與中原早期冶銅業的區域特征及交互作用》,《考古學報》2005年第3期。 a. 馬敏敏:《公元前兩千紀河湟及其毗鄰地區的食譜變化與農業發展——穩定同位素證據》,蘭州大學博士學位論文,2013年;b. Liu X. Y., Lightfoot E., OConnell T. C., et al., “From necessity to choice: dietary revolutions in west China in the second millennium BC,” World Archaeology, 2014, 46(5): 661~680; c. Ma M. M., Dong G. H., Lightfoot E., et al., “Stable Isotope Analysis of Human and Faunal Remains in the Western Loess Plateau, Approximately 2000 calbc,” Archaeometry, 2014, 56: 237~255; d. Ma M. M., Dong G. H., Jia X., et al., “Dietary shift after 3600 calyr BP and its influencing factors in northwestern China: Evidence from stable isotopes,” Quaternary Science Reviews, 2016, 145: 57~70; e. Wang T. T., Wei D., Chang X. E., et al., “Tianshanbeilu and the Isotopic Millet Road: reviewing the late Neolithic/Bronze Age radiation of human millet consumption from north China to Europe,” National Science Review, 2017, 6(5): 1024~1039. 同[7]b。 戴玲玲、李志鵬、胡耀武等:《新砦遺址出土羊的死亡年齡及畜產品開發策略》,《考古》2014年第1期。 a. 陳相龍:《龍山時代家畜飼養策略研究》,中國科學院大學博士學位論文,2012年;b. Dai L. L., Balasse M., Yuan J., et al., “Cattle and sheep raising and millet growing in the Longshan age in central China: Stable isotope investigation at the Xinzhai site,” Quaternary International, 2016, 426: 145~157; c. Dai L. L., Li Z. P., Zhao C. Q., et al., “An Isotopic Perspective on Animal Husbandry at the Xinzhai Site During the Initial Stage of the Legendary Xia Dynasty (2070-1600 BC),” International Journal of Osteoarchaeology, 2016, 26(5): 885~896. a. Linduff K. M. and Mei J. J., “Metallurgy in Ancient Eastern Asia: Retrospect and Prospects,” Journal of World Prehistory, 2009, 22(3): 265~281; b. Liu X. Y., Lister D. L., Zhao Z. J., et al., “Journey to the east: Diverse routes and variable flowering times for wheat and barley en route to prehistoric China,” PLoS One, 2017, 12(11): 1932~6203. a. 趙志軍:《中華文明形成時期的農業經濟發展特點》,《中國國家博物館館刊》2011年第1期;b. 袁靖:《中華文明探源工程十年回顧:中華文明起源與早期發展過程中的技術與生業研究》,《南方文物》2012年第4期。 王煒林、郭小寧:《陜北地區龍山至夏時期的聚落與社會初論》,《考古與文物》2016年第4期。 同[15]。 同[15]。 a. 山西省考古研究所、興縣文物旅游局:《2015年山西興縣碧村遺址發掘簡報》,《考古與文物》2016年第4期;b. 山西省考古研究所、山西大學歷史文化學院考古系、興縣文物旅游局:《2016年山西興縣碧村遺址發掘簡報》,《中原文物》2017年第6期。 同[18]。 同[18]a。 Atahan P., Dodson J., Li X. Q., et al., “Temporal trends in millet consumption in northern China,” Journal of Archaeological Science, 2014, 50: 171~177. 陳相龍、郭小寧、胡耀武等:《陜西神木木柱柱梁遺址先民的食譜分析》,《考古與文物》2015年第5期。 Richards M. P. and Hedges R. E. M., “Stable Isotope Evidence for Similarities in the Types of Marine Foods Used by Late Mesolithic Humans at Sites Along the Atlantic Coast of Europe,” Journal of Archaeological Science, 1999, 26(6): 717~722. Jay M. and Richards M. P., “Diet in the Iron Age cemetery population at Wetwang Slack, East Yorkshire, UK: carbon and nitrogen stable isotope evidence,” Journal of Archaeological Science, 2006, 33(5): 653~662. Ambrose S. H., “Preparation and characterization of bone and tooth collagen for isotopic analysis,” Journal of Archaeological Science, 1990, 17(4): 431~451. Ambrose S. H., Butler B. M., Hanson D. B., et al., “Stable isotopic analysis of human diet in the Marianas Archipelago, western Pacific,” American Journal Physical Anthropology, 1997, 104(3): 343~361. DeNiro M., “Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction,” Nature, 1985, 317(6040): 806~809. a. Stevens R. E., Lister A. M., Hedges R. E. M., “Predicting diet, trophic level and palaeoecology from bone stable isotope analysis: a comparative study of five red deer populations,” Oecologia, 2006, 149(1): 12~21; b. 管理、胡耀武、胡松梅等:《陜北靖邊五莊果墚動物骨的 C和N穩定同位素分析》,《第四紀研究》2008年第6期。 蔣宇超、戴向明、王力之等:《大植物遺存反映的龍山時代山西高原的農業活動與區域差異》,《第四紀研究》2019年第1期。 Makarewicz C. A., “Winter is coming: seasonality of ancient pastoral nomadic practices revealed in the carbon (δ13C) and nitrogen (δ15N) isotopic record of Xiongnucaprines,” Archaeological and Anthropological Sciences, 2015, 9(3): 405~418. a. Ambrose S. H. and Norr L., “Isotopic composition of dietary protein and energy versus bone collagen and apatite: Purified diet growth experiments”, “Molecular Archaeology of Prehistoric Human Bone (Prehistoric Human Bone-Archaeology at the Molecular Level)”, Springer, 1993: 1~37; b. Hedges R. E. M. and Reynard L. M., “Nitrogen isotopes and the trophic level of humans in archaeology,” Journal of Archaeological Science, 2007, 34(8): 1240~1251; c. 同[28]。 侯亮亮:《穩定同位素視角下重建先民生業經濟的替代性指標》,《南方文物》2019年第2期。 同[21]。 同[21]。 侯亮亮、趙杰、鄧惠等:《穩定同位素和植物微體化石證據所見山西忻定盆地4000a BP前后的生業經濟》,《中國科學:地球科學》2019年第3期。 張雪蓮、仇士華、薄官成等:《二里頭遺址、陶寺遺址部分人骨碳十三、氮十五分析》,《科技考古(第2輯)》,科學出版社,2007年。 a. 吳小紅、肖懷德、魏彩云等:《河南新砦遺址人、豬食物結構與農業形態和家豬馴養的穩定同位素證據》,《科技考古(第2輯)》,科學出版社,2007年;b. 同[12]b。 Lanehart R. E., Tykot R. H., 加里·費曼等:《山東日照市兩城鎮遺址龍山文化先民食譜的穩定同位素分析》,《考古》2008年第8期。 陳相龍、郭小寧、王煒林等:《陜北神圪垯墚遺址4000a BP前后生業經濟的穩定同位素記錄》,《中國科學:地球科學》2017年第1期。 陜西省考古研究院、榆林市文物保護研究所、府谷縣文管辦:《陜西府谷寨山遺址廟墕地點居址發掘簡報》,《文博》2021年第5期。 同[9]d。 同[39]。 同[18]b。 同[18]b。 同[5]a、[40]。 郭小寧:《陜北地區龍山晚期的生業方式——以木柱柱梁、神圪垯梁遺址的植物、動物遺存為例》,《農業考古》2017年第3期。 劉向:《戰國策》,上海古籍出版社,2020年。 同[47]。 同[39]。 同[22]。 同[36]。 同[37]a。 同[35]。 同[46]。 同[46]。 楊瑞琛、邸楠、賈鑫等:《從石峁遺址出土植物遺存看夏時代早期榆林地區先民的生存策略選擇》,《第四紀研究》2022年第1期。 胡松梅、楊苗苗、孫周勇等:《2012~2013年度陜西神木石峁遺址出土動物遺存研究》,《考古與文物》2016年第4期。 同[39]。 同[12]b、[12]c。 陳相龍、袁靖、胡耀武等:《陶寺遺址家畜飼養策略初探:來自碳、氮穩定同位素的證據》,《考古》2012年第9期。 同[35]。 何駑:《制度文明:陶寺文化對中國文明的貢獻》,《南方文物》2020年第3期。 戴向明:《中原地區龍山時代社會復雜化的進程》,《考古學研究(十)》,科學出版社,2013年。 Sealy J. C., van der Merwe N. J., Lee Thorp J. A., et al., “Nitrogen isotopic ecology in southern Africa: Implications for environmental and dietary tracing,” Geochimica et Cosmochimica Acta, 1987, 51(10): 2707~2717. 同[35]。 a. 王輝、王曉毅、張光輝等:《山西興縣碧村遺址聚落選址的地貌背景》,《南方文物》2021年第5期;b. 同[39]。 同[29]、[46]。 a. 四川農業大學:《畜牧學》,中國農業出版社,1989年;b. 同[30]。 a. 山西農業大學:《養羊學》,農業出版社,1992年;b. 夏明、閻志堅:《放牧綿羊選食次序和季節性食譜重疊》,《中國草地》2000年第2期;c. 汪詩平:《不同放牧季節綿羊的食性及食物多樣性與草地植物多樣性間的關系》,《生態學報》2000年第6期。 a. 同[12]b、[12]c、[39];b. 侯亮亮、李素婷、胡耀武等:《先商文化時期家畜飼養方式初探》,《華夏考古》2013年第2期。 昝林森:《牛生產學》,中國農業出版社,2007年。 同[60]。 趙輝:《“古國時代”》,《華夏考古》2020年第6期。 楊英、沈承德、易惟熙等:《21ka以來渭南黃土剖面的元素碳記錄》,《科學通報》2001年第8期。 a. 同[7]b;b. 張之恒:《黃河流域的史前粟作農業》,《中原文物》1998年第3期;c. 董廣輝、張山佳、楊誼時等:《中國北方新石器時代農業強化及對環境的影響》,《科學通報》2016年第26期。 a. 同[7]b;b. 趙志軍:《小麥傳入中國的研究——植物考古資料》,《南方文物》2015年第3期;c. 董廣輝、楊誼時、韓建業等:《農作物傳播視角下的歐亞大陸史前東西方文化交流》,《中國科學:地球科學》,2017年第5期。 a. Barton L., Newsome S. D., Chen F. H., et al., “Agricultural origins and the isotopic identity of domestication in North China,” Proceedings of the National Academy of Sciences of the United States of America, 2009, 106, 5523~5528; b. Atahan P., Dodson J., Li X. Q., et al., “Early Neolithic diets at Baijia, Wei River valley, China: Stable carbon and nitrogen isotope analysis of human and faunal remains,” Journal of Archaeological Science, 2011, 38, 2811~2817; c. Hongo H., Kikuchi H., Nasu H., “Beginning of pig management in Neolithic China: comparison of domestication processes between northern and southern regions,” Animal Frontiers, 2011, 11, 30~42. 武莊:《先秦時期家犬研究》,中國社會科學院研究生院博士學位論文,2014年。 同[11]、[12]b、[12]c。 同[39]。 同[12]b、[12]c。 同[60]。 同[35]。 a. 同[63];b. 高江濤:《新砦遺址與中國古代文明起源問題》,《中原文物》2005年第4期。 a. 同[60];b. 姚政權、吳妍、王昌燧等:《河南新密市新砦遺址的植硅石分析》,《考古》2007年第3期;c. 鐘華、趙春青、魏繼印等:《河南新密新砦遺址2014年浮選結果及分析》,《農業考古》2016年第1期。 同[60]。 同[30]。 同[18]b。 同[63]。 同[64]、[66]a。 同[30]、[64]。 同[30]。 同[30]、[64]。 胡松梅、楊曈、楊苗苗等:《陜北靖邊廟梁遺址動物遺存研究兼論中國牧業的形成》,《第四紀研究》2022年第1期。 同[66]a。 同[73]。 a. 裴學松:《石峁文化墓葬初探》,《考古與文物》2022年第2期;b.同[40];c. 陜西省考古研究院、榆林市文物保護研究所、府谷縣文管辦:《陜西府谷寨山遺址廟地點墓地發掘簡報》,《考古與文物》2022年第2期。 同[40]、[97]c。 王輝、莫多聞、潘永剛等:《陜西神木新華遺址環境考古研究》,《神木新華》,科學出版社,2005年。
